
Functional Alteration and Differential Expression of the Bitter Taste Receptor T2R38 in Human Paranasal Sinus in Patients with Chronic Rhinosinusitis
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Background and Characteristics of Subjects in the Study
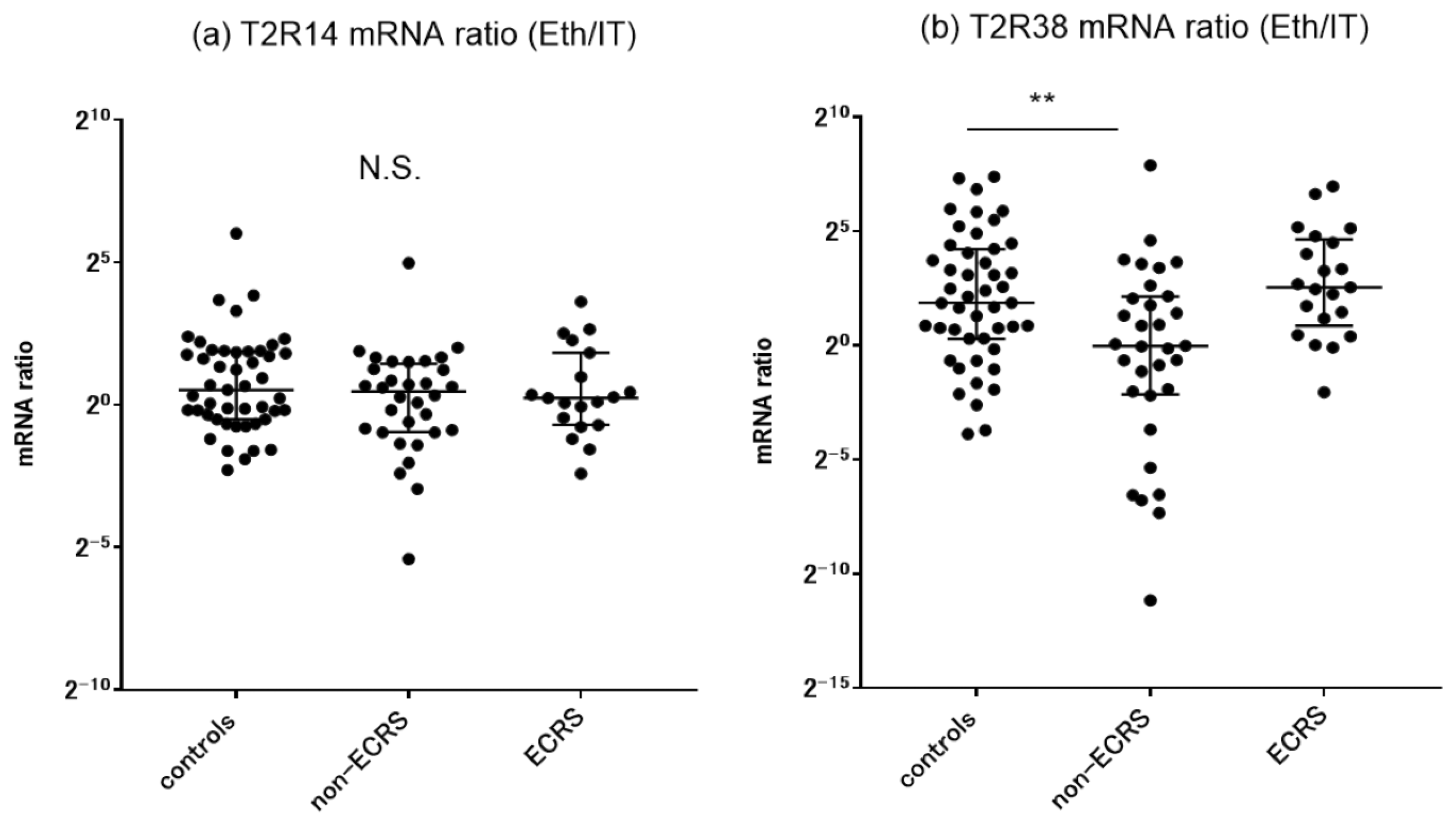
2.2. Target Genes Expression in Sinonasal Mucosa and Nasal Polyps
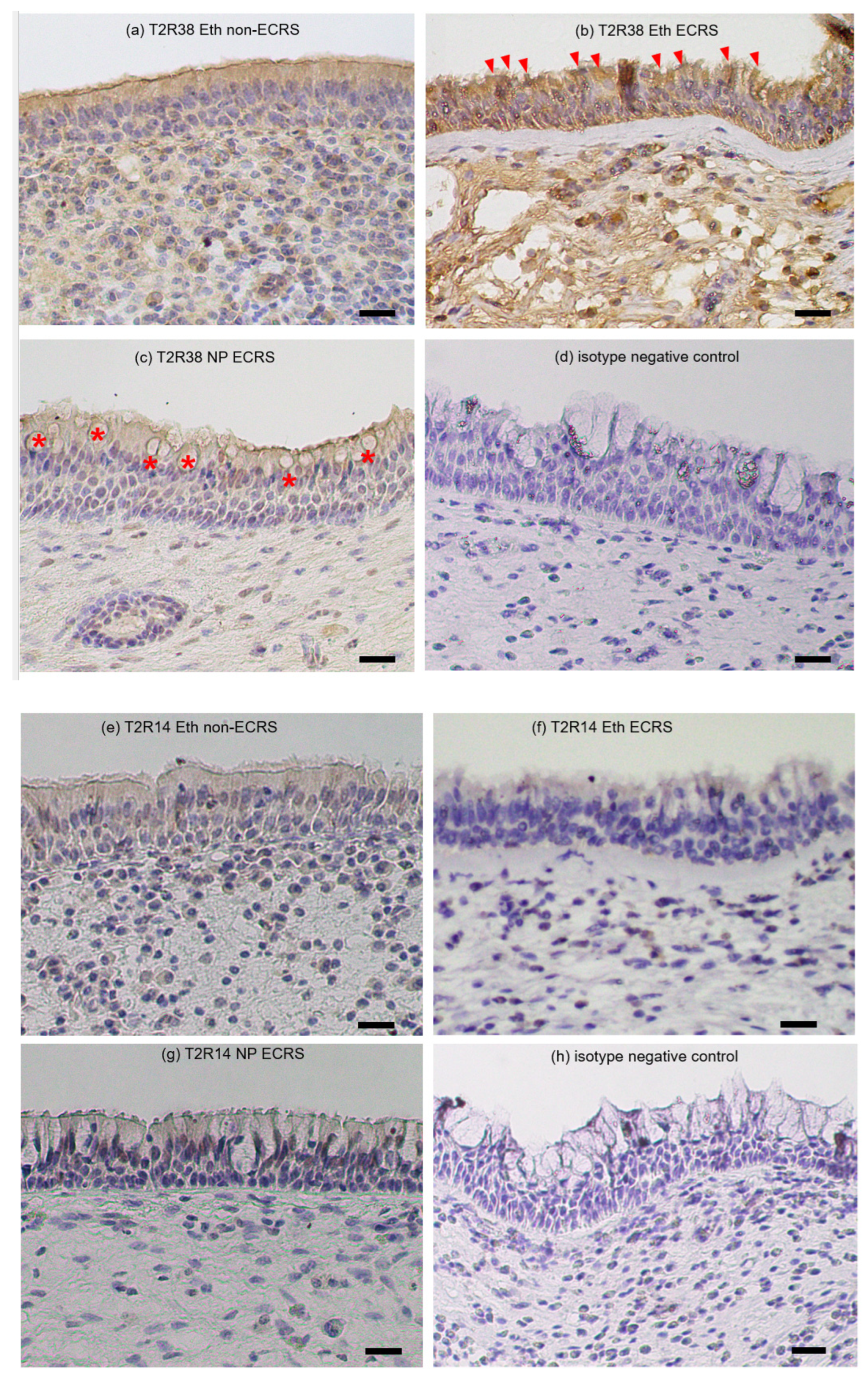
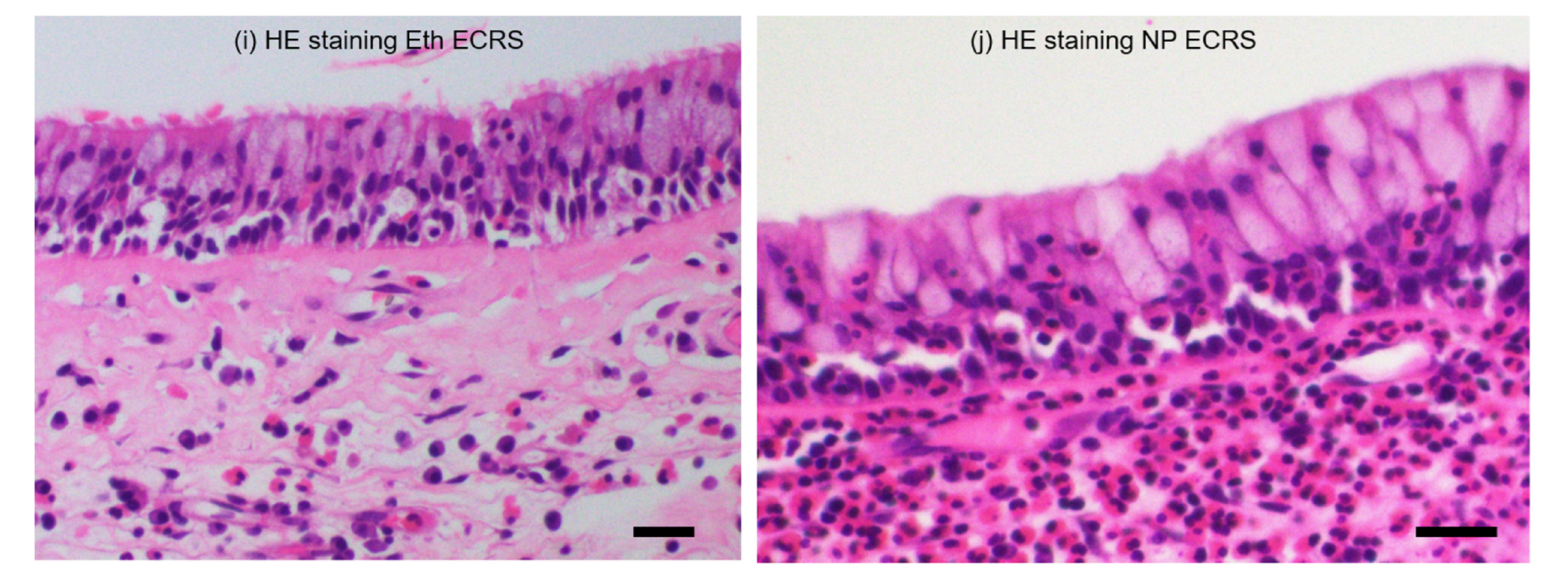
2.3. Immunohistochemical Distribution of T2R14 and T2R38 Proteins
2.4. FeNO Levels in CRS Patients
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. RT-PCR Analysis
4.3. Immunohistochemistry
4.4. Genotyping the SNP Polymorphism in the TAS2R38 Gene
4.5. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindemann, B. Chemoreception: Tasting the sweet and the bitter. Curr. Biol. 1996, 6, 1234–1237. [Google Scholar] [CrossRef] [Green Version]
- Dagan-Wiener, A.; Di Pizio, A.; Nissim, I.; Bahia, M.S.; Dubovski, N.; Margulis, E.; Niv, M.Y. BitterDB: Taste ligands and receptors database in 2019. Nucleic Acids Res. 2019, 47, D1179–D1185. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef]
- Jaggupilli, A.; Howard, R.; Aluko, R.E.; Chelikani, P. Advanced Glycation End-Products Can Activate or Block Bitter Taste Receptors. Nutrients 2019, 11, 1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avau, B.; Depoortere, I. The bitter truth about bitter taste receptors: Beyond sensing bitter in the oral cavity. Acta. Physiol. 2016, 216, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Shaik, F.A.; Singh, N.; Arakawa, M.; Duan, K.; Bhullar, R.P.; Chelikani, P. Bitter taste receptors: Extraoral roles in pathophysiology. Int. J. Biochem. Cell Biol. 2016, 77, 197–204. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, C.H.; Lifshitz, L.M.; ZhuGe, R. Extraoral bitter taste receptors in health and disease. J. Gen. Physiol. 2017, 149, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggupilli, A.; Singh, N.; Jesus, V.C.; Duan, K.; Chelikani, P. Characterization of the Binding Sites for Bacterial Acyl Homoserine Lactones (AHLs) on Human Bitter Taste Receptors (T2Rs). ACS Infect. Dis. 2018, 4, 1146–1156. [Google Scholar] [CrossRef]
- Workman, A.D.; Patel, N.N.; Carey, R.M.; Kuan, E.C.; Cohen, N.A. The Role of Taste Receptors in Airway Innate Immune Defense. Sinusitis 2018, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Kuek, L.E.; Lee, R.J. First contact: The role of respiratory cilia in host-pathogen interactions in the airways. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L603–L619. [Google Scholar] [CrossRef]
- Mfuna-Endam, L.; Filali-Mouhim, A.; Boisvert, P.; Boulet, L.P.; Bossé, Y.; Desrosiers, M. Genetic variations in taste receptors are associated with chronic rhinosinusitis: A replication study. Int. Forum Allergy Rhinol. 2014, 4, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Adappa, N.D.; Zhang, Z.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Lysenko, A.; Reed, D.R.; Scott, T.; Zhao, N.W.; Owens, D.; et al. The bitter taste receptor T2R38 is an independent risk factor for chronic rhinosinusitis requiring sinus surgery. Int. Forum Allergy Rhinol. 2014, 4, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adappa, N.D.; Truesdale, C.M.; Workman, A.D.; Doghramji, L.; Mansfield, C.; Kennedy, D.W.; Palmer, J.N.; Cowart, B.J.; Cohen, N.A. Correlation of T2R38 taste phenotype and in vitro biofilm formation from nonpolypoid chronic rhinosinusitis patients. Int. Forum Allergy Rhinol. 2016, 6, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Rom, D.I.; Christensen, J.M.; Alvarado, R.; Sacks, R.; Harvey, R.J. The impact of bitter taste receptor genetics on culturable bacteria in chronic rhinosinusitis. Rhinology 2017, 55, 90–94. [Google Scholar] [CrossRef]
- Kawasumi, T.; Takeno, S.; Ishikawa, C.; Takahara, D.; Taruya, T.; Takemoto, K.; Hamamoto, T.; Ishino, T.; Ueda, T. The Functional Diversity of Nitric Oxide Synthase Isoforms in Human Nose and Paranasal Sinuses: Contrasting Pathophysiological Aspects in Nasal Allergy and Chronic Rhinosinusitis. Int. J. Mol. Sci. 2021, 22, 7561. [Google Scholar] [CrossRef]
- Chen, J.; Larson, E.D.; Anderson, C.B.; Agarwal, P.; Frank, D.N.; Kinnamon, S.C.; Ramakrishnan, V.R. Expression of Bitter Taste Receptors and Solitary Chemosensory Cell Markers in the Human Sinonasal Cavity. Chem. Senses 2019, 44, 483–495. [Google Scholar] [CrossRef]
- Hariri, B.M.; McMahon, D.B.; Chen, B.; Freund, J.R.; Mansfield, C.J.; Doghramji, L.J.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; Reed, D.R.; et al. Flavones modulate respiratory epithelial innate immunity: Anti-inflammatory effects and activation of the T2R14 receptor. J. Biol. Chem. 2017, 292, 8484–8497. [Google Scholar] [CrossRef] [Green Version]
- Levit, A.; Nowak, S.; Peters, M.; Wiener, A.; Meyerhof, W.; Behrens, M.; Niv, M.Y. The bitter pill: Clinical drugs that activate the human bitter taste receptor TAS2R14. FASEB J. 2014, 28, 1181–1197. [Google Scholar] [CrossRef]
- Kuek, L.E.; McMahon, D.B.; Ma, R.Z.; Miller, Z.A.; Jolivert, J.F.; Adappa, N.D.; Palmer, J.N.; Lee, R.J. Cilia Stimulatory and Antibacterial Activities of T2R Bitter Taste Receptor Agonist Diphenhydramine: Insights into Repurposing Bitter Drugs for Nasal Infections. Pharmaceuticals 2022, 15, 452. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef]
- Tan, B.K.; Klingler, A.I.; Poposki, J.A.; Stevens, W.W.; Peters, A.T.; Suh, L.A.; Norton, J.; Carter, R.G.; Hulse, K.E.; Harris, K.E.; et al. Heterogeneous inflammatory patterns in chronic rhinosinusitis without nasal polyps in Chicago, Illinois. J. Allergy Clin. Immunol. 2017, 139, 699–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokunaga, T.; Sakashita, M.; Haruna, T.; Asaka, D.; Takeno, S.; Ikeda, H.; Nakayama, T.; Seki, N.; Ito, S.; Murata, J.; et al. Novel scoring system and algorithm for classifying chronic rhinosinusitis: The JESREC Study. Allergy 2015, 70, 995–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujieda, S.; Imoto, Y.; Kato, Y.; Ninomiya, T.; Tokunaga, T.; Tsutsumiuchi, T.; Yoshida, K.; Kidoguchi, M.; Takabayashi, T. Eosinophilic chronic rhinosinusitis. Allergol. Int. 2019, 68, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Vandeplas, G.; Huynh, T.M.T.; Joish, V.N.; Mannent, L.; Tomassen, P.; Van Zele, T.; Cardell, L.O.; Arebro, J.; Olze, H.; et al. The Global Allergy and Asthma European Network (GALEN rhinosinusitis cohort: A large European cross-sectional study of chronic rhinosinusitis patients with and without nasal polyps. Rhinology 2019, 57, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Civantos, A.M.; Arnold, M.; Stevens, E.M.; Cowart, B.J.; Colquitt, L.R.; Mansfield, C.; Kennedy, D.W.; Brooks, S.G.; Workman, A.D.; et al. Divergent bitter and sweet taste perception intensity in chronic rhinosinusitis patients. Int. Forum Allergy Rhinol. 2021, 11, 857–865. [Google Scholar] [CrossRef]
- Lin, Y.T.; Yeh, T.H. Studies on Clinical Features, Mechanisms, and Management of Olfactory Dysfunction Secondary to Chronic Rhinosinusitis. Front. Allergy 2022, 3, 835151. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, J.; Jang, S.S.; Wrobel, B.B.; Ference, E.H. Smell and Taste Impairment in a Nationwide Sample of US Adults with Chronic Rhinosinusitis Symptoms. OTO Open 2021, 5, 2473974X20986756. [Google Scholar] [CrossRef] [PubMed]
- Civantos, A.M.; Maina, I.W.; Arnold, M.; Lin, C.; Stevens, E.M.; Tan, L.H.; Gleeson, P.K.; Colquitt, L.R.; Cowart, B.J.; Bosso, J.V.; et al. Denatonium benzoate bitter taste perception in chronic rhinosinusitis subgroups. Int. Forum Allergy Rhinol. 2021, 11, 967–975. [Google Scholar] [CrossRef]
- Mennella, J.A.; Pepino, M.Y.; Reed, D.R. Genetic and environmental determinants of bitter perception and sweet preferences. Pediatrics 2005, 115, e216–e222. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.E.; Wallace, M.R.; Knopik, V.S.; Herbstman, D.M.; Bartoshuk, L.M.; Duffy, V.B. Allelic variation in TAS2R bitter receptor genes associates with variation in sensations from and ingestive behaviors toward common bitter beverages in adults. Chem. Senses 2011, 36, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.H.; Hahn, S.; McMahon, D.; Bonislawski, D.; Kennedy, D.W.; Adappa, N.D.; Palmer, J.N.; Jiang, P.; Lee, R.J.; Cohen, N.A. Nitric oxide production is stimulated by bitter taste receptors ubiquitously expressed in the sinonasal cavity. Am. J. Rhinol. Allergy 2017, 31, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavinia, M.; Keshavarzian, A.; Tobin, M.C.; Landay, A.L.; Schleimer, R.P. A Comprehensive Review of the Nasal Microbiome in Chronic Rhinosinusitis (CRS). Clin. Exp. Allergy 2016, 46, 21–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, S.H.; Bachert, C.; Lockey, R.F. Chronic Rhinosinusitis Phenotypes: An Approach to Better Medical Care for Chronic Rhinosinusitis. J. Allergy Clin. Immunol. Pract. 2016, 4, 639–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomassen, P.; Vandeplas, G.; Van Zele, T.; Cardell, L.O.; Arebro, J.; Olze, H.; Förster-Ruhrmann, U.; Kowalski, M.L.; Olszewska-Ziąber, A.; Holtappels, G.; et al. Inflammatory endotypes of chronic rhinosinusitis based on cluster analysis of biomarkers. J. Allergy Clin. Immunol. 2016, 137, 1449–1456.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, A.; Peters, A.T.; Stevens, W.W.; Schleimer, R.P.; Tan, B.K.; Kern, R.C. Endotypes of chronic rhinosinusitis: Relationships to disease phenotypes, pathogenesis, clinical findings, and treatment approaches. Allergy 2022, 77, 812–826. [Google Scholar] [CrossRef]
- Brzost, J.; Czerwaty, K.; Dżaman, K.; Ludwig, N.; Piszczatowska, K.; Szczepański, M.J. Perspectives in Therapy of Chronic Rhinosinusitis. Diagnostics 2022, 12, 2301. [Google Scholar] [CrossRef]
- Malik, B.; Elkaddi, N.; Turkistani, J.; Spielman, A.I.; Ozdener, M.H. Mammalian Taste Cells Express Functional Olfactory Receptors. Chem. Senses 2019, 44, 289–301. [Google Scholar] [CrossRef]
- Barham, H.P.; Cooper, S.E.; Anderson, C.B.; Tizzano, M.; Kingdom, T.T.; Finger, T.E.; Kinnamon, S.C.; Ramakrishnan, V.R. Solitary chemosensory cells and bitter taste receptor signaling in human sinonasal mucosa. Int. Forum Allergy Rhinol. 2013, 3, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Piatt, G.; Ambrosetti, U.; Aldè, M.; Girotto, G.; Concas, M.P.; Torretta, S. Chronic Rhinosinusitis: T2r38 Genotyping and Nasal Cytology in Primary Ciliary Dyskinesia. Laryngoscope 2023, 133, 248–254. [Google Scholar] [CrossRef]
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzaki, H.; Fujieda, S.; Masuyama, K. Japanese Guidelines for Allergic Rhinitis 2017. Allergol. Int. 2017, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Stout, S.L.; Wyatt, T.A.; Adams, J.J.; Sisson, J.H. Nitric Oxide-Dependent Cilia Regulatory Enzyme Localization in Bovine Bronchial Epithelial Cells. J. Histochem. Cytochem. 2007, 55, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile cilia of human airway epithelia are chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef] [Green Version]
- Zborowska-Piskadło, K.; Stachowiak, M.; Rusetska, N.; Sarnowska, E.; Siedlecki, J.; Dżaman, K. The expression of bitter taste receptor TAS2R38 in patients with chronic rhinosinusitis. Arch. Immunol. Ther. Exp. 2020, 68, 26. [Google Scholar] [CrossRef]
- Cohen, N.A. The Genetics of the Bitter Taste Receptor T2R38 in Upper Airway Innate Immunity and Implications for Chronic Rhinosinusitis. Laryngoscope 2017, 127, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Workman, A.D.; Carey, R.M.; Kohanski, M.A.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Cohen, N.A. Relative susceptibility of airway organisms to antimicrobial effects of nitric oxide. Int. Forum Allergy Rhinol. 2017, 7, 770–776. [Google Scholar] [CrossRef]
- American Thoracic Society; European Respiratory Society. ATS/ERS recommendations for standardized procedures for the online and offline measurement of exhaled lower respiratory nitric oxide and nasal nitric oxide, 2005. Am. J. Respir. Crit. Care Med. 2005, 171, 912–930. [Google Scholar] [CrossRef] [Green Version]
- Takeno, S.; Taruya, T.; Ueda, T.; Noda, N.; Hirakawa, K. Increased Exhaled Nitric Oxide and Its Oxidation Metabolism in Eosinophilic Chronic Rhinosinusitis. Auris Nasus Larynx 2013, 40, 458–464. [Google Scholar] [CrossRef]
- Kurokawa, R.; Kanemitsu, Y.; Fukumitsu, K.; Takeda, N.; Yap, J.M.; Ozawa, Y.; Masaki, A.; Ono, J.; Izuhara, K.; Nishiyama, H.; et al. Nasal Polyp Eosinophilia and FeNO May Predict Asthma Symptoms Development after Endoscopic Sinus Surgery in CRS Patients without Asthma. J. Asthma 2022, 59, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Frendø, M.; Hakansson, K.; Schwer, S.; Ravn, A.; Meteran, H.; Porsbjerg, C.; Backer, V.; von Buchwald, C. Exhaled and Nasal Nitric Oxide in Chronic Rhinosinusitis Patients with Nasal Polyps in Primary Care. Rhinology 2018, 56, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Triantafillou, V.; Workman, A.D.; Patel, N.N.; Maina, I.W.; Tong, C.C.L.; Kuan, E.C.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Waizel-Haiat, S.; et al. Broncho-Vaxom® (OM-85 BV) soluble components stimulate sinonasal innate immunity. Int. Forum Allergy Rhinol. 2019, 9, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Adappa, N.D.; Palmer, J.N.; Lee, R.J. Neuropeptide Y Reduces Nasal Epithelial T2R Bitter Taste Receptor-Stimulated Nitric Oxide Production. Nutrients 2021, 13, 3392. [Google Scholar] [CrossRef] [PubMed]
- Biarnés, X.; Marchiori, A.; Giorgetti, A.; Lanzara, C.; Gasparini, P.; Carloni, P.; Born, S.; Brockhoff, A.; Behrens, M.; Meyerhof, W. Insights into the binding of Phenyltiocarbamide (PTC) agonist to its target human TAS2R38 bitter receptor. PLoS ONE 2010, 5, e12394. [Google Scholar] [CrossRef] [PubMed]
- Lipchock, S.V.; Mennella, J.A.; Spielman, A.I.; Reed, D.R. Human bitter perception correlates with bitter receptor messenger RNA expression in taste cells. Am. J. Clin. Nutr. 2013, 98, 1136–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adappa, N.D.; Farquhar, D.; Palmer, J.N.; Kennedy, D.W.; Doghramji, L.; Morris, S.A.; Owens, D.; Mansfield, C.; Lysenko, A.; Lee, R.J.; et al. TAS2R38 genotype predicts surgical outcome in nonpolypoid chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 6, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Tizzano, M.; Gulbransen, B.D.; Vandenbeuch, A.; Clapp, T.R.; Herman, J.P.; Sibhatu, H.M.; Churchill, M.E.; Silver, W.L.; Kinnamon, S.C.; Finger, T.E. Nasal chemosensory cells use bitter taste signaling to detect irritants and bacterial signals. Proc. Natl. Acad. Sci. USA 2010, 107, 3210–3215. [Google Scholar] [CrossRef] [Green Version]
- Takemoto, K.; Takeno, S.; Ishino, T.; Ueda, T.; Hamamoto, T.; Horibe, Y.; Takahara, D.; Kawasumi, T. Nitric oxide synthase-2 (CCTTT)n polymorphism is associated with local gene expression and clinical manifestations in patients with chronic rhinosinusitis. Eur. J. Inflamm. 2022, 20, 20587392211052948. [Google Scholar] [CrossRef]
- Lund, V.J.; Kennedy, D.W. Quantification for staging sinusitis. The Staging and Therapy Group. Ann. Otol. Rhinol. Laryngol. Suppl. 1995, 167, 17–21. [Google Scholar] [CrossRef]
- Takeno, S.; Noda, N.; Hirakawa, K. Measurements of nasal fractional exhaled nitric oxide with a hand-held device in patients with allergic rhinitis: Relation to cedar pollen dispersion and laser surgery. Allergol. Int. 2012, 61, 93–100. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls | Non-ECRS | ECRS | |
|---|---|---|---|
| Number (male/female) | 51 (27/24) | 56 (35/21) | 36 (18/18) |
| Age (mean ± SD) | 44.7 ± 17 | 56.1 ± 15.7 ††† | 54 ± 11.3 †† |
| Allergic rhinitis (%) | 35 (68.6%) | 31 (55.4%) | 28 (77.8%) |
| BMI (kg/mm2) (mean ± SD) | 23.8 ± 3.7 | 23.2 ± 3.3 | 22.6 ± 3.5 |
| Bronchial asthma (%) | 6 (11.8%) | 6 (10.7%) | 19 (52.8%) *** |
| Blood eosinophils (%) (median, range) | 3.6 (1.6–5.7) | 1.9 (1.5–3.5) † | 6.95 (5.5–9.6) *** |
| Tissue eosinophils (cells/HPF) (median, range) | 4.6 (0.7–14) | 5 (2.4–18.2) | 117 (75–233) *** |
| CT score (mean ± SD) | 7.27 ± 5.8 | 15 ± 5.2 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takemoto, K.; Lomude, L.S.; Takeno, S.; Kawasumi, T.; Okamoto, Y.; Hamamoto, T.; Ishino, T.; Ando, Y.; Ishikawa, C.; Ueda, T. Functional Alteration and Differential Expression of the Bitter Taste Receptor T2R38 in Human Paranasal Sinus in Patients with Chronic Rhinosinusitis. Int. J. Mol. Sci. 2023, 24, 4499. https://doi.org/10.3390/ijms24054499
Takemoto K, Lomude LS, Takeno S, Kawasumi T, Okamoto Y, Hamamoto T, Ishino T, Ando Y, Ishikawa C, Ueda T. Functional Alteration and Differential Expression of the Bitter Taste Receptor T2R38 in Human Paranasal Sinus in Patients with Chronic Rhinosinusitis. International Journal of Molecular Sciences. 2023; 24(5):4499. https://doi.org/10.3390/ijms24054499
Chicago/Turabian StyleTakemoto, Kota, Luga Santo Lomude, Sachio Takeno, Tomohiro Kawasumi, Yukako Okamoto, Takao Hamamoto, Takashi Ishino, Yuki Ando, Chie Ishikawa, and Tsutomu Ueda. 2023. "Functional Alteration and Differential Expression of the Bitter Taste Receptor T2R38 in Human Paranasal Sinus in Patients with Chronic Rhinosinusitis" International Journal of Molecular Sciences 24, no. 5: 4499. https://doi.org/10.3390/ijms24054499