1. Introduction
Extracellular chromatin, such as neutrophil extracellular traps (NETs), although beneficial in fighting bacteria [
1,
2], can also have detrimental effects such as prolonging tissue repair, promoting unwanted coagulation, and serving as a nidus for autoantigen generation [
2,
3,
4]. Tools and methods to conveniently evaluate the ability of bodily fluids to degrade extracellular chromatin, therefore, are sought after for their potential to understand and discern disease states [
5,
6,
7]. In systemic lupus erythematosus (SLE), for example, decreased NET degradation by serum may indicate imminent flares, including serious manifestations such as lupus nephritis [
8]. With an eye towards bioengineering useful tools for the evaluation of extracellular chromatin degradation by bodily fluids and in vitro cell cultures, this study describes a time-lapse imaging and image processing method for use in the analysis of the nuclease- and serum-mediated degradation of a NET-mimetic material that our lab recently developed with a defined composition and mesoscale structures.
DNA–histone mesostructures (DHMs) [
9] and microwebs [
10,
11] resemble aspects of the chromatin backbone structure of NETs. For example, we previously reported that deoxyribonuclease I (DNase I) degrades both DHMs [
9] and microwebs [
10] in a manner that resembles the nuclease-mediated degradation that NETs undergo. Here, we analyzed DHM degradation in more detail including a comparison to NETs. The types of nucleases tested were also expanded to include micrococcal nuclease (MNase), which has a different specificity from DNase I. MNase is known to preferentially cut the linker DNA between nucleosomes well before it cleaves DNA that is wrapped around octamers [
12,
13]. DNase cleaves DNA less selectively with regards to DNA strands wrapped around histones. We also performed an exploratory analysis of DHM degradation by human serum. A revised method of DHM preparation that provides structures that are more consistent and wash resistant was also developed for more reliable degradation analyses. Finally, the degradation process was analyzed in more detail by visualizing DHMs with time-lapse imaging and automated analysis of the many images captured. DHMs spotted into 96-well microplates have a defined initial mesoscale (sub-millimeter scale) structure that facilitates image-based degradation monitoring (
Supplementary Figure S1) in ways not possible with the numerous smaller structures presented by NETs. The time-lapse imaging of DHM degradation by serum revealed a process involving a combination of structural dissolution and detachment not observed in DNase-I-mediated degradation. Interestingly, DHMs, unlike NETs, were not degraded well by MNase. Compared to NETs, we also found that DHMs degraded slower when exposed to serum. Despite these differences, DHMs could distinguish differences in the ability of normal control vs. severely-ill SLE patient serum in order to degrade them.
3. Discussion
Our group previously described a minimal NET-mimetic microweb, comprised of DNA and histones, for their ability to mimic aspects of the bacteria-suppressive features of NETs [
10,
11]. We have also described a surface-attached NET-mimetic biomaterial we call DHMs, which have a defined mesoscale structure also comprised of DNA and histones, for their NET-mimetic, immune-cell-activating features [
9]. The microbicidal and cell-activating functions of these bioengineered NET-mimetic biomaterials, however, may be affected by their degradation rate. Therefore, in this study, we characterized the nuclease- and serum-mediated degradation of DHMs, with additional comparisons to NETs. Furthermore, given the reported potential diagnostic applications of NET degradation [
5,
6,
7], we also evaluated whether DHM degradation rates differ between treatment with normal versus SLE serum.
The degradation assay was designed to quantify the kinetics and visualize the degradation, including detachment and associated shape changes, of DHM structures in real time. To select a time frame of observation, we considered the treatment incubation time of NET degradation assay protocols, which range widely from 90 min to 24 h [
17]. Physiologically, it is unknown how long NETs persist in circulation or in tissues, with reports ranging from hours [
18] to weeks [
19]. We optimized the time frame to be one automated image capture per hour for at least 24 h, which allowed observations of the gradual degradation responses that DHMs exhibit, and the full extent of degradation for a given treatment once it reached a plateau.
Several fluorescence-based NET degradation assays rely on end-point measurements taken before and after the degradation incubation in a buffer or PBS [
7,
20]. It is challenging to report live fluorescence kinetics because serum introduces a great deal of signal to the wells, which interferes with the pre-stained NET signal. Since our study involved the real-time monitoring of wells that contain serum, we developed an image segmentation method to enable the automated real-time quantification of DHM area change to overcome the background serum samples and confounding fluorescence signals from degraded DHM fragments in suspension. The defined starting shape of the DHMs coupled with the image segmentation method allowed the visualization of different degradation patterns from different treatments, as well as the quantitative tracking of DHM area. Area metrics, when paired with DHM raw intensity values, could also be useful in capturing the brightness of the structures during instances of delamination, where folding of the structure could lead to over- and underestimation of the area. There is an opportunity to improve the signal-to-noise ratio of DHMs in future work, for simplified real-time fluorescence tracking that does not rely on image analysis.
DNase I treatment led to gradual DHM fading consistent with the gradual digestion of the DNA backbone into smaller nucleotide fragments (
Figure 3). Buffer controls caused minimal DHM shape change. Treatment with 5% normal serum led to more detachment in the early phases accompanied by a slower digestion process, leading to the formation of gathered aggregates during the intermediate stages of the degradation process (
Figure 4). The differences in how DHM degradation proceeded between DNase I and serum are consistent with reports that there are mechanisms beyond DNase I that contribute to NET degradation in vivo [
21]. We note that the use of an in-incubator microscope has previously been used in the real-time imaging of NETosis [
22,
23] but not NET degradation. Our DHM degradation analysis method expands the use of such instruments for NET-degradation-related studies.
Compared to NETs, DHMs were degraded more slowly by diluted serum (
Figure 5). Additionally, we observed minor variability between serum aliquots of the same origin, as observed between the DHM degradation of 5% serum in
Figure 4A, which was slower to degrade than 5% serum in
Figure 5A. To dissect the differences between the cell-based NETs and DHMs in the presence of nucleases, we tested both MNase and DNase. DNase I and MNase are two of the most commonly reported nucleases in NET degradation assays [
17]. DNase I is active in human blood [
16,
24], while MNase is produced by bacteria; however, MNase is of interest because it is known to preferentially cut the linker DNA between nucleosomes well before it cuts DNA that is wrapped around octamers [
12,
13]. DNase cleaves DNA less selectively with regards to DNA strands wrapped around histones. DHMs and NETs were similarly fully degraded by 10 U/mL DNase. In particular, we were surprised that DHMs were not susceptible to degradation by 10 U/mL MNase, whereas NETs were readily degraded under the same conditions. During NETosis, histone citrullination mediates chromatin decondensation in neutrophils and granulocytes via peptidyl arginine deiminase 4 (PAD4) [
25]. PAD4 facilitates NETosis, and it has been reported that MNase more readily digests externalized NETs after PAD4 treatment [
25]. We speculate that these and other differences may result in DHM chromatin fibers being less accessible compared to those found in NETs. Other nucleases of interest to compare in future applications are the leukocyte-secreted nuclease DNase IL3 [
26], and nucleases secreted by common Gram-positive bacteria to digest NETs as an evasion mechanism [
27].
We note that we did not normalize the enzyme:DNA ratio between DHMs and NETs, which could impact the length of oligonucleotide byproducts and therefore the degradation accessibility. The coverage of DHMs was larger, and centered, versus NETs which were distributed across the well but individually in the micrometer size range. Despite the increased aggregation of DHMs, by our estimates, cell-derived NETs were composed of 6.7 times more DNA than DHMs per well. The DHMs were fabricated with a mass of 30 ng DNA per well, while the cellular NETs in this study had approximately 180 ng to 240 ng per well (equivalent to 30,000–40,000 NETotic cells per well of 100,000 plated neutrophils stimulated with a 20 nM phorbol 12-myristate 13-acetate (PMA) concentration). Despite the lower DNA amount per well, the DHMs still were slower to degrade using the same 10 U/mL MNase treatment, hence our conclusion that there are likely also structural differences that are behind the lower DHM degradation by MNase.
Impaired NET clearance contributes to the onset and progression of autoimmunity such as in SLE [
2]. NETs become a source of self-antigens that can generate autoantibodies against components of NETs, such as anti-nuclear antibodies (ANAs). ANAs may lead to SLE flares by forming immune complexes that deposit on, and damage, skin and kidney tissues [
4,
8]. NET degradation assays have been reported for their potential to discern between disease states and predict SLE flares [
28]. Here, we tested the potential of serum-mediated DHM degradation to also relate to SLE disease states. DHMs were degraded less by serum from a patient with higher disease activity (higher SLEDAI score) (
Figure 6), consistent with the lowered chromatin and NET degradation observed in active SLE [
4,
20]. These results suggest promise for the further development of DHM degradation assays of clinical samples. The convenient-to-use protocol and stable long-term storage of DHMs, which is not possible with NETs, may facilitate translational research. On the other hand, our analysis showed that DHMs were more resistant to nuclease- and serum-mediated degradation compared to NETs, suggesting that modifications may be needed in how DHMs are prepared to better mimic the degradation profiles of NETs. Future development can also improve the well-to-well homogeneity of the DHM structures upon drying, which could introduce differences in DNA density. There is inherent biological variability in the nuclease and NET degradation activity among serum donors; therefore, these improvements can enable the application of the DHM degradation assay to a larger population of autoimmune patients in order to address this variability.
4. Materials and Methods
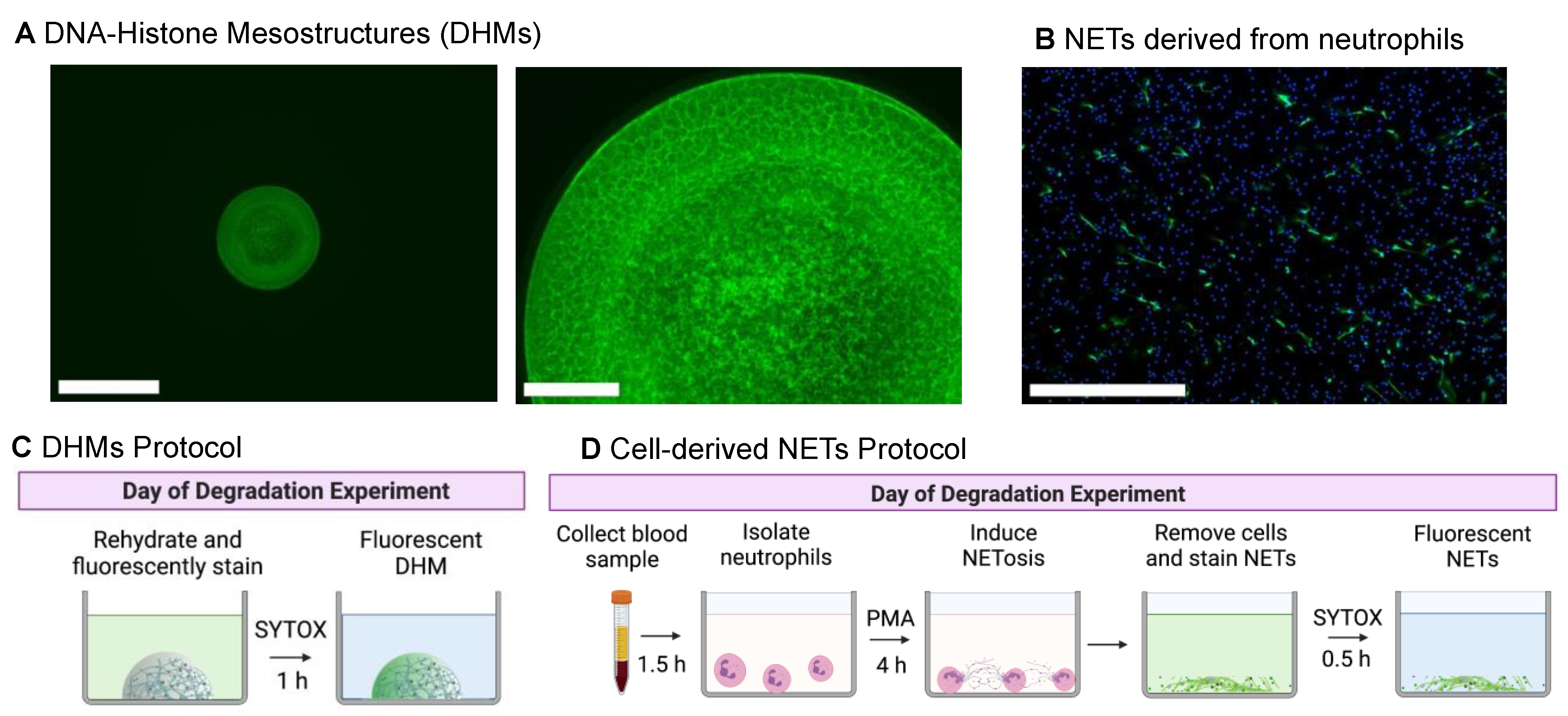
4.1. DNA–Histone Mesostructure (DHM) Fabrication
DHMs were fabricated by the step-wise spotting of DNA and histone droplets into wells of standard 96-well microplates. To increase the binding efficiency of DHMs, untreated 96-well microplates (CLS3370; Corning, Corning, NY, USA) were coated with 0.001% poly-L-lysine (P4707; Sigma-Aldrich, St. Louis, MO, USA) for 10 min and rinsed once with water. Once dry, the DNA layer was prepared by combining a solution of 0.1 mg/mL lambda-phage methylated DNA (D9768; Sigma-Aldrich, St. Louis, MO, USA) with a 400 mM trehalose solution (T-104-4; Pfanstiehl, Waukegan, IL, USA) at a 1:1 ratio. Suspending the soluble DNA within a trehalose phase added stability, localization, and uniformity to the final structures. The trehalose phase could be washed off prior to testing, leaving only the chromatin fibers in each well. As a note, histones could not be pre-mixed with the DNA solution as they instantly formed fibers and therefore could not be dispensed with liquid-handling techniques.
A sub-microliter droplet, 0.6 μL, of the DNA–trehalose solution was dispensed into each well. The droplets were vitrified for 24 h in a vacuum desiccator. A 0.6 μL droplet containing a 0.5 mg/mL histone solution (H9250; Sigma-Aldrich, St. Louis, MO, USA) was then deposited over the vitrified DNA spot and dried for another 24 h in a vacuum desiccator. The concentration of DNA and histone solutions, as well as the volume, can be modified to vary the resulting morphology and compaction of the DHM chromatin structure. The binding of histones to DNA strands resulted in a defined sub-millimeter mesh of condensed chromatin fibers within the boundaries of the initial droplet area. Once fabricated, the resulting vitrified fibrous DNA structures were stable at room temperature until use.
4.2. Degradation Assay Using DHMs
Rehydration and fluorescent staining of DHMs. Each DHM structure was rehydrated from its dry state, and simultaneously, the DNA component was fluorescently stained by adding 100 μL of a 1 μM SYTOX Green (S7020; Thermo Fisher Scientific, Waltham, MA, USA) in PBS for one hour, protected from light. The DHMs were washed three times in PBS to remove the trehalose phase and rinsed once with a 1% bovine serum albumin (BSA) solution (126609; EMD Millipore, Burlington, MA, USA) in PBS to further stabilize the structures.
Sample preparation. DHMs were treated with DNase I from bovine pancreas (D4263; Sigma-Aldrich, St. Louis, MO, USA), MNase (88216; Sigma-Aldrich, St. Louis, MO, USA), or thawed normal control serum (H4522, or S1-M; reagents from Sigma-Aldrich, St. Louis, MO, USA) at the specified concentrations above. DNase I units (U) represented Kunitz units per manufacturer information. Serum was commercially available from pooled donor samples collected from the clots of healthy normal humans. With regards to H4522 male AB blood, per the manufacturer, male blood is chosen because female serum can have more antibodies resulting from pregnancy. Type AB serum is used as anti-A and/or anti-B antibodies are absent, which are found in other blood types. Serum samples were aliquoted upon receipt and stored at −80 °C until use. All degradation treatment samples were diluted in nuclease buffer (10 mM Tris–HCl pH 7.5, 10 mM MgCl2, 2 mM CaCl2, and 150 mM NaCl). Nuclease buffer solution served as the background control.
Real-time imaging procedure. To visualize DHM degradation, each well was gently aspirated, and 100 μL of the sample treatment was added to each DHM structure. The degradation was monitored continuously for 24 to 48 h at 37 °C and 5% CO2, using an Incucyte S3® Live-Cell Analysis System (Essen BioScience, Ann Arbor, MI, USA). Each sample was tested at least in triplicate. After 5 min in the incubator, the lid and the bottom of the plate were wiped with a kimwipe to remove any condensation. Automated images were captured every hour using a 4X objective with brightfield and green (400-ms) channels. This enabled the real-time visualization of degradation for each DHM structure. After the degradation period, the residual DHMs were imaged using a 20X objective in brightfield and green fluorescence channels.
Degradation analysis. The instrument’s image analysis software, Incucyte® Software 2022A (Essen BioScience, Ann Arbor, MI, USA) was used to quantify the kinetics and the extent of DHM degradation throughout the assay. The Incucyte® Software Basic Analyzer algorithm is an image processing tool applied to define and segment a DHM object area for each image. First, images were selected to train the image segmentation algorithm to identify which objects to be classified as DHMs. The image thresholding and segmentation parameters were set to a radius of 100 μm, and a threshold value of 2 GCU, with Top-Hat background correction. The software generated previews of the DHM object mask using the selected object definition parameters to confirm that the green object mask captured the DHMs. Once the image segmentation definition was finalized, the algorithm processed all the captured images to generate a DHM mask area, observed as a magenta overlay in each image and calculated as μm2 per image by the Incucyte® Software 2022A (Essen BioScience, Ann Arbor, MI, USA). The DHM mask area values were exported for analysis using GraphPad Prism 9 software (GraphPad Software, San Diego, CA, USA).
4.3. NET Degradation Assay
PMA-stimulated NETs were degraded as previously described with minor modifications [
7]. Briefly, microplates were coated with 0.001% poly-L-lysine (P4707; Sigma-Aldrich, St. Louis, MO, USA) for 10 min and rinsed once with water and allowed to air dry. Purified control neutrophils were resuspended in RPMI media supplemented with L-glutamine (Gibco; Thermo Fisher Scientific, Waltham, MA, USA). Next, 1 × 10
5 neutrophils were seeded into each well 96-well black-wall clear-bottom tissue culture plate (Thermo Fisher Scientific, Waltham, MA, USA). To induce NET formation, cells were incubated in the presence of 20 nM PMA (Sigma-Aldrich, St. Louis, MO, USA) for 4 h at 37 °C and 5% CO
2. Following incubation, the culture media was gently aspirated and washed once with 1× PBS. NETosis was quantified by incubating the cells for 30 min at 37 °C, and 5% CO
2 with SYTOX Green (S7020; Thermo Fisher Scientific, Waltham, MA, USA) diluted in PBS to a final concentration of 1 µM. Culture supernatant was gently aspirated, and 1× PBS was added carefully to each well. Fluorescence was quantified at excitation and emission wavelengths of 504 nm and 523 nm using a Cytation 5 Cell Imaging Multi-Mode Reader (BioTek, Santa Clara, CA, USA). To assess NET degradation, PBS was gently aspirated from each well, and NETs were incubated for 90 min (at 37 °C and 5% CO
2) with normal control serum samples diluted to 5% in nuclease buffer. Each sample was tested in triplicate, with MNase- (10 U/mL) and DNase- (10 U/mL) treated wells serving as positive controls. Following incubation, the supernatant was gently discarded, and wells were washed once with 1× PBS. Fresh PBS was added to each well, and residual NETs were quantified by re-measuring SYTOX fluorescence at excitation and emission wavelengths of 504 nm and 523 nm using a Cytation 5 Cell Imaging Multi-Mode Reader (BioTek, Santa Clara, CA, USA).
4.4. Immunofluorescence Microscopy of NETs
First, 1 × 105 normal control neutrophils in serum-free RPMI media supplemented with L-glutamine were seeded onto 0.001% poly-L-lysine-coated coverslips. NET formation was induced by incubating neutrophils with 20 nM PMA for 4 h at 37 °C and 5% CO2. Following stimulation, culture media was gently aspirated, and cells were washed with 1× phosphate-buffered saline (PBS). To visualize the effects of NET degradation, each treatment was incubated with the cells for 90 min at 37 °C and 5% CO2. The treatments for visualization were 5% healthy control serum, MNase (10 U/mL) and DNase (10 U/mL) diluted in nuclease buffer (10 mM Tris–HCl pH 7.5, 10 mM MgCl2, 2 mM CaCl2, and 50 mM NaCl), or nuclease buffer as a control. Following incubation, culture media was gently aspirated and washed once with 1× PBS. Cells were fixed with 4% paraformaldehyde (PFA) at room temperature for 10 min, followed by overnight blocking in blocking buffer (10% fetal bovine serum (FBS) in PBS). For protein staining, fixed cells were incubated with polyclonal antibody to neutrophil elastase (Sigma-Aldrich, St. Louis, MO, USA) in a blocking buffer for 1 h at 4 °C, followed by FITC-conjugated secondary (Southern Biotech, Birmingham, AL, USA) for 1 h at 4 °C. Nuclear DNA was detected with Hoechst 33342 (H3570; Thermo Fisher Scientific, Waltham, MA, USA). Coverslips were mounted with Prolong Gold Antifade (Thermo Fisher Scientific, Waltham, MA, USA), and images were collected with a Cytation 5 Cell Imaging Multi-Mode Reader (BioTek, Santa Clara, CA, USA).
4.5. Immunofluorescence Microscopy of DHM Degradation by SLE Serum
To assess the degradation of DHMs coated on a 96-well microplate, 5% normal control or lupus serum diluted in nuclease buffer was added to the DHM-coated wells. For the SLE serum samples, the low-disease-activity serum was from a patient with SLEDAI-2K = 2. The high-disease-activity serum was from a patient with SLEDAI-2K = 16. The clinical activity level of SLE patients was calculated using SLEDAI-2K according to Gladman et al. [
29]. DHMs were incubated for 12 h at 37 °C and 5% CO
2. Following incubation, the culture supernatant was gently aspirated, and wells were washed once with 1× PBS. Degradation was assessed by incubating the wells with SYTOX Green diluted in PBS to a final concentration of 1 µM for 10 min at 37 °C and 5% CO
2. Following incubation, the culture supernatant was gently aspirated, and wells were washed once with 1× PBS. Fresh PBS was added to each well, and images were captured by Cytation 5 Cell Imaging Multi-Mode Reader (BioTek, Santa Clara, CA, USA).
4.6. Statistical Analysis
The data were analyzed using GraphPad Prism 9.4.1 (458) (GraphPad Software, San Diego, CA, USA) and Excel 16.65 (Microsoft). Results are presented as mean ± SEM. Multiple groups were compared with the ANOVA test or its modification with post-hoc tests. Two groups were compared using the paired t-test. The differences between groups were reported as the following statistical values: * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}