
Circular RNAs and Untranslated Regions in Acute Myeloid Leukemia


Abstract
1. Introduction
2. CircRNAs
2.1. Cellular Localization and Mechanisms of Action of CircRNAs
2.1.1. Transcriptional/Epigenetic Regulators in the Nucleus
2.1.2. Sponge for miRNAs and Binding to mRNAs in the Cytoplasm
2.1.3. Protein Regulator in the Cytoplasm
2.1.4. Extracellular Transport via Exosomes
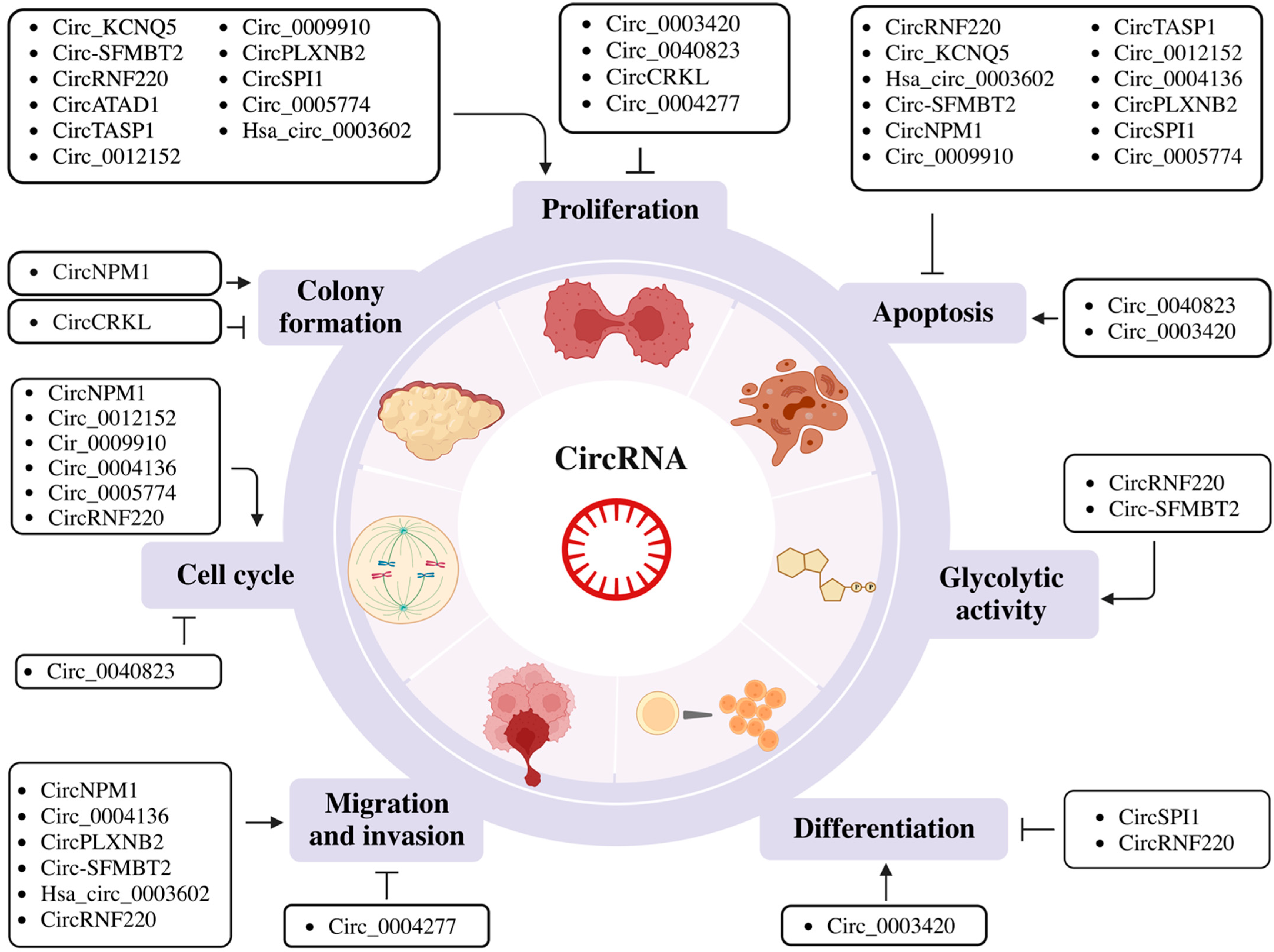
2.2. Biological Roles of CircRNAs
2.2.1. Regulation of Cell Cycle Progression, Proliferation, Apoptosis and Metabolism
2.2.2. Regulation of Cell Phenotype or Differentiation
2.2.3. Regulation of Migration and Invasion
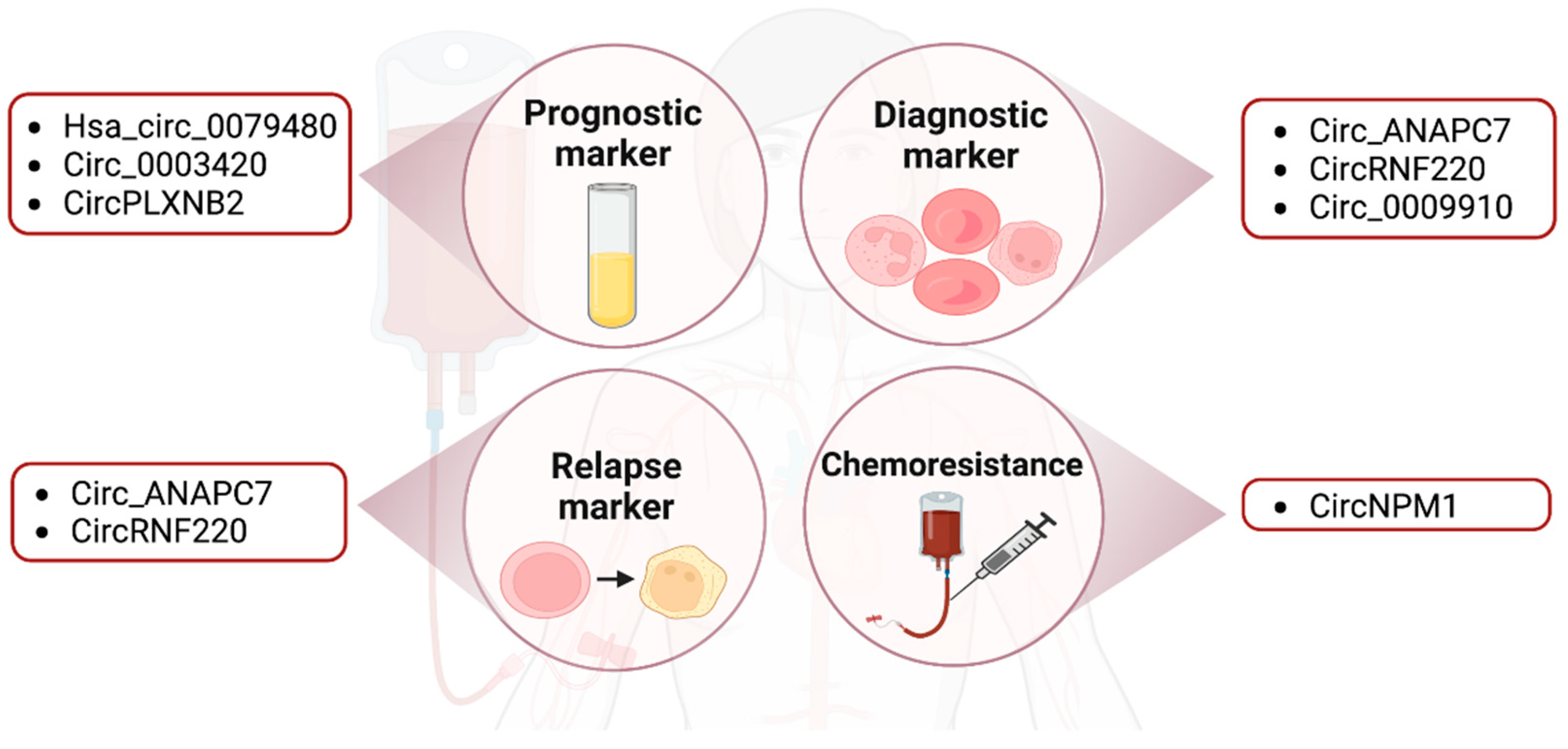
2.3. Clinical Relevance of CircRNAs
2.3.1. Prognostic, Diagnostic, Relapse and Subtype Markers
2.3.2. Drug Resistance
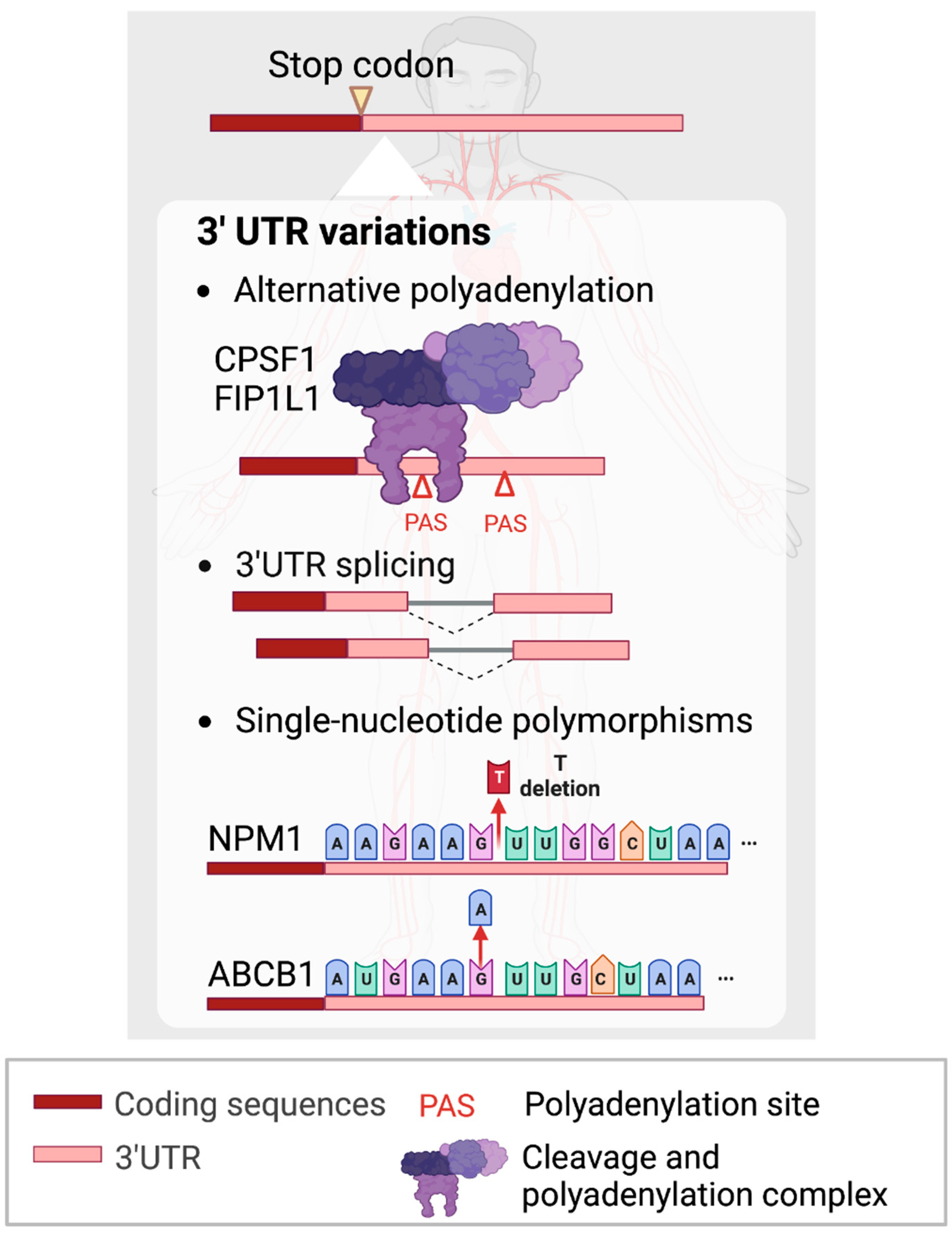
3. 3’ Untranslated Regions (3’UTRs) of mRNAs
3.1. APA within 3’UTRs
3.2. 3’UTR Splicing
3.3. SNPs within 3’UTRs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dohner, H.; Wei, A.H.; Appelbaum, F.R.; Craddock, C.; DiNardo, C.D.; Dombret, H.; Ebert, B.L.; Fenaux, P.; Godley, L.A.; Hasserjian, R.P.; et al. Diagnosis and management of AML in adults: 2022 recommendations from an international expert panel on behalf of the ELN. Blood 2022, 140, 1345–1377. [Google Scholar] [CrossRef] [PubMed]
- Olsson, I.; Bergh, G.; Ehinger, M.; Gullberg, U. Cell differentiation in acute myeloid leukemia. Eur. J. Haematol. 1996, 57, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gu, Y.; Chen, B. Mechanisms of drug resistance in acute myeloid leukemia. Onco. Targets Ther. 2019, 12, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.J.; Hourigan, C.S.; Smith, T.J. Adult Acute Myeloid Leukemia Long-term Survivors. J. Leuk. 2014, 2, 26855. [Google Scholar] [CrossRef] [PubMed]
- Megias-Vericat, J.E.; Rojas, L.; Herrero, M.J.; Boso, V.; Montesinos, P.; Moscardo, F.; Poveda, J.L.; Sanz, M.A.; Alino, S.F. Influence of ABCB1 polymorphisms upon the effectiveness of standard treatment for acute myeloid leukemia: A systematic review and meta-analysis of observational studies. Pharm. J. 2015, 15, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Luppi, M.; Fabbiano, F.; Visani, G.; Martinelli, G.; Venditti, A. Novel Agents for Acute Myeloid Leukemia. Cancers 2018, 10, 429. [Google Scholar] [CrossRef]
- Dombret, H.; Gardin, C. An update of current treatments for adult acute myeloid leukemia. Blood 2016, 127, 53–61. [Google Scholar] [CrossRef]
- Alotaibi, A.S.; Yilmaz, M.; Kanagal-Shamanna, R.; Loghavi, S.; Kadia, T.M.; DiNardo, C.D.; Borthakur, G.; Konopleva, M.; Pierce, S.A.; Wang, S.A.; et al. Patterns of Resistance Differ in Patients with Acute Myeloid Leukemia Treated with Type I versus Type II FLT3 inhibitors. Blood Cancer Discov. 2021, 2, 125–134. [Google Scholar] [CrossRef]
- Bennett, J.M.; Catovsky, D.; Daniel, M.T.; Flandrin, G.; Galton, D.A.; Gralnick, H.R.; Sultan, C. Proposed revised criteria for the classification of acute myeloid leukemia. A report of the French-American-British Cooperative Group. Ann. Intern. Med. 1985, 103, 620–625. [Google Scholar] [CrossRef]
- Arber, D.A.; Brunning, R.D.; Le Beau, M.M. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, Revised 4th ed.; Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Eds.; IARC Press: Lyon, France, 2017; pp. 130–171, ISBN-13 978-928-324-494-3. [Google Scholar]
- Hwang, S.M. Classification of acute myeloid leukemia. Blood Res. 2020, 55, S1–S4. [Google Scholar] [CrossRef]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.Y.; Sun, H.S.; Tsai, S.J. Circular RNA—New member of noncoding RNA with novel functions. Exp. Biol. Med. 2017, 242, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, C.; Du, Y.; Li, Z.; Li, M.; Hou, P.; Shen, Z.; Chu, S.; Zheng, J.; Bai, J. Expanding uncapped translation and emerging function of circular RNA in carcinomas and noncarcinomas. Mol. Cancer 2022, 21, 13. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, M.; Zhang, X.; Huang, F.; Wu, K.; Zhang, J.; Liu, J.; Huang, Z.; Luo, H.; Tao, L.; et al. Cellular microRNAs up-regulate transcription via interaction with promoter TATA-box motifs. RNA 2014, 20, 1878–1889. [Google Scholar] [CrossRef]
- Wallace, J.A.; O’Connell, R.M. MicroRNAs and acute myeloid leukemia: Therapeutic implications and emerging concepts. Blood 2017, 130, 1290–1301. [Google Scholar] [CrossRef]
- Guo, B.; Li, D.; Du, L.; Zhu, X. piRNAs: Biogenesis and their potential roles in cancer. Cancer Metastasis Rev. 2020, 39, 567–575. [Google Scholar] [CrossRef]
- Katayama, S.; Tomaru, Y.; Kasukawa, T.; Waki, K.; Nakanishi, M.; Nakamura, M.; Nishida, H.; Yap, C.C.; Suzuki, M.; Kawai, J.; et al. Antisense transcription in the mammalian transcriptome. Science 2005, 309, 1564–1566. [Google Scholar] [CrossRef]
- Li, W.; Notani, D.; Rosenfeld, M.G. Enhancers as non-coding RNA transcription units: Recent insights and future perspectives. Nat. Rev. Genet. 2016, 17, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Pelechano, V.; Steinmetz, L.M. Gene regulation by antisense transcription. Nat. Rev. Genet. 2013, 14, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Xiong, F.; Li, W. Enhancer RNAs in cancer: Regulation, mechanisms and therapeutic potential. RNA Biol. 2020, 17, 1550–1559. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, Z.; Pang, Y.; Cui, L.; Qian, T.; Quan, L.; Zhao, H.; Shi, J.; Ke, X.; Fu, L. Role of microRNAs, circRNAs and long noncoding RNAs in acute myeloid leukemia. J. Hematol. Oncol. 2019, 12, 51. [Google Scholar] [CrossRef]
- Anelli, L.; Zagaria, A.; Specchia, G.; Musto, P.; Albano, F. Dysregulation of miRNA in Leukemia: Exploiting miRNA Expression Profiles as Biomarkers. Int. J. Mol. Sci. 2021, 22, 7156. [Google Scholar] [CrossRef]
- Mayr, C.; Bartel, D.P. Widespread shortening of 3’UTRs by alternative cleavage and polyadenylation activates oncogenes in cancer cells. Cell 2009, 138, 673–684. [Google Scholar] [CrossRef]
- Chan, J.J.; Zhang, B.; Chew, X.H.; Salhi, A.; Kwok, Z.H.; Lim, C.Y.; Desi, N.; Subramaniam, N.; Siemens, A.; Kinanti, T.; et al. Pan-cancer pervasive upregulation of 3’ UTR splicing drives tumourigenesis. Nat. Cell Biol 2022, 24, 928–939. [Google Scholar] [CrossRef]
- Yuan, F.; Hankey, W.; Wagner, E.J.; Li, W.; Wang, Q. Alternative polyadenylation of mRNA and its role in cancer. Genes Dis. 2021, 8, 61–72. [Google Scholar] [CrossRef]
- Brandsma, A.M.; Bertrums, E.J.M.; van Roosmalen, M.J.; Hofman, D.A.; Oka, R.; Verheul, M.; Manders, F.; Ubels, J.; Belderbos, M.E.; van Boxtel, R. Mutation signatures of pediatric acute myeloid leukemia and normal blood progenitors associated with differential patient outcomes. Blood Cancer Discov. 2021, 2, 484–499. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.L.; Beesley, J.; French, J.D.; Dunning, A.M. Beyond GWASs: Illuminating the dark road from association to function. Am. J. Hum. Genet. 2013, 93, 779–797. [Google Scholar] [CrossRef] [PubMed]
- Hrdlickova, B.; de Almeida, R.C.; Borek, Z.; Withoff, S. Genetic variation in the non-coding genome: Involvement of micro-RNAs and long non-coding RNAs in disease. Biochim. Biophys. Acta 2014, 1842, 1910–1922. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.F.; Saetrom, P. Single nucleotide polymorphisms can create alternative polyadenylation signals and affect gene expression through loss of microRNA-regulation. PLoS Comput. Biol. 2012, 8, e1002621. [Google Scholar] [CrossRef]
- Chen, M.; Lyu, G.; Han, M.; Nie, H.; Shen, T.; Chen, W.; Niu, Y.; Song, Y.; Li, X.; Li, H.; et al. 3’ UTR lengthening as a novel mechanism in regulating cellular senescence. Genome Res. 2018, 28, 285–294. [Google Scholar] [CrossRef]
- Mularoni, L.; Sabarinathan, R.; Deu-Pons, J.; Gonzalez-Perez, A.; Lopez-Bigas, N. OncodriveFML: A general framework to identify coding and non-coding regions with cancer driver mutations. Genome Biol. 2016, 17, 128. [Google Scholar] [CrossRef]
- Weinhold, N.; Jacobsen, A.; Schultz, N.; Sander, C.; Lee, W. Genome-wide analysis of noncoding regulatory mutations in cancer. Nat. Genet. 2014, 46, 1160–1165. [Google Scholar] [CrossRef]
- Puente, X.S.; Bea, S.; Valdes-Mas, R.; Villamor, N.; Gutierrez-Abril, J.; Martin-Subero, J.I.; Munar, M.; Rubio-Perez, C.; Jares, P.; Aymerich, M.; et al. Non-coding recurrent mutations in chronic lymphocytic leukaemia. Nature 2015, 526, 519–524. [Google Scholar] [CrossRef]
- Zhang, Y.; Xue, W.; Li, X.; Zhang, J.; Chen, S.; Zhang, J.L.; Yang, L.; Chen, L.L. The Biogenesis of Nascent Circular RNAs. Cell Rep. 2016, 15, 611–624. [Google Scholar] [CrossRef]
- Chen, I.; Chen, C.Y.; Chuang, T.J. Biogenesis, identification, and function of exonic circular RNAs. Wiley Interdiscip. Rev. RNA 2015, 6, 563–579. [Google Scholar] [CrossRef]
- Perez de Acha, O.; Rossi, M.; Gorospe, M. Circular RNAs in Blood Malignancies. Front. Mol. Biosci. 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Bonizzato, A.; Gaffo, E.; Te Kronnie, G.; Bortoluzzi, S. CircRNAs in hematopoiesis and hematological malignancies. Blood Cancer J. 2016, 6, e483. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.; Wang, Y.; Li, Z.; Zhang, M. Role of circular RNA in hematological malignancies. Oncol. Lett. 2019, 18, 4385–4392. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Uddin, M.H.; Zonder, J.A.; Azmi, A.S.; Balasubramanian, S.K. Circular RNAs in acute myeloid leukemia. Mol. Cancer 2021, 20, 149. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, B.; Qi, X.; Bai, L.; Li, B.; Bao, H.; Wu, X.; Wu, X.; Zhao, Y. Circular RNA ATAD1 is upregulated in acute myeloid leukemia and promotes cancer cell proliferation by downregulating miR-34b via promoter methylation. Oncol. Lett. 2021, 22, 799. [Google Scholar] [CrossRef]
- Lin, L.; Wang, Y.; Bian, S.; Sun, L.; Guo, Z.; Kong, D.; Zhao, L.; Guo, D.; Li, Q.; Wu, M.; et al. A circular RNA derived from PLXNB2 as a valuable predictor of the prognosis of patients with acute myeloid leukaemia. J. Transl. Med. 2021, 19, 123. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, S.; Yin, J.; Yu, W.; Xu, C. CircRNF220 plays a pathogenic role to facilitate cell progression of AML in vitro via sponging miR-330-5p to induce upregulation of SOX4. Histol. Histopathol. 2022, 37, 18472. [Google Scholar]
- Hu, X.; Yin, J.; He, R.; Chao, R.; Zhu, S. Circ_KCNQ5 participates in the progression of childhood acute myeloid leukemia by enhancing the expression of RAB10 via binding to miR-622. Hematology 2022, 27, 431–440. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Liu, J.; Jin, Y.; Wang, W. Circular RNA circ_0004277 Inhibits Acute Myeloid Leukemia Progression Through MicroRNA-134-5p / Single stranded DNA binding protein 2. Bioengineered 2022, 13, 9662–9673. [Google Scholar] [CrossRef]
- Ye, Q.; Li, N.; Zhou, K.; Liao, C. Homo sapiens circular RNA 0003602 (Hsa_circ_0003602) accelerates the tumorigenicity of acute myeloid leukemia by modulating miR-502-5p/IGF1R axis. Mol. Cell. Biochem. 2022, 477, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yang, B.; Jin, J.; He, Y.; Wu, X.; Yang, Y.; Zhou, W.; He, Z. Circular RNA circ_0040823 inhibits the proliferation of acute myeloid leukemia cells and induces apoptosis by regulating miR-516b/PTEN. J. Gene Med. 2022, 24, e3404. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Shang, Z.; Ming, X.; Wu, J.; Xiao, Y. Circ-SFMBT2 facilitates the malignant growth of acute myeloid leukemia cells by modulating miR-582-3p/ZBTB20 pathway. Histol. Histopathol. 2022, 37, 137–149. [Google Scholar] [PubMed]
- Ding, J.; Zhang, X.; Xue, J.; Fang, L.; Ban, C.; Song, B.; Wu, L. CircNPM1 strengthens Adriamycin resistance in acute myeloid leukemia by mediating the miR-345-5p/FZD5 pathway. Cent. Eur. J. Immunol. 2021, 46, 162–182. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, B.; Chen, X.; Geng, X.; Zhang, Z. Circ_0009910 sponges miR-491-5p to promote acute myeloid leukemia progression through modulating B4GALT5 expression and PI3K/AKT signaling pathway. Int. J. Lab. Hematol. 2022, 44, 320–332. [Google Scholar] [CrossRef]
- Liu, X.; Liu, X.; Cai, M.; Luo, A.; He, Y.; Liu, S.; Zhang, X.; Yang, X.; Xu, L.; Jiang, H. CircRNF220, not its linear cognate gene RNF220, regulates cell growth and is associated with relapse in pediatric acute myeloid leukemia. Mol. Cancer 2021, 20, 139. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, F. Circular RNA circCRKL inhibits the proliferation of acute myeloid leukemia cells via the miR-196a-5p/miR-196b-5p/p27 axis. Bioengineered 2021, 12, 7704–7713. [Google Scholar] [CrossRef]
- Shang, Z.; Ming, X.; Wu, J.; Xiao, Y. Downregulation of circ_0012152 inhibits proliferation and induces apoptosis in acute myeloid leukemia cells through the miR-625-5p/SOX12 axis. Hematol. Oncol. 2021, 39, 539–548. [Google Scholar] [CrossRef]
- Wang, D.; Ming, X.; Xu, J.; Xiao, Y. Circ_0009910 shuttled by exosomes regulates proliferation, cell cycle and apoptosis of acute myeloid leukemia cells by regulating miR-5195-3p/GRB10 axis. Hematol. Oncol. 2021, 39, 390–400. [Google Scholar] [CrossRef]
- Bi, J.; Pu, Y.; Yu, X. Exosomal circ_0004136 enhances the progression of pediatric acute myeloid leukemia depending on the regulation of miR-570-3p/TSPAN3 axis. Anticancer Drugs 2021, 32, 802–811. [Google Scholar] [CrossRef]
- Wang, X.; Jin, P.; Zhang, Y.; Wang, K. CircSPI1 acts as an oncogene in acute myeloid leukemia through antagonizing SPI1 and interacting with microRNAs. Cell Death Dis. 2021, 12, 297. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Luan, Q.; Zhu, H.; Zhao, Y.; Ji, J.; Wu, F.; Yan, J. Circular RNA circ_0005774 contributes to proliferation and suppresses apoptosis of acute myeloid leukemia cells via circ_0005774/miR-192-5p/ULK1 ceRNA pathway. Biochem. Biophys. Res. Commun. 2021, 551, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Fei, Y.; Zhang, Y. Hsa-circ_0003420 induces apoptosis in acute myeloid leukemia stem cells and impairs stem cell properties. Immunopharmacol. Immunotoxicol. 2021, 43, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, Y.; Liang, C.; Xie, S.; Xie, A. Silencing of circTASP1 inhibits proliferation and induces apoptosis of acute myeloid leukaemia cells through modulating miR-515-5p/HMGA2 axis. J. Cell. Mol. Med. 2021, 25, 7367–7380. [Google Scholar] [CrossRef]
- Guo, L.; Kou, R.; Song, Y.; Li, G.; Jia, X.; Li, Z.; Zhang, Y. Serum hsa_circ_0079480 is a novel prognostic marker for acute myeloid leukemia. J. Clin. Lab. Anal. 2022, 36, e24337. [Google Scholar] [CrossRef]
- Shen, Y.; Jia, Y.; Zhang, R.; Chen, H.; Feng, Y.; Li, F.; Wang, T.; Bai, J.; He, A.; Yang, Y. Using Circ-ANAPC7 as a Novel Type of Biomarker in the Monitoring of Acute Myeloid Leukemia. Acta Haematol. 2022, 145, 176–183. [Google Scholar] [CrossRef]
- Liu, Y.C.; Li, J.R.; Sun, C.H.; Andrews, E.; Chao, R.F.; Lin, F.M.; Weng, S.L.; Hsu, S.D.; Huang, C.C.; Cheng, C.; et al. CircNet: A database of circular RNAs derived from transcriptome sequencing data. Nucleic Acids Res. 2016, 44, D209–D215. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Q.; Shen, J.; Yang, B.B.; Ding, X. Circbank: A comprehensive database for circRNA with standard nomenclature. RNA Biol. 2019, 16, 899–905. [Google Scholar] [CrossRef]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef]
- Lei, P.; Chen, J.J.; Liao, C.S.; Liu, G.H.; Zhou, M. Silencing of circ_0009910 inhibits acute myeloid leukemia cell growth through increasing miR-20a-5p. Blood Cells Mol. Dis. 2019, 75, 41–47. [Google Scholar]
- Yuan, D.M.; Ma, J.; Fang, W.B. Identification of non-coding RNA regulatory networks in pediatric acute myeloid leukemia reveals circ-0004136 could promote cell proliferation by sponging miR-142. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9251–9258. [Google Scholar] [PubMed]
- Hirsch, S.; Blatte, T.J.; Grasedieck, S.; Cocciardi, S.; Rouhi, A.; Jongen-Lavrencic, M.; Paschka, P.; Kronke, J.; Gaidzik, V.I.; Dohner, H.; et al. Circular RNAs of the nucleophosmin (NPM1) gene in acute myeloid leukemia. Haematologica 2017, 102, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs Co-Precipitate with Extracellular Vesicles: A Possible Mechanism for circRNA Clearance. PLoS ONE 2016, 11, e0148407. [Google Scholar] [CrossRef]
- Milane, L.; Singh, A.; Mattheolabakis, G.; Suresh, M.; Amiji, M.M. Exosome mediated communication within the tumor microenvironment. J. Control. Release 2015, 219, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Lei, K.; Huang, F.; Jiang, Z.; Zhou, X. Exo-circRNAs: A new paradigm for anticancer therapy. Mol. Cancer 2019, 18, 56. [Google Scholar] [CrossRef]
- Seimiya, T.; Otsuka, M.; Iwata, T.; Shibata, C.; Tanaka, E.; Suzuki, T.; Koike, K. Emerging Roles of Exosomal Circular RNAs in Cancer. Front. Cell Dev. Biol. 2020, 8, 568366. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, F.; Jia, R.; Chang, H.; Li, H.; Miao, M.; Wang, H.; Yang, Z. MiR-570 inhibits cell proliferation and glucose metabolism by targeting IRS1 and IRS2 in human chronic myelogenous leukemia. Iran. J. Basic Med. Sci. 2017, 20, 481–488. [Google Scholar]
- Kwon, H.Y.; Bajaj, J.; Ito, T.; Blevins, A.; Konuma, T.; Weeks, J.; Lytle, N.K.; Koechlein, C.S.; Rizzieri, D.; Chuah, C.; et al. Tetraspanin 3 Is Required for the Development and Propagation of Acute Myelogenous Leukemia. Cell Stem Cell 2015, 17, 152–164. [Google Scholar] [CrossRef]
- Deguchi, K.; Gilliland, D.G. Cooperativity between mutations in tyrosine kinases and in hematopoietic transcription factors in AML. Leukemia 2002, 16, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Riccioni, R. Deregulation of apoptosis in acute myeloid leukemia. Haematologica 2007, 92, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Lehuede, C.; Dupuy, F.; Rabinovitch, R.; Jones, R.G.; Siegel, P.M. Metabolic Plasticity as a Determinant of Tumor Growth and Metastasis. Cancer Res. 2016, 76, 5201–5208. [Google Scholar] [CrossRef] [PubMed]
- Kreitz, J.; Schonfeld, C.; Seibert, M.; Stolp, V.; Alshamleh, I.; Oellerich, T.; Steffen, B.; Schwalbe, H.; Schnutgen, F.; Kurrle, N.; et al. Metabolic Plasticity of Acute Myeloid Leukemia. Cells 2019, 8, 805. [Google Scholar] [CrossRef] [PubMed]
- Hiller, K.; Metallo, C.M. Profiling metabolic networks to study cancer metabolism. Curr. Opin. Biotechnol. 2013, 24, 60–68. [Google Scholar] [CrossRef]
- Ganapathy-Kanniappan, S.; Geschwind, J.F. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef]
- Yan, D.; Franzini, A.; Pomicter, A.D.; Halverson, B.J.; Antelope, O.; Mason, C.C.; Ahmann, J.M.; Senina, A.V.; Vellore, N.A.; Jones, C.L.; et al. Sirt5 Is a Druggable Metabolic Vulnerability in Acute Myeloid Leukemia. Blood Cancer Discov. 2021, 2, 266–287. [Google Scholar] [CrossRef]
- Schnerch, D.; Yalcintepe, J.; Schmidts, A.; Becker, H.; Follo, M.; Engelhardt, M.; Wasch, R. Cell cycle control in acute myeloid leukemia. Am. J. Cancer Res. 2012, 2, 508–528. [Google Scholar]
- Carroll, M.; Zhu, Y.; D’Andrea, A.D. Erythropoietin-induced cellular differentiation requires prolongation of the G1 phase of the cell cycle. Proc. Natl. Acad. Sci. USA 1995, 92, 2869–2873. [Google Scholar] [CrossRef]
- Chen, H.; Liu, T.; Liu, J.; Feng, Y.; Wang, B.; Wang, J.; Bai, J.; Zhao, W.; Shen, Y.; Wang, X.; et al. Circ-ANAPC7 is Upregulated in Acute Myeloid Leukemia and Appears to Target the MiR-181 Family. Cell Physiol. Biochem. 2018, 47, 1998–2007. [Google Scholar] [CrossRef]
- Shang, J.; Chen, W.M.; Liu, S.; Wang, Z.H.; Wei, T.N.; Chen, Z.Z.; Wu, W.B. CircPAN3 contributes to drug resistance in acute myeloid leukemia through regulation of autophagy. Leuk. Res. 2019, 85, 106198. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Chen, W.M.; Wang, Z.H.; Wei, T.N.; Chen, Z.Z.; Wu, W.B. CircPAN3 mediates drug resistance in acute myeloid leukemia through the miR-153-5p/miR-183-5p-XIAP axis. Exp. Hematol. 2019, 70, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Beltran, M.; Damizia, M.; Grelloni, C.; Colantoni, A.; Setti, A.; Di Timoteo, G.; Dattilo, D.; Centron-Broco, A.; Nicoletti, C.; et al. Circular RNA ZNF609/CKAP5 mRNA interaction regulates microtubule dynamics and tumorigenicity. Mol. Cell 2022, 82, 75–89.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. The characteristics of circRNA as competing endogenous RNA in pathogenesis of acute myeloid leukemia. BMC Cancer 2021, 21, 277. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Donehower, L.A.; Cooper, T.A.; Neilson, J.R.; Wheeler, D.A.; Wagner, E.J.; Li, W. Dynamic analyses of alternative polyadenylation from RNA-seq reveal a 3’-UTR landscape across seven tumour types. Nat. Commun. 2014, 5, 5274. [Google Scholar] [CrossRef]
- Xue, Z.; Warren, R.L.; Gibb, E.A.; MacMillan, D.; Wong, J.; Chiu, R.; Hammond, S.A.; Yang, C.; Nip, K.M.; Ennis, C.A.; et al. Recurrent tumor-specific regulation of alternative polyadenylation of cancer-related genes. BMC Genom. 2018, 19, 536. [Google Scholar] [CrossRef]
- Kim, N.; Chung, W.; Eum, H.H.; Lee, H.O.; Park, W.Y. Alternative polyadenylation of single cells delineates cell types and serves as a prognostic marker in early stage breast cancer. PLoS ONE 2019, 14, e0217196. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, L.; Shi, Q.; Zhao, G.; Wang, F. Comprehensive Analysis of APA Events and Their Association With Tumor Microenvironment in Lung Adenocarcinoma. Front. Genet. 2021, 12, 645360. [Google Scholar] [CrossRef]
- Rejeski, K.; Duque-Afonso, J.; Lubbert, M. AML1/ETO and its function as a regulator of gene transcription via epigenetic mechanisms. Oncogene 2021, 40, 5665–5676. [Google Scholar] [CrossRef]
- Yang, E.; Guan, W.; Gong, D.; Gao, X.; Han, C.; Zhang, J.; Wang, H.; Wang, M.; Li, Y.; Yu, L. Epigenetic silencing of miR564 contributes to the leukemogenesis of t(8;21) acute myeloid leukemia. Clin. Sci. 2020, 134, 3079–3091. [Google Scholar] [CrossRef]
- Tijchon, E.; Yi, G.; Mandoli, A.; Smits, J.G.A.; Ferrari, F.; Heuts, B.M.H.; Wijnen, F.; Kim, B.; Janssen-Megens, E.M.; Schuringa, J.J.; et al. The acute myeloid leukemia associated AML1-ETO fusion protein alters the transcriptome and cellular progression in a single-oncogene expressing in vitro induced pluripotent stem cell based granulocyte differentiation model. PLoS ONE 2019, 14, e0226435. [Google Scholar] [CrossRef]
- Shima, T.; Davis, A.G.; Johnson, D.T.; Kochi, Y.; Miyauchi, S.; Stoner, S.A.; Yuda, J.; Miyamoto, T.; Zhou, J.-H.; Ball, E.D. CPSF1-Mediated Acute Myeloid Leukemia Fusion 3’UTR Alternation Is a Potential Therapeutic Target. Blood 2018, 132, 885. [Google Scholar] [CrossRef]
- Davis, A.G.; Johnson, D.T.; Zheng, D.; Wang, R.; Jayne, N.D.; Liu, M.; Shin, J.; Wang, L.; Stoner, S.A.; Zhou, J.H.; et al. Alternative polyadenylation dysregulation contributes to the differentiation block of acute myeloid leukemia. Blood 2022, 139, 424–438. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Zhou, H.; Fan, H.; Yuan, Y. Single nucleotide polymorphisms and cancer susceptibility. Oncotarget 2017, 8, 110635–110649. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.; Maharry, K.; Radmacher, M.D.; Mrozek, K.; Metzeler, K.H.; Whitman, S.P.; Schwind, S.; Kohlschmidt, J.; Wu, Y.Z.; Powell, B.L.; et al. Clinical outcome and gene- and microRNA-expression profiling according to the Wilms tumor 1 (WT1) single nucleotide polymorphism rs16754 in adult de novo cytogenetically normal acute myeloid leukemia: A Cancer and Leukemia Group B study. Haematologica 2011, 96, 1488–1495. [Google Scholar] [CrossRef]
- Damm, F.; Heuser, M.; Morgan, M.; Yun, H.; Grosshennig, A.; Gohring, G.; Schlegelberger, B.; Dohner, K.; Ottmann, O.; Lubbert, M.; et al. Single nucleotide polymorphism in the mutational hotspot of WT1 predicts a favorable outcome in patients with cytogenetically normal acute myeloid leukemia. J. Clin. Oncol. 2010, 28, 578–585. [Google Scholar] [CrossRef]
- Dohner, K.; Schlenk, R.F.; Habdank, M.; Scholl, C.; Rucker, F.G.; Corbacioglu, A.; Bullinger, L.; Frohling, S.; Dohner, H. Mutant nucleophosmin (NPM1) predicts favorable prognosis in younger adults with acute myeloid leukemia and normal cytogenetics: Interaction with other gene mutations. Blood 2005, 106, 3740–3746. [Google Scholar] [CrossRef]
- Chou, W.C.; Tang, J.L.; Lin, L.I.; Yao, M.; Tsay, W.; Chen, C.Y.; Wu, S.J.; Huang, C.F.; Chiou, R.J.; Tseng, M.H.; et al. Nucleophosmin mutations in de novo acute myeloid leukemia: The age-dependent incidences and the stability during disease evolution. Cancer Res. 2006, 66, 3310–3316. [Google Scholar] [CrossRef]
- Cheng, C.K.; Kwan, T.K.; Cheung, C.Y.; Ng, K.; Liang, P.; Cheng, S.H.; Chan, N.P.; Ip, R.K.; Wong, R.S.; Lee, V.; et al. A polymorphism in the 3’-untranslated region of the NPM1 gene causes illegitimate regulation by microRNA-337-5p and correlates with adverse outcome in acute myeloid leukemia. Haematologica 2013, 98, 913–917. [Google Scholar] [CrossRef]
- Bajuaifer, N.; Grundy, M.; Hills, R.; Russell, N.H.; Pallis, M.; Seedhouse, C. A polymorphism in the 3’UTR region of ABCB1 is associated with increased allele activity with corresponding increases in P-glycoprotein expression and function. Blood 2010, 116, 3990. [Google Scholar] [CrossRef]
- Gao, F.; Dong, W.; Yang, W.; Liu, J.; Zheng, Z.; Sun, K. Expression of P-gp in acute myeloid leukemia and the reversal function of As2O3 on drug resistance. Oncol. Lett. 2015, 9, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Broxterman, H.J.; Sonneveld, P.; van Putten, W.J.; Lankelma, J.; Eekman, C.A.; Ossenkoppele, G.J.; Pinedo, H.M.; Lowenberg, B.; Schuurhuis, G.J. P-glycoprotein in primary acute myeloid leukemia and treatment outcome of idarubicin/cytosine arabinoside-based induction therapy. Leukemia 2000, 14, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Pamudurti, N.R.; Patop, I.L.; Krishnamoorthy, A.; Ashwal-Fluss, R.; Bartok, O.; Kadener, S. An in vivo strategy for knockdown of circular RNAs. Cell Discov. 2020, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Guarnerio, J.; Zhang, Y.; Cheloni, G.; Panella, R.; Mae Katon, J.; Simpson, M.; Matsumoto, A.; Papa, A.; Loretelli, C.; Petri, A.; et al. Intragenic antagonistic roles of protein and circRNA in tumorigenesis. Cell Res. 2019, 29, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Xue, W.; Zhang, L.; Yang, L.Z.; Cao, S.M.; Lei, Y.N.; Liu, C.X.; Guo, S.K.; Shan, L.; et al. Screening for functional circular RNAs using the CRISPR-Cas13 system. Nat. Methods 2021, 18, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.X.; Chen, L.L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016–2034. [Google Scholar] [CrossRef] [PubMed]
- Thol, F.; Ganser, A. Treatment of Relapsed Acute Myeloid Leukemia. Curr. Treat. Options Oncol. 2020, 21, 66. [Google Scholar] [CrossRef]
- Romine, K.A.; Nechiporuk, T.; Bottomly, D.; Jeng, S.; McWeeney, S.K.; Kaempf, A.; Corces, M.R.; Majeti, R.; Tyner, J.W. Monocytic differentiation and AHR signaling as Primary Nodes of BET Inhibitor Response in Acute Myeloid Leukemia. Blood Cancer Discov. 2021, 2, 518–531. [Google Scholar] [CrossRef]
- Wang, V.E.; Blaser, B.W.; Patel, R.K.; Behbehani, G.K.; Rao, A.A.; Durbin-Johnson, B.; Jiang, T.; Logan, A.C.; Settles, M.; Mannis, G.N.; et al. Inhibition of MET Signaling with Ficlatuzumab in Combination with Chemotherapy in Refractory AML: Clinical Outcomes and High-Dimensional Analysis. Blood Cancer Discov. 2021, 2, 434–449. [Google Scholar] [CrossRef]
- Winer, E.S.; Stone, R.M. Novel therapy in Acute myeloid leukemia (AML): Moving toward targeted approaches. Ther. Adv. Hematol. 2019, 10, 2040620719860645. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Non-Coding RNA | Host Gene | Localization | Sample and Cell Lines/Expression Pattern | Target miRNA/mRNA or Protein | Function | Ref. |
|---|---|---|---|---|---|---|
| CircATAD1 | ATAD1 | Nucleus | Up in BM | miR-34b | Induces proliferation and miR-34b methylation. | [47] |
| CircPLXNB2 | PLXNB2 | Nucleus | Up in AML patients, OCI-AML3 (Not in HL-60, THP1, MV4-11) | Prognostic marker. Induces cell proliferation and migration. Inhibits apoptosis. | [48] | |
| CircRNF220 | Ring Finger Protein 220 | Cytoplasm | Up in serum, THP-1 MV4-11, KG-1, HL-60 and AML2 | miR-330-5p/SOX4 | Induces cell progression, invasion and glycolytic metabolism. Inhibits apoptosis. | [49] |
| Circ_KCNQ5 | KCNQ5 | Cytoplasm | Up in BM from childhood patients, Kasumi-1 and MV4-11 | miR-622/RAB10 | Induces proliferation and inhibits cell apoptosis. | [50] |
| Circ_0004277 | Cytoplasm | Down in BM, THP-1, K-562, U937 and KG-1a | miR-134-5p/SSBP2 | Inhibits cell viability, migration and invasion. | [51] | |
| Hsa_circ_0003602 | Up in BM, HL60, K-562 and THP-1 | miR-502-5p/IGF1R | Induces cell proliferation, migration and invasion. Inhibits apoptosis. | [52] | ||
| Circ_0040823 | Down in peripheral blood, HL-60, THP-1, U937 and KG-1 | miR-516b/PTEN | Inhibits proliferation. Induces apoptosis and cell cycle arrest. | [53] | ||
| Circ-SFMBT2 | SFMBT2 | Cytoplasm | Up in serum, U937, NB4, K-562 and HL-60 | miR-582-3p/ZBTB20 | Induces proliferation, migration, invasion and glycolysis. Inhibits apoptosis. | [54] |
| CircNPM1 | NPM1 | Cytoplasm | Up in serum, THP-1 and HL-60 | miR-345-5p/FZD5 | Induces colony formation, migration, invasion and Adriamycin resistance. Inhibits apoptosis and cell cycle arrest. | [55] |
| Circ_0009910 | MFN2 | Up in BM, HL60 and MOLM-13 | miR-491-5P/B4GALT5 and PI3K/AKT signaling pathway | Induces proliferation, sphere formation and autophagy. Inhibits apoptosis. | [56] | |
| CircRNF220 | RNF220 | Cytoplasm | Up in BM of pediatric patients and primary AML cells | miR-30a | Diagnosis, prognosis and relapse markers. Induces proliferation. Inhibits apoptosis and regulates cell differentiation. | [57] |
| CircCRKL | CRKL | Cytoplasm | Down in BM, KG-1a, KG-1, Kasumi-1, THP1, MOLM-13 | miR-196a-5p and miR196b-5p/p27 | Inhibits proliferation and colony-forming ability. | [58] |
| Circ_0012152 | RNF220 | Up in BM, U937 and HL60 cells | miR-625-5p/SOX12 | Induces proliferation. Inhibits apoptosis and regulates cell cycle. | [59] | |
| Cir_0009910 | Cytoplasm and shuttled by exosome | Up in BM, HL-60 and MOLM-13 | miR-5195-3p/GRB10 | Potential diagnostic marker for myeloid leukemia. Induces proliferation. Inhibits apoptosis and regulates cell cycle. | [60] | |
| Circ_0004136 | KCNQ5 | Cytoplasm and shuttled by exosome | Up in serum of pediatric AML patients, HL60 and K-562 | miR-570-3p/TSPAN3 | Induces cell viability, migration and invasion. Inhibits apoptosis and regulates cell cycle. | [61] |
| CircSPI1 | SPI1 | Cytoplasm | Up in AML patients, THP1 and NB4 | miR-1307-3p, miR-382-5P, miR-767-5p | Induces proliferation. Inhibits myeloid differentiation and apoptosis. | [62] |
| Circ_0005774 | Up in blood of pediatric AML, HL-60 and NB4 | miR-192-5p/ULK1 | Induces proliferation. Inhibits apoptosis and regulates cell cycle. | [63] | ||
| Circ_0003420 (Hsa_circ_0003420) | Down in non-m3 AML stem (BM) cells | 3’UTR of IGF2BP1 | Prognostic marker. Inhibits leukemogenesis. Induces apoptosis. | [64] | ||
| CircTASP1 (Hsa_circ_0007340) | TASP1 | Cytoplasm | Up in peripheral blood, HL60, KG-1, U937 and THP-1 | miR-515-5p/HMGA2 | Induces proliferation. Inhibits apoptosis. | [65] |
| Hsa_circ_0079480 | Up in serum and most profound in M4/M5 type | Prognostic marker. | [66] | |||
| Circ-ANAPC7 | ANAPC7 | Up in BM | Diagnosed and relapsed AML. | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, S.-C.; Cheong, F.J.F.; Tay, Y. Circular RNAs and Untranslated Regions in Acute Myeloid Leukemia. Int. J. Mol. Sci. 2023, 24, 3215. https://doi.org/10.3390/ijms24043215
Yeh S-C, Cheong FJF, Tay Y. Circular RNAs and Untranslated Regions in Acute Myeloid Leukemia. International Journal of Molecular Sciences. 2023; 24(4):3215. https://doi.org/10.3390/ijms24043215
Chicago/Turabian StyleYeh, Shih-Chia, Faith J. F. Cheong, and Yvonne Tay. 2023. "Circular RNAs and Untranslated Regions in Acute Myeloid Leukemia" International Journal of Molecular Sciences 24, no. 4: 3215. https://doi.org/10.3390/ijms24043215
APA StyleYeh, S.-C., Cheong, F. J. F., & Tay, Y. (2023). Circular RNAs and Untranslated Regions in Acute Myeloid Leukemia. International Journal of Molecular Sciences, 24(4), 3215. https://doi.org/10.3390/ijms24043215