
Glycyl-tRNA Synthetase (GARS) Expression Is Associated with Prostate Cancer Progression and Its Inhibition Decreases Migration, and Invasion In Vitro

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Moderate and High GARS Expression in PCa Patients
2.2. GARS Expression in Relation to Gleason Grade Grouping (GG)
2.3. GARS in Relation to Overall Survival (OS) and Cause-Specific Survival (CSS)
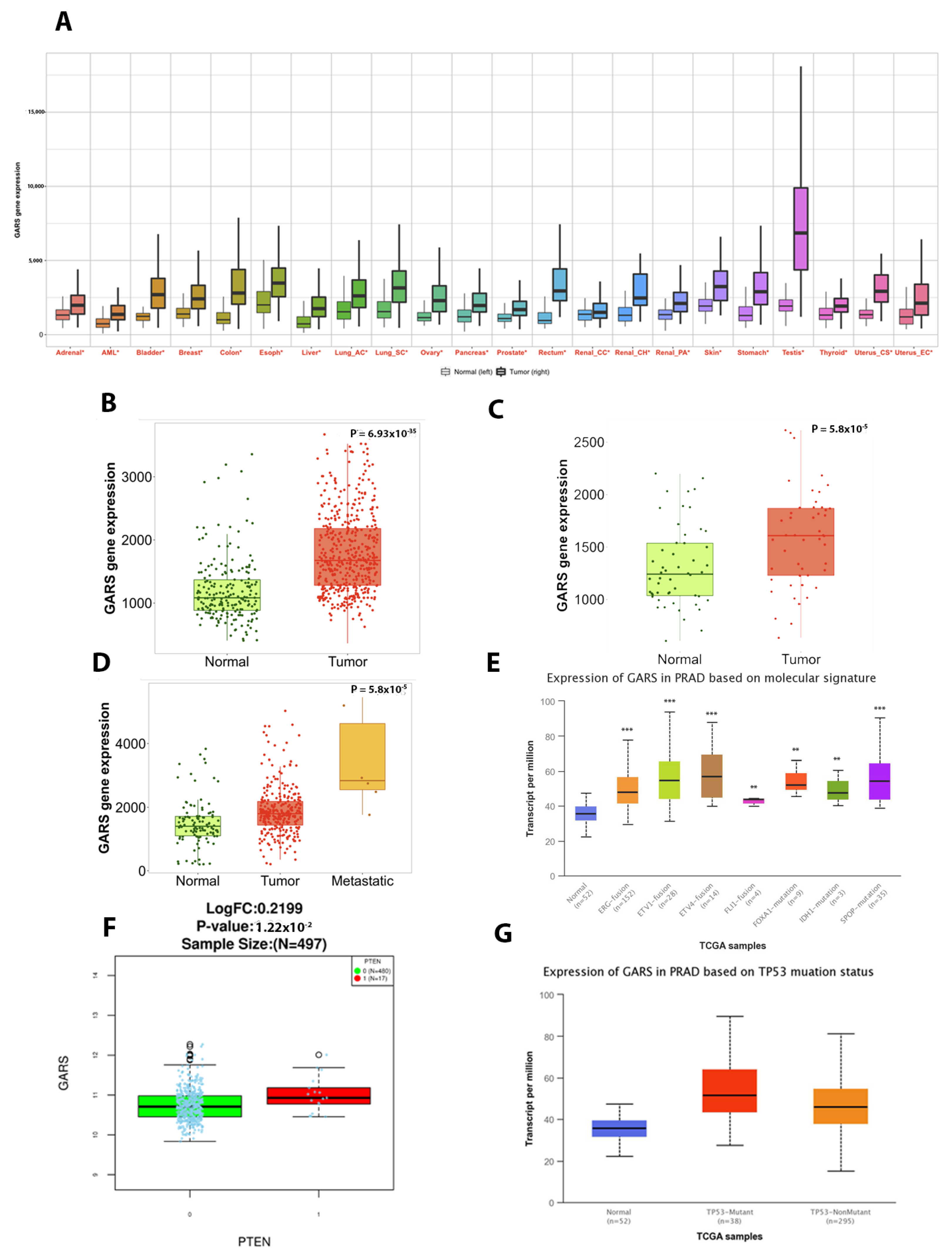
2.4. GARS mRNA Is Upregualted in Most Cancer Types and Significantly Associated with Poor Prognostic Biomarkers
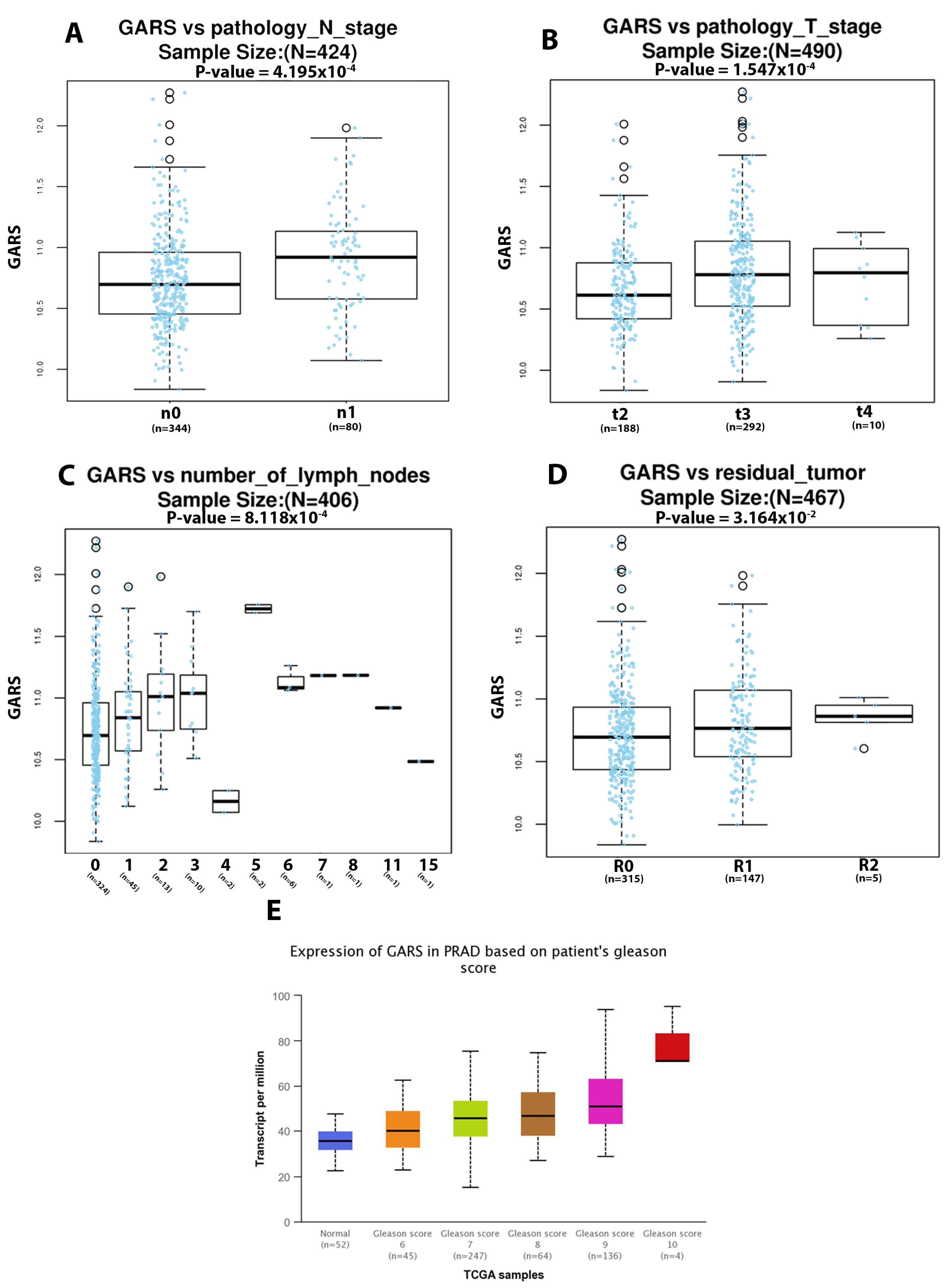
2.5. GARS Expression Is Associated to Lymph Node and Pathological Staging in PCa Based on TCGA PRAD Database
2.6. GARS Overexpression Depicted Potential Biological and Molecualar Functions Associated with Poor Clinical Outcomes in TCGA PRAD
2.7. GARS Overrepresentation Enrichment Analysis (ORA) in PCa TCGA PRAD Database
2.8. GARS Expression in PCa Cell Lines and GARS Knockdown
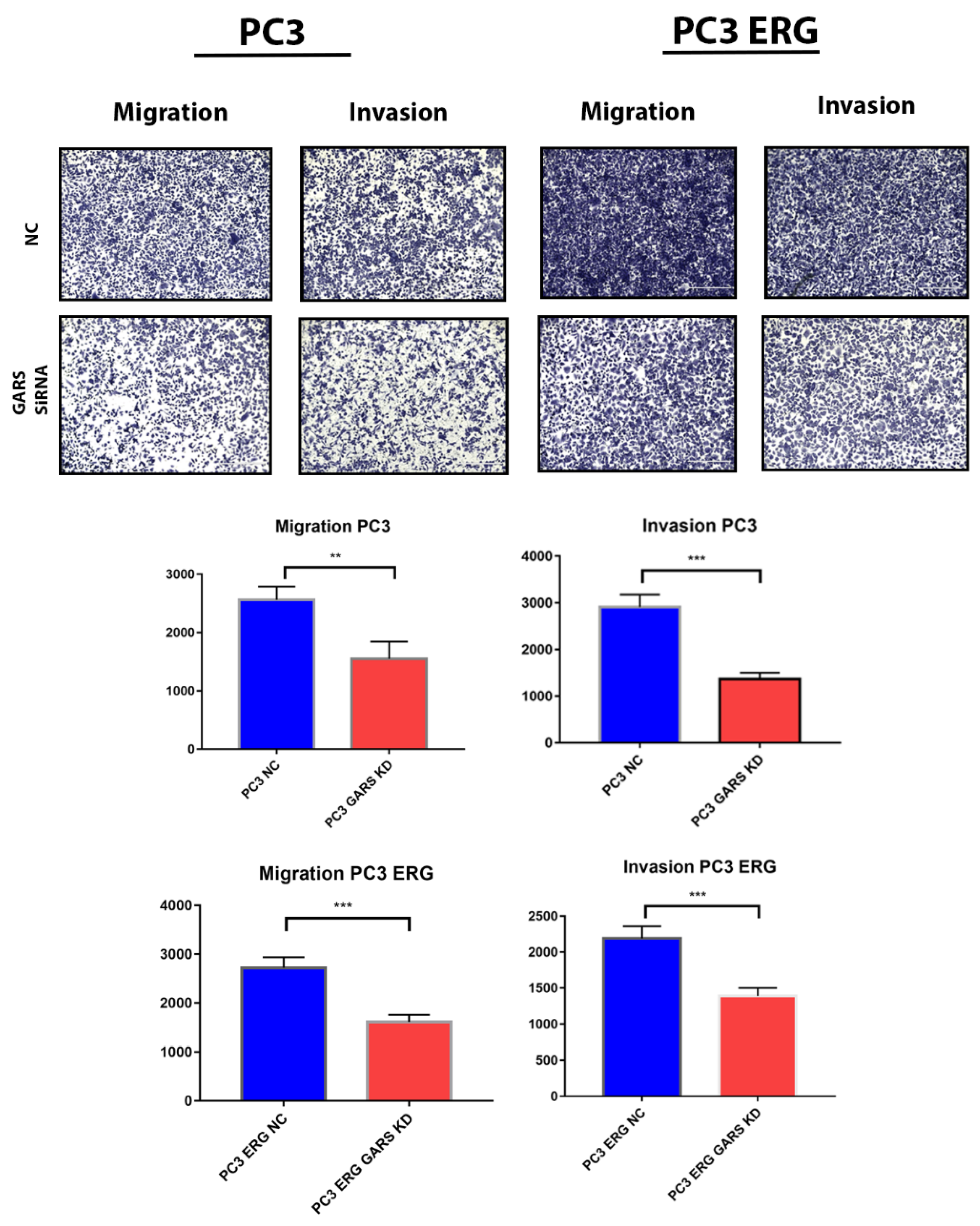
2.9. GARS Knockdown Attenuated PCa Cell Migration and Invasion
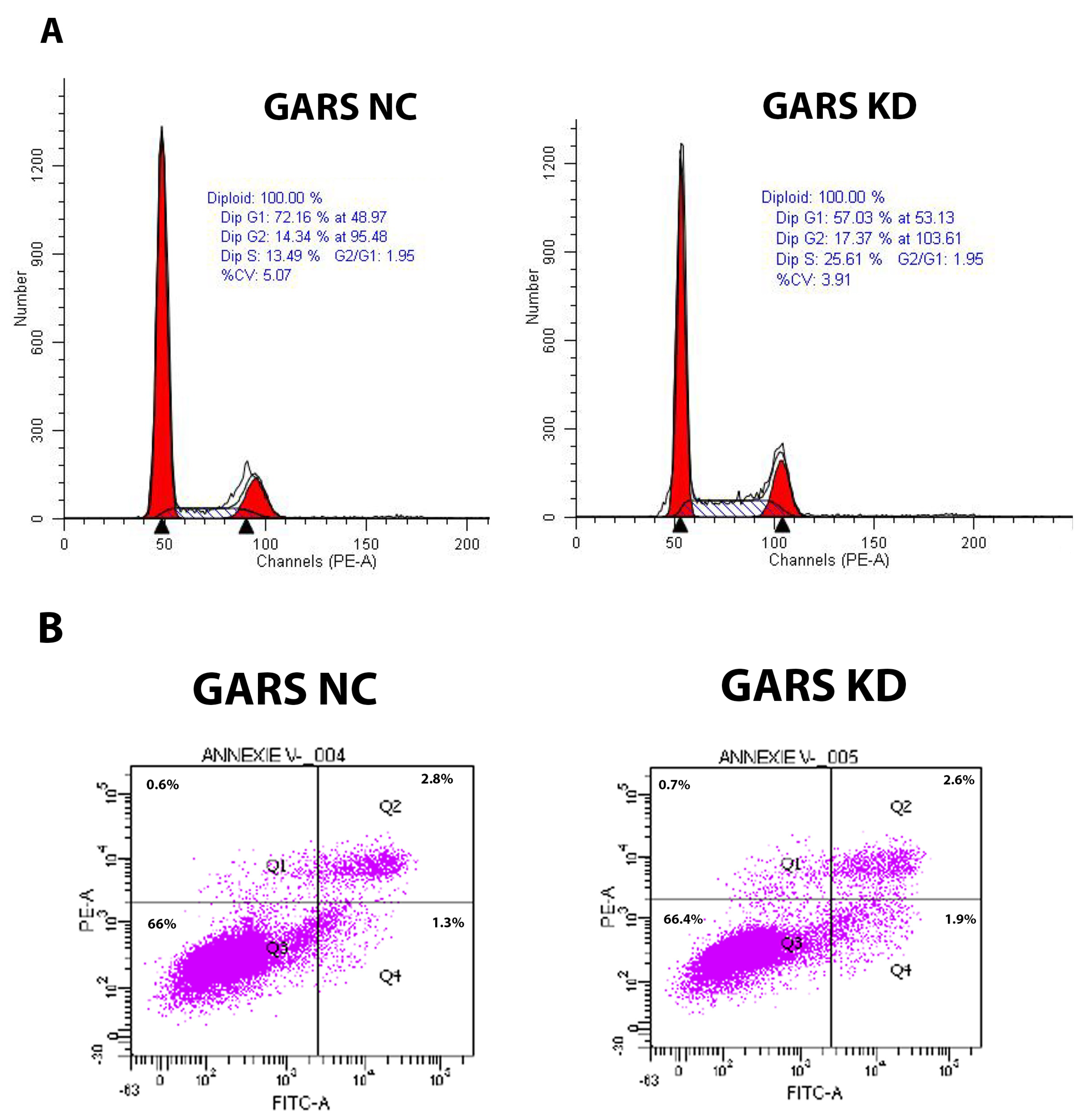
2.10. GARS Is a Cell Cycle Regulator in PCa
3. Discussion
4. Materials and Methods
4.1. Study Population, Tissue Microarray Construction, and Pathological Analysis
4.2. Immunohistochemistry (IHC)
4.3. Cell Lines
4.4. Cell Line Transfection via RNA Silencing
4.5. Western Blot
4.6. Migration and Invasion Assay
4.7. Flow Cytometry
4.8. GARS Expression in the Cancer Genome Atlas Prostate Adenocarcinoma (TCGA PRAD)
4.9. GARS Expression in TCGA PRAD Analyzed through UALCAN
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GARS | Glycyl-tRNA synthetase |
| PCa | Prostate Cancer |
| OS | Overall Survival |
| CSS | Cause Specific Survival |
| HCC | Hepatocellular Carcinoma |
| ERG | ETS-related gene |
| PTEN | Phosphatase tensin homologue |
| ETV1 | ETS variant transcription factor 1 |
| ETV4 | ETS variant transcription factor 4 |
| FOXA1 | Forkhead box protein A1 |
| IDH1 | Isocitrate dehydrogenase 1 |
| SPOP | Speckle type BTB/POZ protein |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Fujita, K.; Nonomura, N. Urinary biomarkers of prostate cancer. Int. J. Urol. 2018, 25, 770–779. [Google Scholar] [CrossRef] [Green Version]
- Wise, H.M.; Hermida, M.A.; Leslie, N.R. Prostate cancer, pi3k, pten and prognosis. Clin. Sci. 2017, 131, 197–210. [Google Scholar] [CrossRef]
- Kumari, S.; Sharma, V.; Tiwari, R.; Maurya, J.P.; Subudhi, B.B.; Senapati, D. Therapeutic potential of p53 reactivation in prostate cancer: Strategies and opportunities. Eur. J. Pharmacol. 2022, 919, 174807. [Google Scholar] [CrossRef]
- Wang, J.; Yang, B.; Wang, D.; Han, R.; Bi, Z.; Lin, L. Gars is implicated in poor survival and immune infiltration of hepatocellular carcinoma. Cell. Signal. 2022, 94, 110302. [Google Scholar] [CrossRef]
- Malissovas, N.; Griffin, L.B.; Antonellis, A.; Beis, D. Dimerization is required for gars-mediated neurotoxicity in dominant cmt disease. Hum. Mol. Genet. 2016, 25, 1528–1542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Dong, W.; Zhang, J.; Liu, W.; Yin, J.; Shi, D.; Ma, W. A novel mitochondrial-related nuclear gene signature predicts overall survival of lung adenocarcinoma patients. Front. Cell Dev. Biol. 2021, 9, 740487. [Google Scholar] [CrossRef]
- Chen, C.J.; Chou, C.Y.; Shu, K.H.; Chen, H.C.; Wang, M.C.; Chang, C.C.; Hsu, B.G.; Wu, M.S.; Yang, Y.L.; Liao, W.L.; et al. Discovery of novel protein biomarkers in urine for diagnosis of urothelial cancer using itraq proteomics. J. Proteome Res. 2021, 20, 2953–2963. [Google Scholar] [CrossRef]
- Li, X.; Sun, H.; Liu, Q.; Liu, Y.; Hou, Y.; Jin, W. Conjoint analysis of circulating tumor cells and solid tumors for exploring potential prognostic markers and constructing a robust novel predictive signature for breast cancer. Cancer Cell Int. 2021, 21, 708. [Google Scholar] [CrossRef]
- Luo, R.; Zheng, C.; Song, W.; Tan, Q.; Shi, Y.; Han, X. High-throughput and multi-phases identification of autoantibodies in diagnosing early-stage breast cancer and subtypes. Cancer Sci. 2022, 113, 770–783. [Google Scholar] [CrossRef]
- Vellaichamy, A.; Sreekumar, A.; Strahler, J.R.; Rajendiran, T.; Yu, J.; Varambally, S.; Li, Y.; Omenn, G.S.; Chinnaiyan, A.M.; Nesvizhskii, A.I. Proteomic interrogation of androgen action in prostate cancer cells reveals roles of aminoacyl trna synthetases. PLoS ONE 2009, 4, e7075. [Google Scholar] [CrossRef] [Green Version]
- Ritch, C.; Cookson, M. Recent trends in the management of advanced prostate cancer. F1000Research 2018, 7, 1513. [Google Scholar] [CrossRef] [Green Version]
- Udager, A.M.; Tomlins, S.A. Molecular biomarkers in the clinical management of prostate cancer. Cold Spring Harb. Perspect Med. 2018, 8, a030601. [Google Scholar] [CrossRef]
- Shi, Q.; Zhu, Y.; Ma, J.; Chang, K.; Ding, D.; Bai, Y.; Gao, K.; Zhang, P.; Mo, R.; Feng, K.; et al. Prostate cancer-associated spop mutations enhance cancer cell survival and docetaxel resistance by upregulating caprin1-dependent stress granule assembly. Mol. Cancer 2019, 18, 170. [Google Scholar] [CrossRef]
- Qian, C.; Li, D.; Chen, Y. Ets factors in prostate cancer. Cancer Lett. 2022, 530, 181–189. [Google Scholar] [CrossRef]
- Martin, N.E.; Mucci, L.A.; Loda, M.; Depinho, R.A. Prognostic determinants in prostate cancer. Cancer J. 2011, 17, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Sehn, J.K. Prostate cancer pathology: Recent updates and controversies. Mo. Med. 2018, 115, 151–155. [Google Scholar]
- Daneshmand, S.; Quek, M.L.; Stein, J.P.; Lieskovsky, G.; Cai, J.; Pinski, J.; Skinner, E.C.; Skinner, D.G. Prognosis of patients with lymph node positive prostate cancer following radical prostatectomy: Long-term results. J. Urol. 2004, 172, 2252–2255. [Google Scholar] [CrossRef]
- Murray, N.P. Minimal residual disease in prostate cancer patients after primary treatment: Theoretical considerations, evidence and possible use in clinical management. Biol. Res. 2018, 51, 32. [Google Scholar] [CrossRef]
- McCabe, N.; Kennedy, R.D.; Prise, K.M. The role of pten as a cancer biomarker. Oncoscience 2016, 3, 54–55. [Google Scholar] [CrossRef]
- Arora, K.; Barbieri, C.E. Molecular subtypes of prostate cancer. Curr. Oncol. Rep. 2018, 20, 58. [Google Scholar] [CrossRef]
- De Laere, B.; Oeyen, S.; Mayrhofer, M.; Whitington, T.; van Dam, P.J.; Van Oyen, P.; Ghysel, C.; Ampe, J.; Ost, P.; Demey, W.; et al. Tp53 outperforms other androgen receptor biomarkers to predict abiraterone or enzalutamide outcome in metastatic castration-resistant prostate cancer. Clin. Cancer Res. 2019, 25, 1766–1773. [Google Scholar] [CrossRef] [Green Version]
- Kivinummi, K.; Urbanucci, A.; Leinonen, K.; Tammela, T.L.J.; Annala, M.; Isaacs, W.B.; Bova, G.S.; Nykter, M.; Visakorpi, T. The expression of aurka is androgen regulated in castration-resistant prostate cancer. Sci. Rep. 2017, 7, 17978. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Yin, H. Pdk1 promotes tumor cell proliferation and migration by enhancing the warburg effect in non-small cell lung cancer. Oncol. Rep. 2017, 37, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; He, C.; Zhu, B.; Jiang, J.; Chen, Y.; Ma, T. 3-phosphoinositide dependent protein kinase-1 (pdk-1) promotes migration and invasion in gastric cancer cells through activating the nf-kappab pathway. Oncol. Res. 2017, 25, 1153–1159. [Google Scholar] [CrossRef]
- Li, W.; Qian, L.; Lin, J.; Huang, G.; Hao, N.; Wei, X.; Wang, W.; Liang, J. Cd44 regulates prostate cancer proliferation, invasion and migration via pdk1 and pfkfb4. Oncotarget 2017, 8, 65143–65151. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Brenner, J.C.; Sabolch, A.; Jackson, W.; Speers, C.; Wilder-Romans, K.; Knudsen, K.E.; Lawrence, T.S.; Chinnaiyan, A.M.; Feng, F.Y. Targeted radiosensitization of ets fusion-positive prostate cancer through parp1 inhibition. Neoplasia 2013, 15, 1207–1217. [Google Scholar] [CrossRef]
- Bartha, A.; Gyorffy, B. Tnmplot.Com: A web tool for the comparison of gene expression in normal, tumor and metastatic tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Hermanek, P.; Wittekind, C. Residual tumor (r) classification and prognosis. Semin. Surg. Oncol. 1994, 10, 12–20. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. Linkedomics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. Webgestalt 2019: Gene set analysis toolkit with revamped uis and apis. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. Ualcan: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. Ualcan: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GARS | Gleason Grade Group 1 | Gleason Grade Group 2 | Gleason Grade Group 3 | Gleason Grade Group 4 | Gleason Grade Group 5 |
|---|---|---|---|---|---|
| Score 0, 1, 2 | 54 (91.5%) | 9 (60.0%) | 15 (78.9%) | 9 (81.8%) | 55 (71.4%) |
| Score 3 | 5 (8.5%) | 6 (40.0%) | 4 (21.1%) | 2 (18.2%) | 22 (28.6%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khosh Kish, E.; Gamallat, Y.; Choudhry, M.; Ghosh, S.; Seyedi, S.; Bismar, T.A. Glycyl-tRNA Synthetase (GARS) Expression Is Associated with Prostate Cancer Progression and Its Inhibition Decreases Migration, and Invasion In Vitro. Int. J. Mol. Sci. 2023, 24, 4260. https://doi.org/10.3390/ijms24054260
Khosh Kish E, Gamallat Y, Choudhry M, Ghosh S, Seyedi S, Bismar TA. Glycyl-tRNA Synthetase (GARS) Expression Is Associated with Prostate Cancer Progression and Its Inhibition Decreases Migration, and Invasion In Vitro. International Journal of Molecular Sciences. 2023; 24(5):4260. https://doi.org/10.3390/ijms24054260
Chicago/Turabian StyleKhosh Kish, Ealia, Yaser Gamallat, Muhammad Choudhry, Sunita Ghosh, Sima Seyedi, and Tarek A. Bismar. 2023. "Glycyl-tRNA Synthetase (GARS) Expression Is Associated with Prostate Cancer Progression and Its Inhibition Decreases Migration, and Invasion In Vitro" International Journal of Molecular Sciences 24, no. 5: 4260. https://doi.org/10.3390/ijms24054260