
Long Noncoding RNAs and Cancer Stem Cells: Dangerous Liaisons Managing Cancer

Abstract
:1. Introduction
2. Long Non-Coding RNAs
2.1. Biogenesis and Conservation of lncRNAs
2.2. LncRNAs Classification
- -
- sense lncRNAs and antisense lncRNAs overlap with one or more exons of neighboring mRNAs, respectively on the same and the opposite strand;
- -
- intronic lncRNAs are transcribed from introns of the protein coding genes;
- -
- promoter upstream lncRNAs or promoter-associated lncRNAs whose genes are located upstream or close to the promoter of a different gene;
- -
- intergenic lncRNAs, whose genes lie within the genomic interval between two genes;
- -
- bidirectional lncRNAs, which share the promoter with protein-encoding genes but are transcribed in the opposite direction;
- -
- 3’ UTR-associated RNAs, derived from the 3’-untranslated region of the protein-coding transcript [22].
2.3. LncRNAs and Organelles
3. Nuclear lncRNAs and CSCs
4. Cytoplasmic lncRNAs and CSCs
4.1. Long Noncoding RNAs Working as Competing Endogenous RNAs
4.2. Long Noncoding RNAs Working through the Interaction with RNA-Binding Proteins
5. Exosomal lncRNAs
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Battle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, D.; Dick, J.E. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat. Med. 1997, 3, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Visvader, J.E.; Lindeman, G.J. Cancer stem cells in solid tumours: Accumulating evidences and unresolving questions. Nat. Rev. Cancer 2008, 8, 755–768. [Google Scholar] [CrossRef]
- Donnenberg, V.S.; Donnenberg, A.D. Stem cell state and the epithelial-to-mesenchymal transition: Implications for cancer therapy. J. Clin. Pharmacol. 2015, 55, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Orkin, S.H. Embryonic stem cell specific signatures in cancer: Insights into genomic regulatory networks and implications for medicine. Genome Med. 2011, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Papaccio, F.; Paino, T.; Regad, G.; Papaccio, V.; Tirino, V. Concise Review: Cancer Cells, Cancer Stem Cells, and Mesenchymal Stem Cells: Influence in Cancer Development. Stem Cells Transl. Med. 2017, 6, 2115–2125. [Google Scholar] [CrossRef]
- Takebe, N.; Miele, L.; Hariss, P.J.; Jeong, W.; Bando, H.; Kahn, M.; Yang, S.Y.; Ivy, S.P. Targeting Notch, Hedgehog, and Wnt pathways in cancer stem cells: Clinical update. Nat. Rev. Clin. Oncol. 2015, 12, 445–464. [Google Scholar] [CrossRef]
- Gonzalez, M.; Moore, H.M.; Li, X.; Toy, K.A.; Huang, W.; Sabel, M.S.; Kidwell, K.M.; Kleer, C.G. EZH2 expands breast stem cells through activation of NOTCH1 signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 3098–3103. [Google Scholar] [CrossRef] [Green Version]
- Valencia, A.; Roman-Gomez, J.; Cervera, J.; Such, E.; Barragán, E.; Bolufer, P.; Moscardó, F.; Sanz, G.F.; Sanz, M.A. Wnt signaling pathway is epigenetically regulated by methylation of Wnt antagonists in acute myeloid leukemia. Leukemia 2009, 23, 1658–1666. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Kwon, H.Y.; Zimdahl, B.; Congdon, K.L.; Blum, J.; Lento, W.E.; Zhao, C.; Lagoo, A.; Gerrard, G.; Foroni, L.; et al. Regulation of myeloid leukaemia by the cell-fate determinant Musashi. Nature 2010, 466, 765–768. [Google Scholar] [CrossRef]
- Marusyk, A.; Polyak, K. Tumor heterogeneity: Causes and consequences. Biochim. Biophys. Acta 2010, 1805, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yu, Q. Intra-tumor heterogeneity of cancer cells and its implications for cancer treatment. Acta Pharmacol. Sin. 2015, 36, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Berabez, N.; Durand, S.; Gabut, M. Post-transcriptional regulations of cancer stem cell homeostasis. Curr. Opin. Oncol. 2019, 31, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Sonawala, K.; Ramalingam, S.; Sellamuthu, I. Influence of Long Non-Coding RNA in the Regulation of Cancer Stem Cell Signaling Pathways. Cells 2022, 11, 3492. [Google Scholar] [CrossRef] [PubMed]
- McCabe, E.M.; Rasmussen, T.P. lncRNA involvement in cancer stem cell function and epithelial-mesenchymal transitions. Semin. Cancer Biol. 2021, 75, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.; Koonin, E.V. Functional long non-coding RNAs evolve from junk transcripts. Cell 2020, 183, 1151–1161. [Google Scholar] [CrossRef]
- Oo, J.A.; Brandes, R.P.; Leisegang, M.S. Long non-coding RNAs: Novel regulators of cellular physiology and function. Pflüg. Arch. 2021, 474, 191–204. [Google Scholar] [CrossRef]
- Flynn, R.A.; Chang, H.Y. Long noncoding RNAs in cell-fate programming and reprogramming. Cell Stem Cell 2014, 14, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Wan, P.; Su, W.; Zhuo, Y. The role of long noncoding RNAs in neurodegenerative diseases. Mol. Neurobiol. 2017, 54, 2012–2021. [Google Scholar] [CrossRef]
- DiStefano, J.K.; Gerhard, G.S. Long noncoding RNAs and human liver disease. Annu. Rev. Pathol. 2022, 17, 1–21. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long non-coding RNA (lncRNA) roles in cell biology, neurodevelopment and neurological disorders. Non-Coding RNA 2021, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, B.; Ulitsky, I. Predictive models of subcellular localization of long RNAs. Rna 2019, 25, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Jégu, T.; Blum, R.; Cochrane, J.C.; Yang, L.; Wang, C.-Y.; Gilles, M.-E.; Colognori, D.; Szanto, A.; Marr, S.K.; Kingston, R.E.; et al. Xist RNA antagonizes the SWI/SNF chromatin remodeler BRG1 on the inactive X chromosome. Nat. Struct. Mol. Biol. 2019, 26, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; He, X.; Lu, F.; Li, L.; Song, H.; Bian, X. Research progress regarding long-chain non-coding RNA in lung cancer: A narrative review. J. Thorac. Dis. 2022, 14, 3016–3029. [Google Scholar] [CrossRef]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Potacio, A.; Flynn, R.A.; Gupta, R.A.; et al. Long noncoding RNA programs active chromatin domain to coordinate homeotic gene activation. Nature 2011, 472, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Wu, H.; He, Y.; Chen, H.; Liu, Y.; Wei, B.; Chen, G.; Lin, H.; Lin, H. LncRNA THOR increases osteosarcoma cell stemness and migration by enhancing SOX 9 mRNA stability. FEBS Open Bio 2019, 9, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.X.; Koirala, P.; Zhang, W.; Ni, C.; Wang, Z.; Yang, L.; Mo, Y.Y. lncRNA RMST enhances DNMT3 expression through interaction with HuR. Mol. Ther. 2020, 28, 9–18. [Google Scholar] [CrossRef]
- López-Urrutia, E.; Bustamante Montes, L.P.; de Guevara Cervantes, D.L.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk between long non-coding RNAs, micro-RNAs and mRNAs: Deciphering molecular mechanisms of master regulators in cancer. Front. Oncol. 2019, 9, 669. [Google Scholar] [CrossRef]
- Kartha, R.V.; Subramanian, S. Competing endogenous RNAs (ceRNAs): New entrants to the intricacies of gene regulation. Front. Genet. 2014, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Pasieka, R.; Zasoński, G.; Raczyńska, K.D. Role of Long Intergenic Noncoding RNAs in Cancers with an Overview of MicroRNA Binding. Mol. Diagn. Ther. 2023, 27, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.H.; Kim, K.M.; Abdelmohsen, K.; Yoon, J.H.; Panda, A.C.; Munk, R.; Kim, J.; Curtis, J.; Moad, C.A.; Wohler, C.M.; et al. HuR and GRSF1 modulate the nuclear export and mitochondrial localization of the lncRNA RMRP. Genes Dev. 2016, 30, 1224–1239. [Google Scholar] [CrossRef] [Green Version]
- Statello, L.; Maugeri, M.; Garre, E.; Nawaz, M.; Wahlgren, J.; Papadimitriou, A.; Lundqvist, C.; Lindfors, L.; Collén, A.; Sunnerhagen, P.; et al. Identification of RNA-binding proteins in exosomes capable of interacting with different types of RNA: RBP-facilitated transport of RNAs into exosomes. PLoS ONE 2018, 13, e0195969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, P.; Luo, J.; Wang, J.; Liu, Z.; Wu, W.; Du, Y.; Ye, B.; Wang, D.; He, L.; et al. LncRNA HAND2-AS1 promotes liver cancer stem cell self-renewal via BMP signaling. EMBO J. 2019, 38, e101110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Wang, D.; Li, J.; Wang, Q.; Wo, L.; Zhang, X.; Hu, Z.; Wang, Z.; Zhan, M.; He, M.; et al. TGFB2-AS1 inhibits triple-negative breast cancer progression via interaction with SMARCA4 and regulating its targets TGFB2 and SOX2. Proc. Natl. Acad. Sci. USA 2022, 119, e2117988119. [Google Scholar] [CrossRef]
- Zagorac, S.; de Giorgio, A.; Dabrowska, A.; Kalisz, M.; Casas-Vila, N.; Cathcart, P.; Yiu, A.; Ottaviani, S.; Degani, N.; Lombardo, Y.; et al. SCIRT lncRNA Restrains Tumorigenesis by Opposing Transcriptional Programs of Tumor-Initiating Cells. Cancer Res. 2021, 81, 580–593. [Google Scholar] [CrossRef]
- Chen, X.; Xie, R.; Gu, P.; Huang, M.; Han, J.; Dong, W.; Xie, W.; Wang, B.; He, W.; Zhong, G.; et al. Long noncoding RNA LBCS inhibits self-renewal and chemoresistance of bladder cancer stem cells through epigenetic silencing of SOX2. Clin. Cancer Res. 2019, 25, 1389–1403. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhao, B.S.; Zhou, A.; Lin, K.; Zheng, S.; Lu, Z.; Chen, Y.; Sulman, E.P.; Xie, Z.; Bögler, O.; et al. m6A demethylase ALKBH5 maintains tumorigenicity of glioblastoma stem-like cells by sustaining FOXM1 expression and cell proliferation program. Cancer Cell 2017, 31, 591–606. [Google Scholar] [CrossRef]
- Ji, J.; Xu, R.; Ding, K.; Bao, G.; Zhang, X.; Huang, B.; Wang, X.; Martinez, A.; Wang, X.; Li, G.; et al. Long Noncoding RNA SChLAP1 Forms a Growth-Promoting Complex with HNRNPL in Human Glioblastoma through Stabilization of ACTN4 and Activation of NF-κB SignalingSChLAP1 Forms a Complex with HNRNPL in Glioblastoma. Clin. Cancer Res. 2019, 25, 6868–6881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, K.L.; Vasudevan, H.N.; Lockney, D.T.; Baum, R.; Hendrickson, R.C.; Raleigh, D.R.; Schmitt, A.M. Miat and interacting protein Metadherin maintain a stem-like niche to promote medulloblastoma tumorigenesis and treatment resistance. Proc. Natl. Acad. Sci. USA 2022, 119, e2203738119. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Jin, X.; Han, L.; Huang, H.; Ji, Z.; Xu, X.; Tang, M.; Jiang, B.; Chen, W. Exosomal lncRNA DOCK9-AS2 derived from cancer stem cell-like cells activated Wnt/β-catenin pathway to aggravate stemness, proliferation, migration, and invasion in papillary thyroid carcinoma. Cell Death Dis. 2020, 11, 743. [Google Scholar] [CrossRef]
- Ayala, R.; Willhoft, O.; Aramayo, R.J.; Wilkinson, M.; McCormack, E.A.; Ocloo, L.; Wigley, D.B.; Zhang, X. Structure and regulation of the human INO80–nucleosome complex. Nature 2018, 556, 391–395. [Google Scholar] [CrossRef]
- Eustermann, S.; Schall, K.; Kostrewa, D.; Lakomek, K.; Strauss, M.; Moldt, M.; Hopfner, K.P. Structural basis for ATP-dependent chromatin remodelling by the INO80 complex. Nature 2018, 556, 386–390. [Google Scholar] [CrossRef]
- Krietenstein, N.; Wal, M.; Watanabe, S.; Park, B.; Peterson, C.L.; Pugh, B.F.; Korber, P. Genomic nucleosome organization reconstituted with pure proteins. Cell 2016, 167, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Du, Y.; Ward, J.M.; Shimbo, T.; Lackford, B.; Zheng, X.; Miao, Y.L.; Zhou, B.; Han, L.; Fargo, D.C.; et al. INO80 facilitates pluripotency gene activation in embryonic stem cell self-renewal, reprogramming, and blastocyst development. Cell Stem Cell 2014, 14, 575–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derynck, R.; Turley, S.J.; Akhurst, R.J. TGFβ biology in cancer progression and immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 9–34. [Google Scholar] [CrossRef]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 2010, 142, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, J.; Zhang, H.; Ge, L.; Li, J.; Wang, H. RNA m6A methylation in cancer. Mol. Oncol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sher, G.; Masoodi, T.; Patil, K.; Akhtar, S.; Kuttikrishnan, S.; Ahmad, A.; Uddin, S. Dysregulated FOXM1 signaling in the regulation of cancer stem cells. Semin. Cancer Biol. 2022, 86, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Aksenova, V.; Turoverova, L.; Khotin, M.; Magnusson, K.E.; Tulchinsky, E.; Melino, G.; Pinaev, G.P.; Barlev, N.; Tentler, D. Actin-binding protein alpha-actinin 4 (ACTN4) is a transcriptional co-activator of RelA/p65 sub-unit of NF-kB. Oncotarget 2013, 4, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, N.; Ozaki, K.; Sato, H.; Mizuno, H.; Saito, S.; Takahashi, A.; Miyamoto, Y.; Ikegawa, S.; Kamatani, N.; Hori, M.; et al. Identification of a novel non-coding RNA, MIAT, that confers risk of myocardial infarction. J. Hum. Genet. 2006, 51, 1087–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.; Sarkar, D. The Scope of Astrocyte Elevated Gene-1/Metadherin (AEG-1/MTDH) in Cancer Clinicopathology: A Review. Genes 2021, 12, 308. [Google Scholar] [CrossRef]
- Peng, F.; Wang, J.H.; Fan, W.J.; Meng, Y.T.; Li, M.M.; Li, T.T.; Cui, B.; Wang, H.F.; Zhao, Y.; An, F.; et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene 2018, 37, 1062–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Wang, L.; Chen, T.; Shi, Y.; Yao, B.; Liu, Z.; Wang, Y.; Li, Q.; Liu, R.; Niu, Y.; et al. LncRNA RUNX1-IT1 which is downregulated by hypoxia-driven histone deacetylase 3 represses proliferation and cancer stem-like properties in hepatocellular carcinoma cells. Cell Death Dis. 2020, 11, 95. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Jiang, X.; Duan, L.; Xiong, Q.; Yuan, Y.; Liu, P.; Jiang, L.; Shen, Q.; Zhao, S.; Yang, C.; et al. LncRNA PKMYT1AR promotes cancer stem cell maintenance in non-small cell lung cancer via activating Wnt signaling pathway. Mol. Cancer 2021, 20, 156. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, Z.; He, S.; Gong, Y.; He, A.; Li, Y.; Zhang, L.; Zhang, X.; Fang, D.; Li, X.; et al. Long non-coding RNA SOX2OT promotes the stemness phenotype of bladder cancer cells by modulating SOX2. Mol. Cancer 2020, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wan, H.; Zhang, X. LncRNA LHFPL3-AS1 contributes to tumorigenesis of melanoma stem cells via the miR-181a-5p/BCL2 pathway. Cell Death Dis. 2020, 11, 950. [Google Scholar] [CrossRef]
- He, W.; Liang, B.; Wang, C.; Li, S.; Zhao, Y.; Huang, Q.; Liu, Z.; Yao, Z.; Wu, Q.; Liao, W.; et al. MSC-regulated lncRNA MACC1-AS1 promotes stemness and chemoresistance through fatty acid oxidation in gastric cancer. Oncogene 2019, 38, 4637–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Liu, B.; Chen, Z.; Li, G.; Zhang, Z. MSC-induced lncRNA HCP5 drove fatty acid oxidation through miR-3619-5p/AMPK/PGC1α/CEBPB axis to promote stemness and chemo-resistance of gastric cancer. Cell Death Dis. 2020, 11, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; He, F.; Hou, Y.; Tu, G.; Li, Q.; Jin, T.; Zeng, H.; Qin, Y.; Wan, X.; Qiao, Y.; et al. A novel hypoxic long noncoding RNA KB-1980E6. 3 maintains breast cancer stem cell stemness via interacting with IGF2BP1 to facilitate c-Myc mRNA stability. Oncogene 2021, 40, 1609–1627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.; Wang, Y.; Lai, S.; Wang, Z.; Yang, Y.; Liu, W.; Wang, H.; Tang, B. The m6A demethylase ALKBH5-mediated upregulation of DDIT4-AS1 maintains pancreatic cancer stemness and suppresses chemosensitivity by activating the mTOR pathway. Mol. Cancer 2022, 21, 174. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, P.; Wu, Q.; Fang, H.; Wang, Y.; Xiao, Y.; Cong, M.; Wang, T.; He, Y.; Ma, C.; et al. Long non-coding RNA NR2F1-AS1 induces breast cancer lung metastatic dormancy by regulating NR2F1 and ΔNp63. Nat. Commun. 2021, 12, 5232. [Google Scholar] [CrossRef]
- Wu, A.C.; Yang, W.B.; Chang, K.Y.; Lee, J.S.; Liou, J.P.; Su, R.Y.; Cheng, S.M.; Hwang, D.Y.; Kikkawa, U.; Hsu, T.I.; et al. HDAC6 involves in regulating the lncRNA-microRNA-mRNA network to promote the proliferation of glioblastoma cells. J. Exp. Clin. Cancer Res. 2022, 41, 47. [Google Scholar] [CrossRef]
- Li, W.; Zhang, L.; Guo, B.; Deng, J.; Wu, S.; Li, F.; Wang, Y.; Lu, J.; Zhou, Y. Exosomal FMR1-AS1 facilitates maintaining cancer stem-like cell dynamic equilibrium via TLR7/NFκB/c-Myc signaling in female esophageal carcinoma. Mol. Cancer 2019, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Du, M.; Wang, S.; Zha, J.; Lei, P.; Wang, X.; Wu, D.; Zhang, J.; Chen, D.; Huang, D.; et al. Clinicopathological implication of long non-coding RNAs SOX2 overlapping transcript and its potential target gene network in various cancers. Front. Genet. 2020, 10, 1375. [Google Scholar] [CrossRef] [Green Version]
- Carracedo, A.; Cantley, L.C.; Pandolfi, P.P. Cancer metabolism: Fatty acid oxidation in the limelight. Nat. Rev. Cancer 2013, 13, 227–232. [Google Scholar] [CrossRef]
- Wang, T.; Fahrmann, J.F.; Lee, H.; Li, Y.J.; Tripathi, S.C.; Yue, C.; Zhang, C.; Lifshitz, V.; Song, J.; Yuan, Y.; et al. JAK/STAT3-regulated fatty acid β-oxidation is critical for breast cancer stem cell self-renewal and chemoresistance. Cell Metab. 2018, 27, 136–150. [Google Scholar] [CrossRef]
- Tan, Z.; Luo, X.; Xiao, L.; Tang, M.; Bode, A.M.; Dong, Z.; Cao, Y. The Role of PGC1α in Cancer Metabolism and its Therapeutic Implications. Mol. Cancer Ther. 2016, 15, 774–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Q.; Tan, Z.; Shi, F.; Tang, M.; Xie, L.; Zhao, L.; Li, Y.; Hu, J.; Zhou, M.; Bode, A.; et al. PGC1α/CEBPB/CPT1A axis promotes radiation resistance of nasopharyngeal carcinoma through activating fatty acid oxidation. Cancer Sci. 2019, 110, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, S.; Ionita-Radu, F.; Stefani, C.; Miricescu, D.; Stanescu-Spinu, I.-I.; Greabu, M.; Totan, A.R.; Jinga, M. Targeting PI3K/AKT/mTOR Signaling Pathway in Pancreatic Cancer: From Molecular to Clinical Aspects. Int. J. Mol. Sci. 2022, 23, 10132. [Google Scholar] [CrossRef] [PubMed]
- Sosa, M.S.; Parikh, F.; Maia, A.G.; Estrada, Y.; Bosch, A.; Bragado, P.; Ekpin, E.; George, A.; Zheng, Y.; Lam, H.M.; et al. NR2F1 controls tumour cell dormancy via SOX9-and RARβ-driven quiescence programmes. Nat. Commun. 2015, 6, 6170. [Google Scholar] [CrossRef] [Green Version]
- Tran, M.N.; Choi, W.; Wszolek, M.F.; Navai, N.; Lee, I.L.C.; Nitti, G.; Wen, S.; Flores, E.R.; Siefker-Radtke, A.; Czerniak, B.; et al. The p63 protein isoform ΔNp63α inhibits epithelial-mesenchymal transition in human bladder cancer cells: Role of MIR-205. J. Biol. Chem. 2013, 288, 3275–3288. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.B.; Hsu, C.C.; Hsu, T.I.; Liou, J.P.; Chang, K.Y.; Chen, P.Y.; Liu, J.J.; Yan, S.T.; Wang, J.Y.; Yeh, S.H.; et al. Increased activation of HDAC1/2/6 and Sp1 underlies therapeutic resistance and tumor growth in glioblastoma. Neuro Oncol. 2020, 22, 1439–1451. [Google Scholar] [CrossRef]
- Wahle, E.; Winkler, G.S. RNA decay machines: Deadenylation by the Ccr4–Not and Pan2–Pan3 complexes. Biochim. Biophys. Acta 2013, 1829, 561–570. [Google Scholar] [CrossRef]
- Zhang, Y.; Sui, R.; Chen, Y.; Liang, H.; Shi, J.; Piao, H. Downregulation of miR-485-3p promotes glioblastoma cell proliferation and migration via targeting RNF135. Exp. Ther. Med. 2019, 18, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Van den Boorn, J.G.; Daßler, J.; Coch, C.; Schlee, M.; Hartmann, G. Exosomes as nucleic acid nanocarriers. Adv. Drug Deliv. Rev. 2013, 65, 331–335. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, L.; Dong, L.; Wang, X. Emerging role of exosome signalling in maintaining cancer stem cell dynamic equilibrium. J. Cell. Mol. Med. 2018, 22, 3719–3728. [Google Scholar] [CrossRef]
- Qu, L.E.; Ding, J.; Chen, C.; Wu, Z.J.; Liu, B.; Gao, Y.; Chen, W.; Liu, F.; Sun, W.; Li, X.F.; et al. Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell 2016, 29, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Rhee, H.; Kim, H.Y.; Choi, J.H.; Woo, H.G.; Yoo, J.E.; Nahm, J.H.; Choi, J.S.; Park, Y.N. Keratin 19 Expression in Hepatocellular Carcinoma Is Regulated by Fibroblast-Derived HGF via a MET-ERK1/2-AP1 and SP1 AxisKRT19 in HCC Is Regulated by CAF via HGF-MET-AP1 and SP1. Cancer Res. 2018, 78, 1619–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zeng, X.; Wang, N.; Zhao, W.; Zhang, X.; Teng, S.; Zhang, Y.; Lu, Z. Long noncoding RNA DANCR, working as a competitive endogenous RNA, promotes ROCK1-mediated proliferation and metastasis via decoying of miR-335-5p and miR-1972 in osteosarcoma. Mol. Cancer 2018, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Boelens, M.C.; Wu, T.J.; Nabet, B.Y.; Xu, B.; Qiu, Y.; Yoon, T.; Azzam, D.J.; Victor, T.S.; Wiemann, B.Z.; Ishwaran, H.; et al. Exosome transfer from stromal to breast cancer cells regulates therapy resistance pathways. Cell 2014, 159, 499–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Dong, J.; Huang, J.; Ye, W.; Zheng, Z.; Huang, K.; Pan, Y.; Cen, J.; Liang, Y.; Shu, G.; et al. Chitosan-Gelatin-EGCG Nanoparticle-Meditated LncRNA TMEM44-AS1 Silencing to Activate the P53 Signaling Pathway for the Synergistic Reversal of 5-FU Resistance in Gastric Cancer. Adv. Sci. 2022, 9, e2105077. [Google Scholar] [CrossRef]
- Mao, W.; Wang, K.; Zhang, W.; Chen, S.; Xie, J.; Zheng, Z.; Li, X.; Zhang, N.; Zhang, Y.; Zhang, H.; et al. Transfection with plasmid-encoding lncRNA-SLERCC nanoparticle-mediated delivery suppressed tumor progression in renal cell carcinoma. J. Exp. Clin. Cancer Res. 2022, 41, 252. [Google Scholar] [CrossRef]
- Wang, P.L.; Teng, L.; Feng, Y.C.; Yue, Y.M.; Han, M.M.; Yan, Q.; Ye, K.; Tang, C.X.; Zhang, S.N.; Qi, T.F.; et al. The N-Myc-responsive lncRNA MILIP promotes DNA double-strand break repair through non-homologous end joining. Proc. Natl. Acad. Sci. USA 2022, 119, e2208904119. [Google Scholar] [CrossRef]
- Zhang, H.; Dai, Z.; Wu, W.; Wang, Z.; Zhang, N.; Zhang, L.; Zeng, W.J.; Liu, Z.; Cheng, Q. Regulatory mechanisms of immune checkpoints PD-L1 and CTLA-4 in cancer. J. Exp. Clin. Cancer Res. 2021, 40, 184. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, N.; Wu, W.; Zhou, R.; Li, S.; Wang, Z.; Dai, Z.; Zhang, L.; Liu, Z.; Zhang, J.; et al. Machine learning-based tumor-infiltrating immune cell-associated lncRNAs for pre-dicting prognosis and immunotherapy response in patients with glioblastoma. Brief. Bioinform. 2022, 23, bbac386. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, H.; Wu, W.; Zhou, R.; Li, S.; Wang, Z.; Dai, Z.; Zhang, L.; Liu, F.; Liu, Z.; et al. Machine learning-based identification of tumor-infiltrating immune cell-associated lncRNAs for improving outcomes and immunotherapy responses in patients with low-grade glioma. Theranostics 2022, 12, 5931–5948. [Google Scholar] [CrossRef]
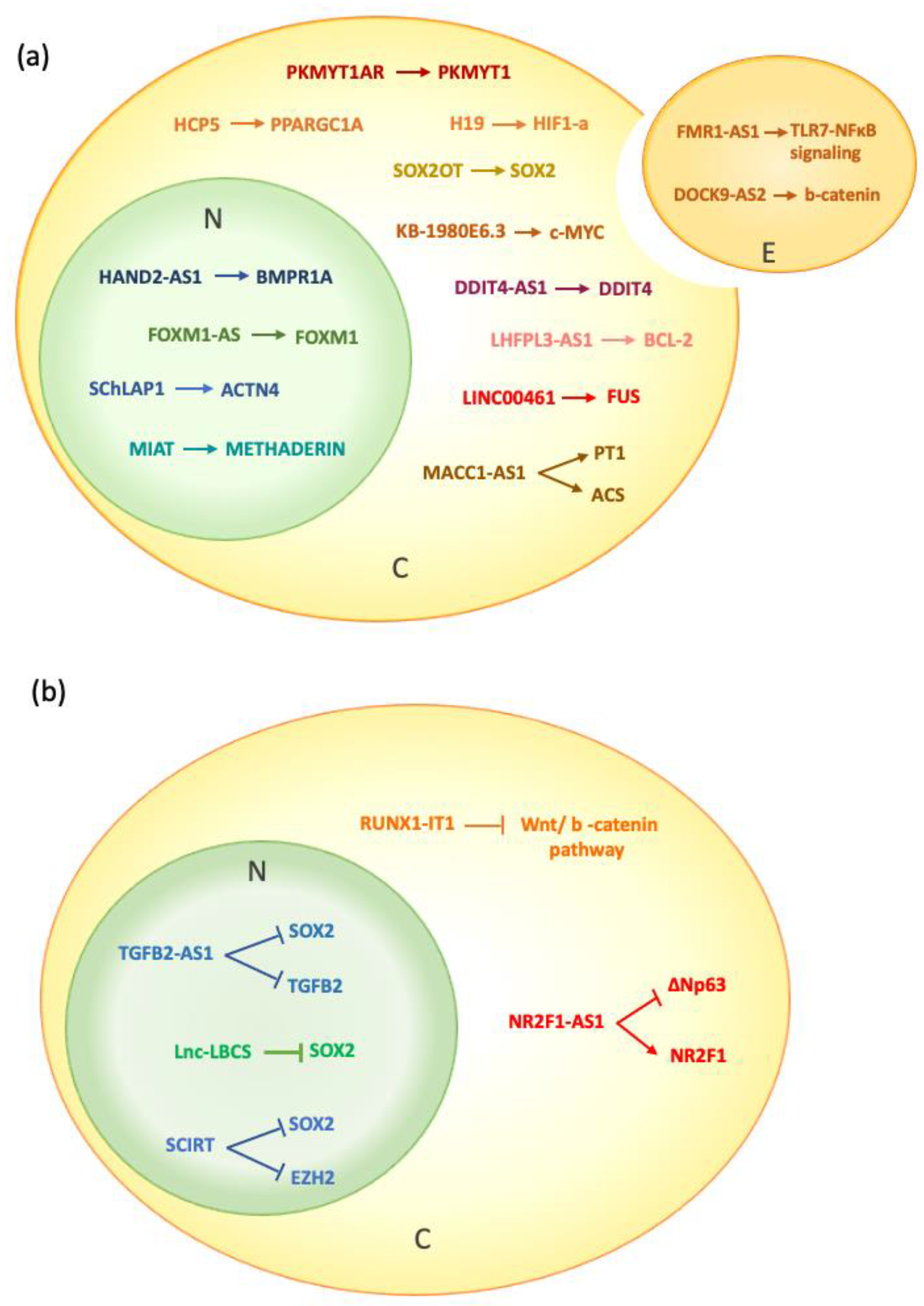

{kind=link}
| lncRNA |
Modulation Tumor Type | Molecular Pathway | Role in Cancer | Ref. |
|---|---|---|---|---|
| HAND2-AS1 | Up Liver cancer | INO80 complex on BMPR1A promoter | Oncogenic: increasing self-renewal and maintenance of liver CSCs. | [36] |
| TGFB2-AS1 | Down Triple-negative breast cancer | Antagonizing SMARCA4 binding capacity on TGFB2 and SOX2 promoters | Tumor suppressor: inhibition of CSCs signaling | [37] |
| SCIRT | Up Breast cancer | EZH2 and SOX2 colocalization and binding | Tumor suppressor: repression of the transcriptional programs of self-renewal in CSCs | [38] |
| lnc-LBCS | Down bladder cancer | Scaffold to induce the formation of hnRNPK–EZH2 complex | Tumor suppressor: Inhibition of Self-Renewal and Chemoresistance of CSCs | [39] |
| FOXM1-AS | Up Glioblastoma | ALKBH5/demethylation of FOXM1 nascent transcripts and FOXM1 expression | Oncogenic: proliferation of glioblastoma stem cells and in vivo tumorigenesis | [40] |
| SChLAP1 | Up Glioblastoma | Enhanced interaction between HNRNPL and ACTN4, increased stability of ACTN4 protein, promoted nuclear translocation of NF-kB | Oncogenic: increased proliferation of glioblastoma cells in vitro and in vivo | [41] |
| MIAT | Up Medulloblastoma | Metadherin/regulation of a set of microRNAs | Oncogenic: maintenance of the tumorigenic stem-like phenotype and resistance to radiotherapy | [42] |
| DOCK9-AS2 | Up papillary thyroid carcinoma | Increasing β-catenin level trought interaction with Sp1 transcription factor | Oncogenic: restoring proliferation, migration, invasion, EMT, and stemness of PTC cells. | [43] |
| lncRNA |
Modulation Tumor Type | Molecular Pathway | Role in Cancer | Ref. |
|---|---|---|---|---|
| H19 | Up Breast cancer | Sponging let-7/up-regulation of HIF1-α | Oncogenic: increase in the glycolysis gatekeeper PDK1 under hypoxia | [56] |
| RUNX1-IT1 | Down HCC | Sponging miR-632/ Up-regulation of GSK-3β | Tumor suppressor: repression of the Wnt/β -catenin pathway | [57] |
| DOCK9-AS2 | Up PTC | Sponging miR-1972/ Up-regulation of CTNNB1 | Oncogenic: enriched in the exosomes from PTC stem-like cells, which transfer this lncRNA to the non-stem tumor cells | [43] |
| PKMYT1AR | Up NSCLC | Sponging miR-485-5p/Up-regulation of PKMYT1 | Oncogenic: activation of Wnt/β-catenin pathway | [58] |
| SOX2OT | Up Bladder cancer | Sponging miR-200c/Up-regulation of SOX2 | Oncogenic: increase in stemness in vitro and tumorigenic and metastatic ability in vivo | [59] |
| LHFPL3-AS1 | Up Melanoma | Sponging miR-181a-5p/Up-regulation of BCL2 | Oncogenic: inhibition of the apoptosis of melanoma stem cells | [60] |
| MACC1-AS1 | Up Gastric cancer | Sponging miR-145-5p/C Up-regulation of PT1 and ACS | Oncogenic: increases FAO-dependent stemness and chemoresistance | [61] |
| HCP5 | Up Gastric cancer | SpongingmiR-3619-5p/Up-regulation of PPARGC1A | Oncogenic: increases FAO, stemness and chemoresistance | [62] |
| KB-1980E6.3 | Up Breast cancer | Enhanced recognition of m6A-modified c-Myc mRNA via IGF2BP1, increased stability of c-Myc mRNA | Oncogenic: breast cancer cells stemness and tumorigenesis in vitro and in vivo | [63] |
| DDIT4-AS1 | Up Pancreatic cancer | UPF1/increased DDIT4-AS1, promotion of UPF1 phosphorylation, degradation of DDIT4 mRNA | Oncogenic: activation of the mTOR pathway, increased stemness and reduced chemosensitivity | [64] |
| NR2F1-AS1 | Higher in late vs early recurring tumors Breast cancer | PTBP1/enhanced translation of NR2F1, suppression of the transcription factor and marker of epithelial stem cells ΔNp63 | Reduced metastatic proficiency. Promotion of the dormancy of breast cancer metastatic cells, EMT changes and impaired tumorigenicity | [65] |
| LINC00461 | Up Glioblastoma | CNOT6, FUS/LINC00461 stabilization, competitive binding to miR-485-3p | Oncogenic: increased expression of key factors promoting cell division and proliferation | [66] |
| FMR1-AS1 | Up Esophageal squamous cell carcinoma | Interaction with TLR7 to activate TLR7-NFκB signaling and c-Myc expression | Oncogenic: inducing ESCC cell proliferation and invasion ability | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciafrè, S.A.; Russo, M.; Michienzi, A.; Galardi, S. Long Noncoding RNAs and Cancer Stem Cells: Dangerous Liaisons Managing Cancer. Int. J. Mol. Sci. 2023, 24, 1828. https://doi.org/10.3390/ijms24031828
Ciafrè SA, Russo M, Michienzi A, Galardi S. Long Noncoding RNAs and Cancer Stem Cells: Dangerous Liaisons Managing Cancer. International Journal of Molecular Sciences. 2023; 24(3):1828. https://doi.org/10.3390/ijms24031828
Chicago/Turabian StyleCiafrè, Silvia Anna, Monia Russo, Alessandro Michienzi, and Silvia Galardi. 2023. "Long Noncoding RNAs and Cancer Stem Cells: Dangerous Liaisons Managing Cancer" International Journal of Molecular Sciences 24, no. 3: 1828. https://doi.org/10.3390/ijms24031828