

Effects of Different Gene Editing Modes of CRISPR/Cas9 on Soybean Fatty Acid Anabolic Metabolism Based on GmFAD2 Family
 and
and
Abstract
:1. Introduction
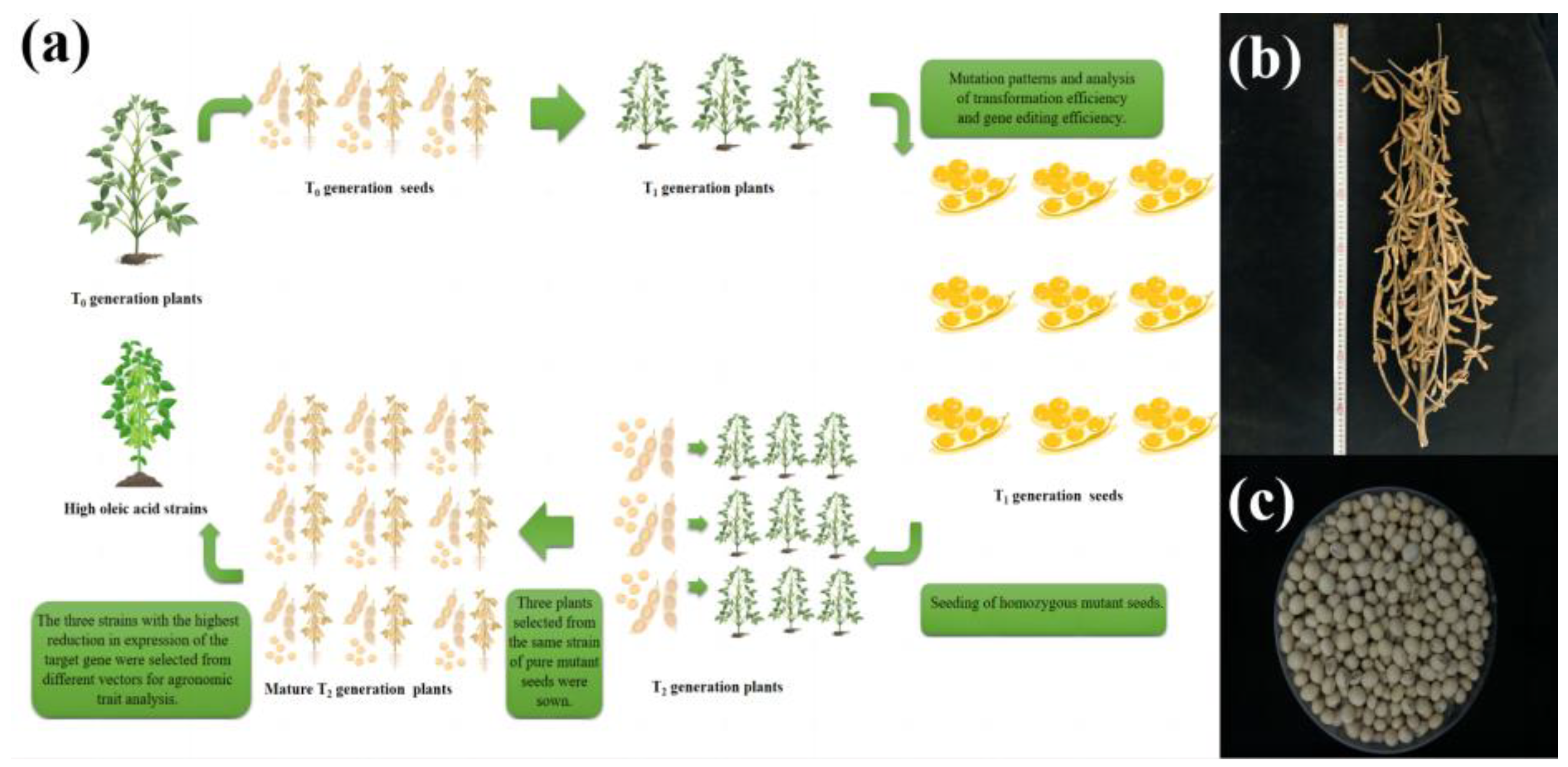
2. Results
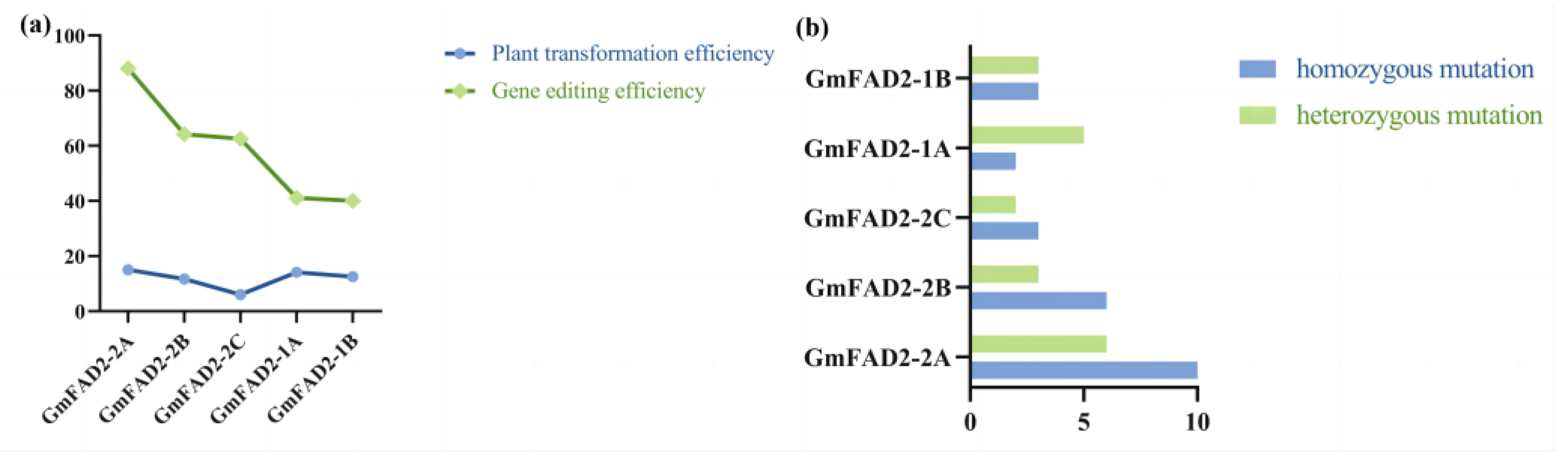
2.1. Construction of Gene-Editing Vectors
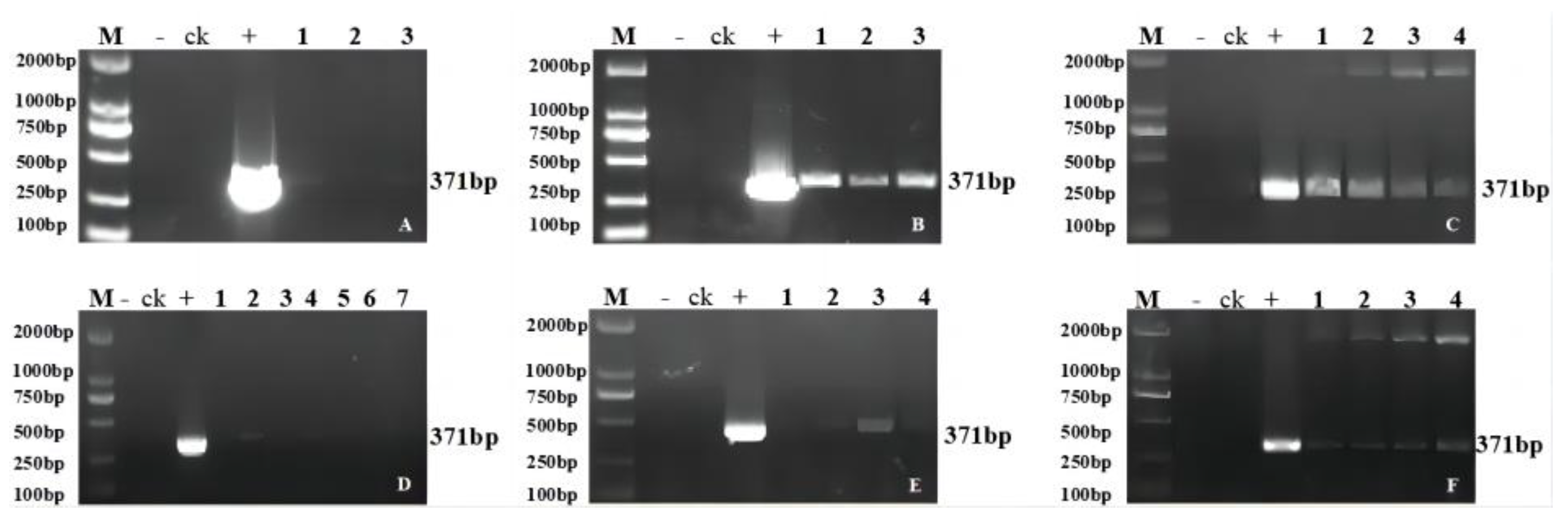
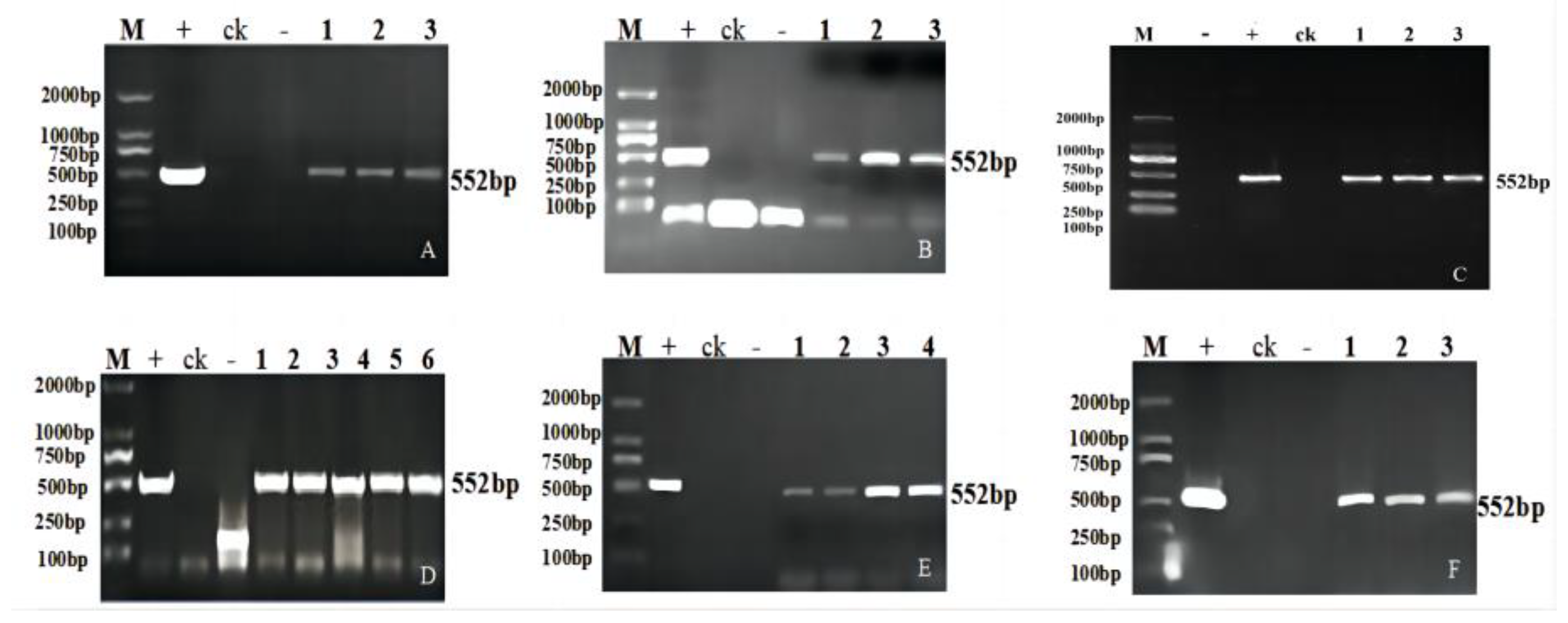
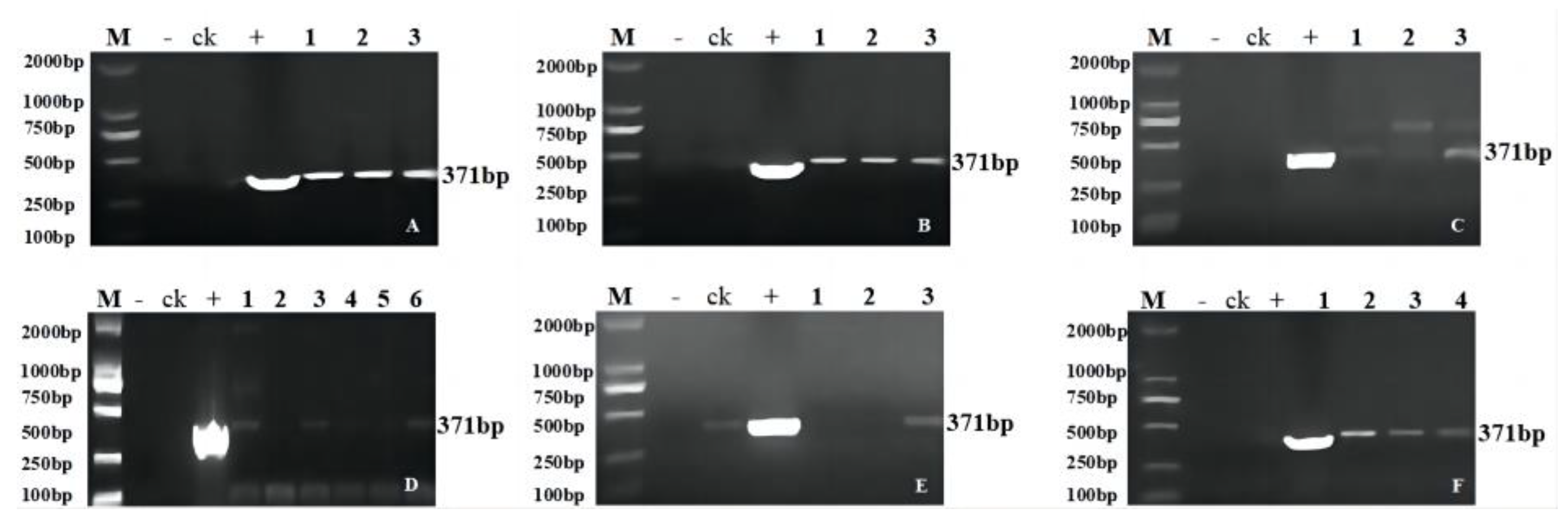
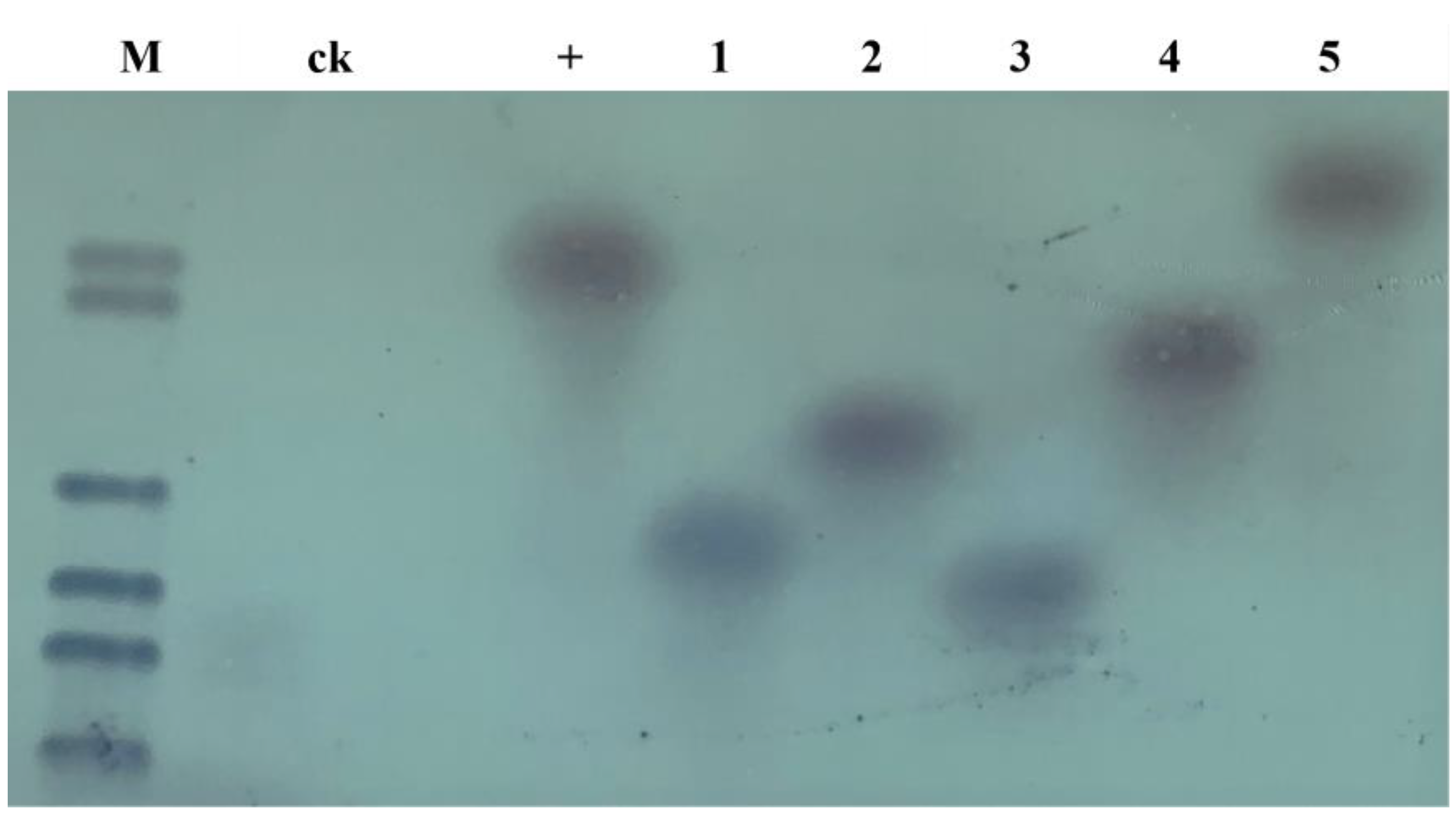
2.2. Molecular Analysis of the Transgenic Progeny
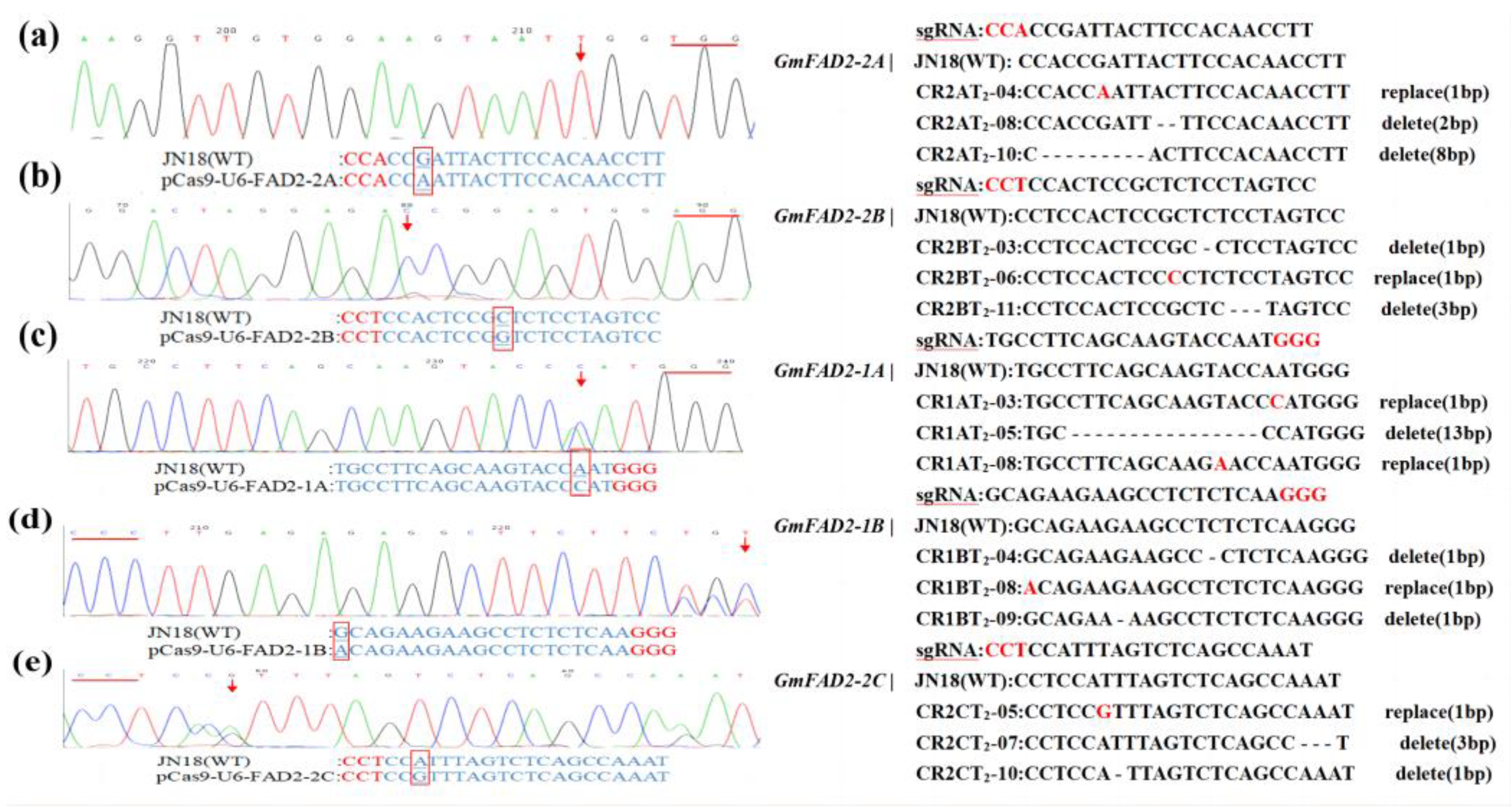
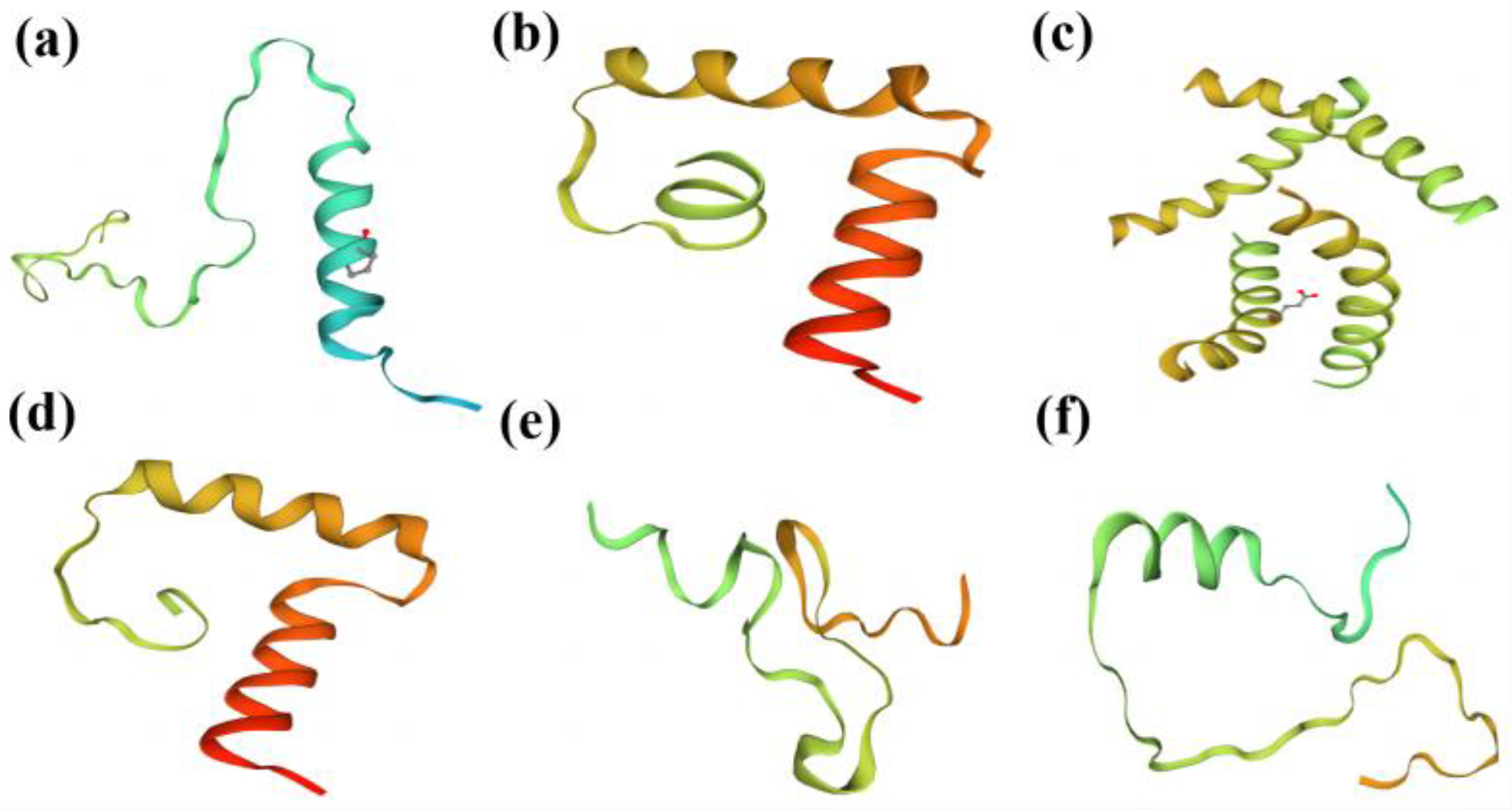
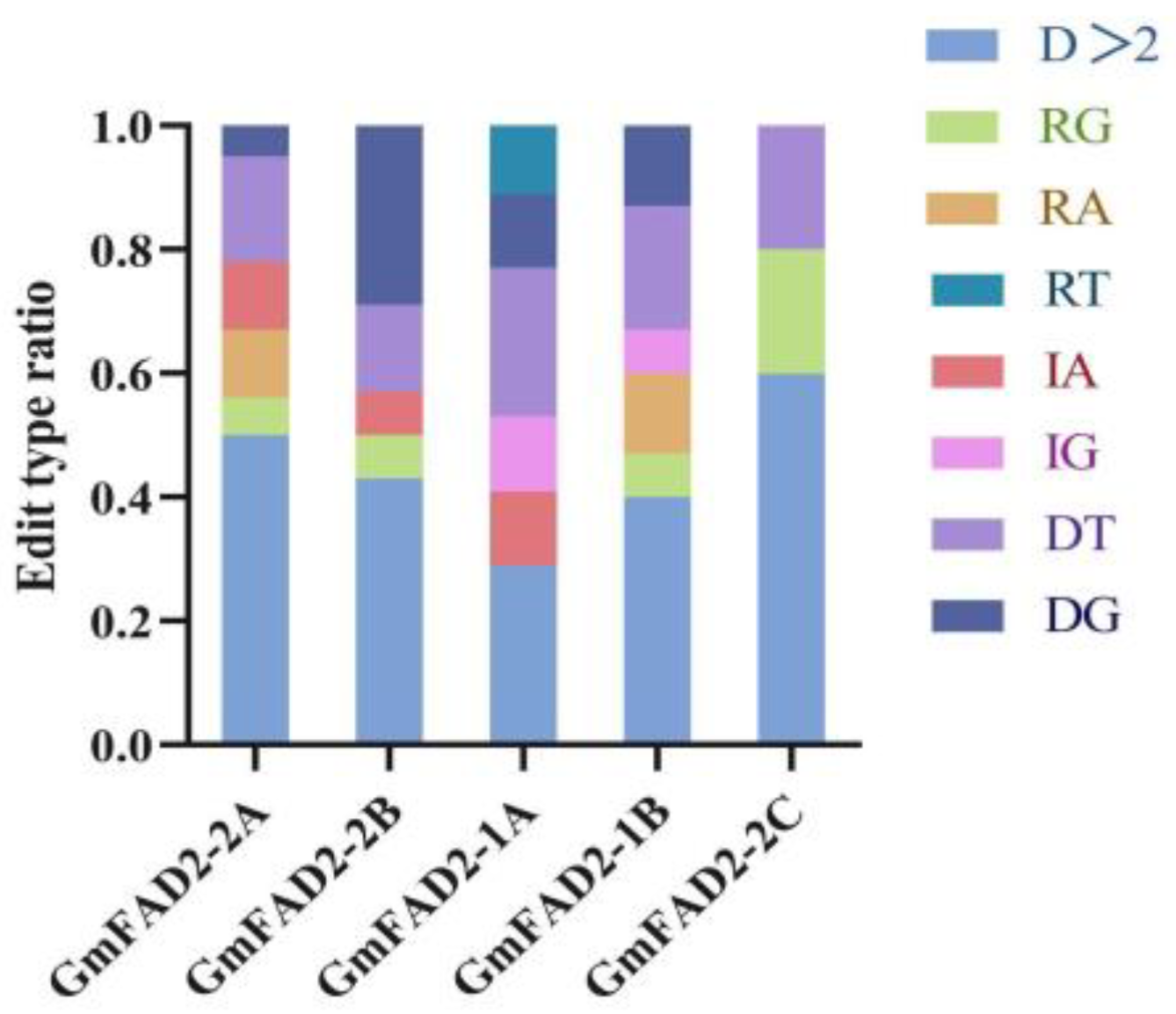
2.3. Sequence Analysis of the Target Gene Mutations in T1 Transgenic Plants
2.4. Phenotypic Identification of T2 Transgenic Soybean Seed
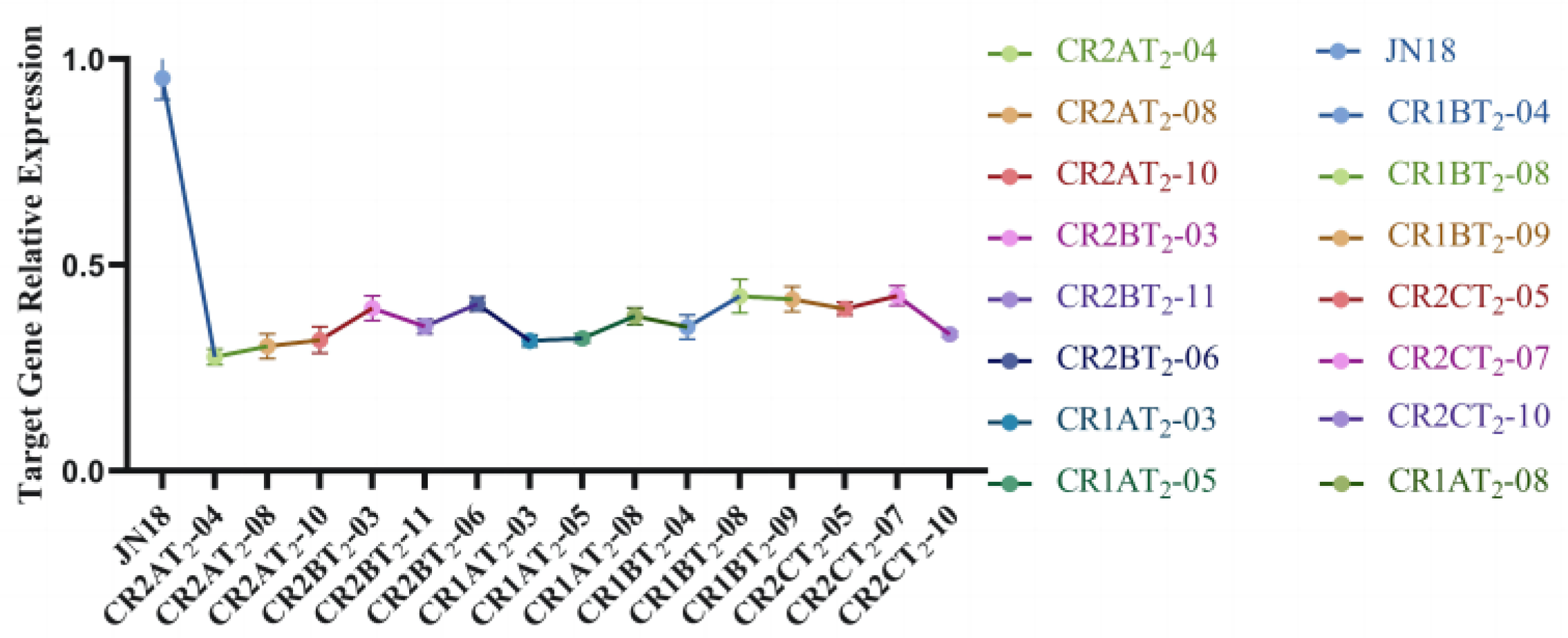
2.4.1. Identification of T2 Transgenic Soybean High Expression Strains
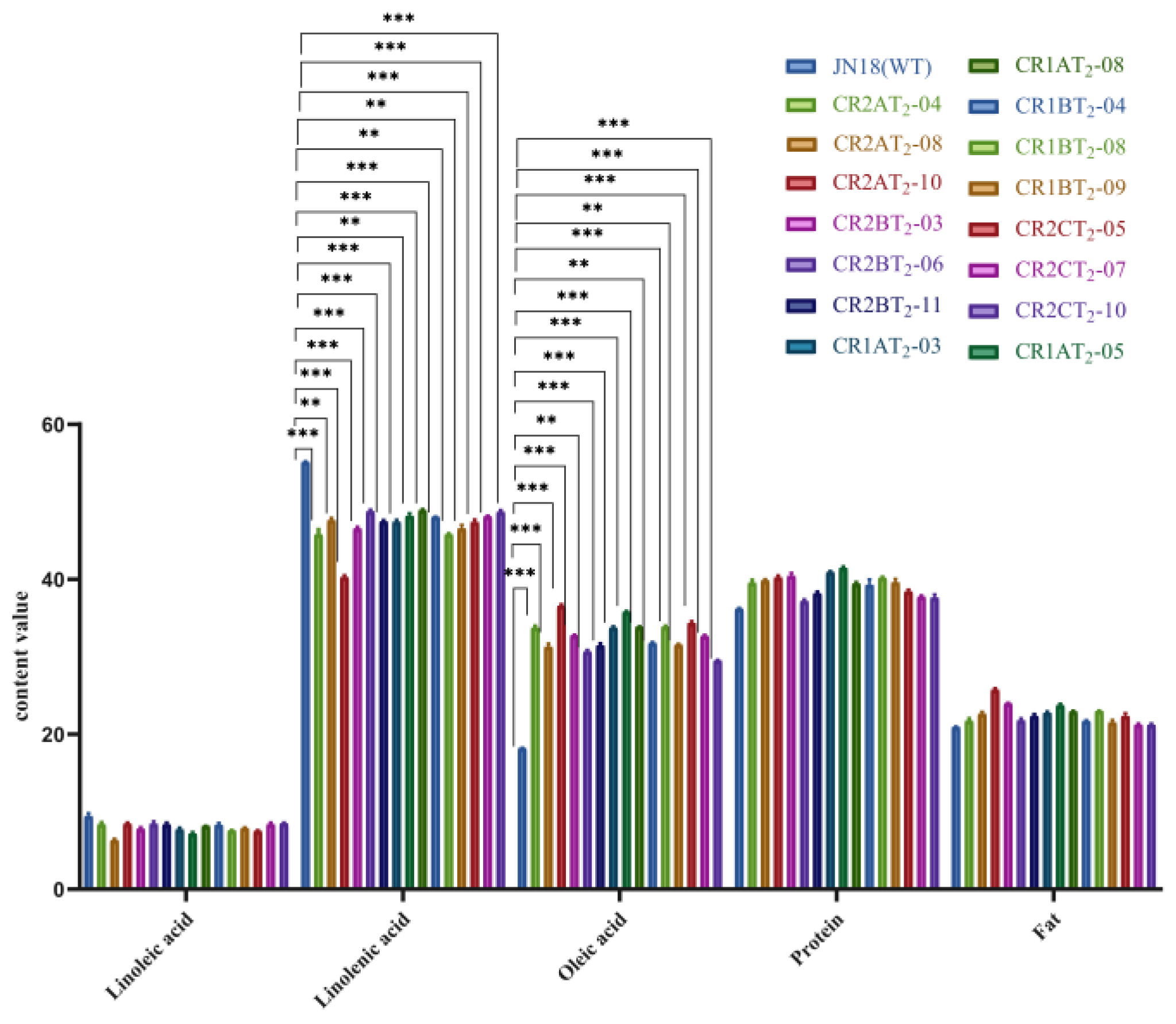
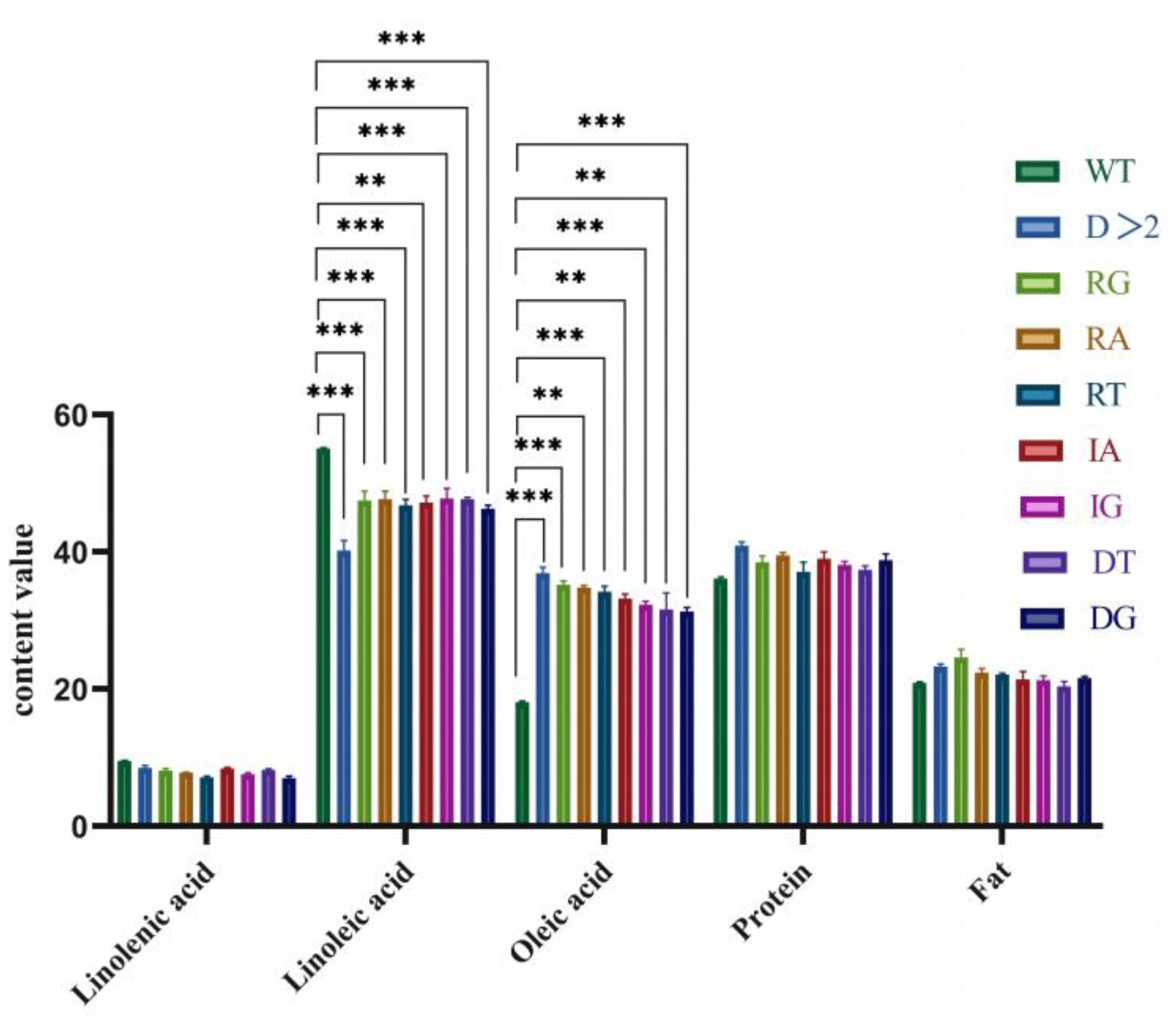
2.4.2. Determination of the Major Fatty Acid Content of T2 Transgenic Soybeans
2.4.3. Determination of Major Agronomic Traits in T2 Transgenic Soybean
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Construction of CRISPR/Cas9 Editing Vectors
4.3. Soybean Genetic Transformation and PCR Analysis of the Progeny of the Transgenic Plants
4.4. Evaluation of Transformation Efficiency and Gene Editing Efficiency
4.5. Southern Blot Analysis of the T2 Transgenic Plants
4.6. Quantitative Real-Time Polymerase Chain Reaction Analysis (q-RT PCR)
4.7. Determining the Primary Characteristics of Quality
4.8. Analysis of the Main Agronomic Traits
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schauberger, B.; Archontoulis, S.; Arneth, A.; Balkovic, J.; Ciais, P.; Deryng, D.; Elliott, J.; Folberth, C.; Khabarov, N.; Muller, C.; et al. Consistent negative response of US crops to high temperatures in observations and crop models. Nat. Commun. 2017, 8, 13931. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Rahman, T.; Song, C.; Su, B.Y.; Yang, F.; Yong, T.; Wu, Y.S.; Zhang, C.; Yang, W. Changes in light environment, morphology, growth and yield of soybean in maize-soybean intercropping systems. Field Crop. Res. 2017, 200, 38–46. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.F.; Zhu, B.G.; Yang, J.X.; Zhou, G.A.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, J.R.J. The meaning of words. Radiology 1998, 208, 829. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.J.; Zhang, Y.L.; Liu, X.; Huang, H.; Zhou, X.E.; Wang, W.L.; Zeng, A.; Zhao, C.Z.; Si, T.; Du, J.M.; et al. Combining chemical and genetic approaches to increase drought resistance in plants. Nat. Commun. 2017, 8, 1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, X.N.; Bi, S.; Qi, B.K.; Wang, Z.J.; Zhang, M.; Li, Y.; Jiang, L.Z. Impact of ultrasonic treatment on an emulsion system stabilized with soybean protein isolate and lecithin: Its emulsifying property and emulsion stability. Food Hydrocoll. 2017, 63, 727–734. [Google Scholar] [CrossRef]
- Tan, Y.Q.; Chang, S.; Zhang, Y. Comparison of alpha-amylase, alpha-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Chen, K.L.; Li, T.D.; Zhang, Y.; Wang, Y.P.; Zhao, Q.; Liu, J.X.; Zhang, H.W.; Liu, C.M.; Ran, Y.D.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.W.; Jiang, L.J.; Gao, Q.; Zhang, J.; Zong, M.; Zhang, H.Y.; Ren, Y.; Guo, S.G.; Gong, G.Y.; Liu, F.; et al. Efficient CRISPR/Cas9-based gene knockout in watermelon. Plant Cell Rep. 2017, 36, 399–406. [Google Scholar] [CrossRef]
- Char, S.N.; Neelakandan, A.K.; Nahampun, H.; Frame, B.; Main, M.; Spalding, M.H.; Becraft, P.W.; Meyers, B.C.; Walbot, V.; Wang, K.; et al. An Agrobacterium-delivered CRISPR/Cas9 system for high-frequency targeted mutagenesis in maize. Plant Biotechnol. J. 2017, 15, 257–268. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Nicolia, A.; Falt, A.S.; Samuelsson, M.; Hofvander, P. Efficient targeted multiallelic mutagenesis in tetraploid potato (Solanum tuberosum) by transient CRISPR-Cas9 expression in protoplasts. Plant Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Showalter, A.M. CRISPR/Cas9 Genome Editing Technology: A Valuable Tool for Understanding Plant Cell Wall Biosynthesis and Function. Front. Plant Sci. 2020, 11, 589517. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.B.; Zhang, L.H.; Zhang, M.; Chu, J.Y.; Xia, Y.Y.; Yang, H.Q.; Liu, L.M.; Chen, X.Z. A CRISPR-Cas9 System-Mediated Genetic Disruption and Multi-fragment Assembly in Starmerella bombicola. ACS Synth. Biol. 2022, 11, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Ueta, R.; Abe, C.; Osakabe, Y.; Osakabe, K. Efficient Multiplex Genome Editing Induces Precise, and Self-Ligated Type Mutations in Tomato Plants. Front. Plant Sci. 2018, 9, 916. [Google Scholar] [CrossRef]
- Do, P.T.; Nguyen, C.X.; Bui, H.T.; Tran, L.; Stacey, G.; Gillman, J.D.; Zhang, Z.; Stacey, M.G. Demonstration of highly efficient dual gRNA CRISPR/Cas9 editing of the homeologous GmFAD2-1A and GmFAD2-1B genes to yield a high oleic, low linoleic and alpha-linolenic acid phenotype in soybean. BMC Plant Biol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Z.; Fu, M.C.; Li, H.; Wang, L.G.; Liu, R.Z.; Liu, Z.J.; Zhang, X.L.; Jin, S.X. High-oleic acid content, nontransgenic allotetraploid cotton (Gossypium hirsutum L.) generated by knockout of GhFAD2 genes with CRISPR/Cas9 system. Plant Biotechnol. J. 2021, 19, 424–426. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Lopes-Caitar, V.S.; Knizia, D.; Cullen, M.A.; Badad, O.; El Baze, A.; Zhou, Z.; Embaby, M.G.; Meksem, J.; Lakhssassi, A.; et al. TILLING-by-Sequencing (+) Reveals the Role of Novel Fatty Acid Desaturases (GmFAD2-2s) in Increasing Soybean Seed Oleic Acid Content. Cells 2021, 10, 1245. [Google Scholar] [CrossRef]
- Cai, Z.D.; Xian, P.Q.; Cheng, Y.B.; Ma, Q.B.; Lian, T.X.; Nian, H.; Ge, L.F. CRISPR/Cas9-mediated gene editing of GmJAGGED1 increased yield in the low-latitude soybean variety Huachun 6. Plant Biotechnol. J. 2021, 19, 1898–1900. [Google Scholar] [CrossRef]
- Wang, T.Y.; Xun, H.W.; Wang, W.; Ding, X.Y.; Tian, H.A.; Hussain, S.; Dong, Q.L.; Li, Y.Y.; Cheng, Y.X.; Wang, C.; et al. Mutation of GmAITR Genes by CRISPR/Cas9 Genome Editing Results in Enhanced Salinity Stress Tolerance in Soybean. Front. Plant Sci. 2021, 12, 2752. [Google Scholar] [CrossRef]
- Liu, X.J.; Yang, J.T.; Song, Y.Y.; Zhang, X.C.; Wang, X.J.; Wang, Z.X. Effects of sgRNA length and number on gene editing efficiency and predicted mutations generated in rice. Crop J. 2022, 10, 577–581. [Google Scholar] [CrossRef]
- Zhang, C.W.; Wang, F.P.; Zhao, S.; Kang, G.T.; Song, J.L.; Li, L.; Yang, J.X. Highly efficient CRISPR-SaKKH tools for plant multiplex cytosine base editing. Crop J. 2020, 8, 418–423. [Google Scholar] [CrossRef]
- Hoshino, T.; Takagi, Y.; Anai, T. Novel GmFAD2-1b mutant alleles created by reverse genetics induce marked elevation of oleic acid content in soybean seeds in combination with GmFAD2-1a mutant alleles. Breed. Sci. 2010, 60, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Song, G.Y.; Jia, M.L.; Chen, K.; Kong, X.C.; Khattak, B.; Xie, C.X.; Li, A.L.; Mao, L. CRISPR/Cas9: A powerful tool for crop genome editing. Crop J. 2016, 4, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Dong, H.R.; Shang, M.Q.; Wang, K.J. CRISPR/Cas systems: The link between functional genes and genetic improvement. Crop J. 2021, 9, 678–687. [Google Scholar] [CrossRef]
- Ma, X.L.; Zhu, Q.L.; Chen, Y.L.; Liu, Y.G. CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Mol. Plant. 2016, 9, 961–974. [Google Scholar] [CrossRef] [Green Version]
- Molla, K.A.; Sretenovic, S.; Bansal, K.C.; Qi, Y.P. Precise plant genome editing using base editors and prime editors. Nat. Plants 2021, 7, 1166–1187. [Google Scholar] [CrossRef]
- Wang, F.P.; Zhang, C.W.; Xu, W.; Yuan, S.; Song, J.L.; Li, L.; Zhao, J.R.; Yang, J.X. Developing high-efficiency base editors by combining optimized synergistic core components with new types of nuclear localization signal peptide. Crop J. 2020, 8, 408–417. [Google Scholar] [CrossRef]
- Yin, K.Q.; Gao, C.X.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Zhou, Z.; Liu, S.M.; Colantonio, V.; AbuGhazaleh, A.; Meksem, K. Characterization of the FAD2 Gene Family in Soybean Reveals the Limitations of Gel-Based TILLING in Genes with High Copy Number. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Nan, H.Y.; Lu, S.J.; Fang, C.; Hou, Z.H.; Yang, C.; Zhang, Q.; Liu, B.H.; Kong, F.J. Molecular breeding of a high oleic acid soybean line by integrating natural variations. Mol. Breed. 2020, 40, 1–10. [Google Scholar] [CrossRef]
- Wu, N.; Lu, Q.; Wang, P.W.; Zhang, Q.; Zhang, J.; Qu, J.; Wang, N. Construction and Analysis of GmFAD2-1A and GmFAD2-2A Soybean Fatty Acid Desaturase Mutants Based on CRISPR/Cas9 Technology. Int. J. Mol. Sci. 2020, 21, 1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.B.; Cheng, Q.; Gan, Z.R.; Hou, Z.H.; Zhang, Y.H.; Li, Y.L.; Li, H.Y.; Nan, H.Y.; Yang, C.; Chen, L.N.; et al. Multiplex CRISPR/Cas9-mediated knockout of soybean LNK2 advances flowering time. Crop J. 2021, 9, 767–776. [Google Scholar] [CrossRef]
- Hu, X.X.; Meng, X.B.; Li, J.Y.; Wang, K.J.; Yu, H. Improving the efficiency of the CRISPR-Cas12a system with tRNA-crRNA arrays. Crop J. 2020, 8, 403–407. [Google Scholar] [CrossRef]
- Zheng, X.L.; Qi, C.Y.; Yang, L.J.; Quan, Q.; Liu, B.L.; Zhong, Z.H.; Tang, X.; Fan, T.T.; Zhou, J.P.; Zhang, Y. The Improvement of CRISPR-Cas9 System with Ubiquitin-Associated Domain Fusion for Efficient Plant Genome Editing. Front. Plant Sci. 2020, 11, 621. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.R.; Ren, C.; Zhang, Z.; Duan, W.; Lecourieux, D.; Li, S.H.; Liang, Z.C. Efficiency Optimization of CRISPR/Cas9-Mediated Targeted Mutagenesis in Grape. Front. Plant Sci. 2019, 10, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wu, S.P.; Hu, C.H.; Yang, Q.S.; Dong, T.; Sheng, O.; Deng, G.M.; He, W.D.; Dou, T.X.; Li, C.Y.; et al. Increased mutation efficiency of CRISPR/Cas9 genome editing in banana by optimized construct. PeerJ 2022, 10, e12664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Q.; Eleouet, M.; Chen, R.; Chen, H.; Zhang, N.; Hu, Y.; Sui, Z. CRISPR/LbCas12a-mediated targeted mutation of Gracilariopsis lemaneiformis (Rhodophyta). Plant Biotechnol. J. 2023, 21, 235–237. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, Y.; Chai, Y. Optimization of CRISPR/LbCas12a-mediated gene editing in Arabidopsis. PLoS ONE 2022, 17, e0265114. [Google Scholar] [CrossRef]
- Jia, H.; Orbovic, V.; Wang, N. CRISPR-LbCas12a-mediated modification of citrus. Plant Biotechnol. J. 2019, 17, 1928–1937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.H.; Wang, W.P.; Ali, S.; Luo, X.; Xie, L.A. CRISPR/Cas9-Mediated Multiple Knockouts in Abscisic Acid Receptor Genes Reduced the Sensitivity to ABA during Soybean Seed Germination. Int. J. Mol. Sci. 2022, 23, 16173. [Google Scholar] [CrossRef]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, C.; Zhang, F.; Mendez, J.; Lozano, Y.; Chatpar, K.; Irish, V.F.; Jacob, Y. Increased efficiency of targeted mutagenesis by CRISPR/Cas9 in plants using heat stress. Plant J. 2018, 93, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Chen, W.D.; Li, Y.S.; Sun, L.; Chai, Y.H.; Chen, H.X.; Nie, H.C.; Huang, C.L. CRISPR/Cas9 Technology and Its Utility for Crop Improvement. Int. J. Mol. Sci. 2022, 23, 10442. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, W.A.; Tsegaw, M.; Zhang, Y.X.; Cao, L.Y. CRISPR-Based Genome Editing: Advancements and Opportunities for Rice Improvement. Int. J. Mol. Sci. 2022, 23, 4454. [Google Scholar] [CrossRef]
- Wang, X.Y.; Lu, J.Y.; Lao, K.W.; Wang, S.K.; Mo, X.W.; Xu, X.T.; Chen, X.M.; Mo, B.X. Increasing the efficiency of CRISPR/Cas9-based gene editing by suppressing RNAi in plants. Sci. China-Life Sci. 2019, 62, 982–984. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Target Sequence | GC Content (%) | The Target Rate |
|---|---|---|---|
| FAD2-2A-Target | AAGGTTGTGGAAGTAATCGGTGG | 45 | 0.5466 |
| FAD2-2B-Target | GGACTAGGAGAGCGGAGTGGAGG | 65 | 0.6594 |
| FAD2-1A-Target | TGCCTTCAGCAAGTACCAATGGG | 45 | 0.6309 |
| FAD2-1B-Target | GCAGAAGAAGCCTCTCTCAAGGG | 50 | 0.7301 |
| FAD2-2C-Target | ATTTGGCTGAGACTAAATGGAGG | 40 | 0.6271 |
| Materials | Oleic Acid (%) | Linoleic Acid (%) | Linolenic Acid (%) | Protein (%) | Fat (%) | Oleic Acid Increase Percentage (%) |
|---|---|---|---|---|---|---|
| JN18-CK | 17.98 c ± 1.16 | 55.25 a ± 0.58 | 8.17 a ± 0.29 | 38.50 b ± 0.83 | 21.85 b ± 1.67 | - |
| CR1AT2 | 34.43 a ± 1.29 | 46.30 bc ± 0.68 | 7.78 a ± 0.52 | 40.16 a ± 0.99 | 23.14 ab ± 0.55 | 91.49 |
| CR1BT2 | 34.02 a ± 1.61 | 46.87 b ± 1.14 | 7.96 a ± 0.40 | 39.14 b ± 0.67 | 22.11 b ± 0.81 | 89.21 |
| CR2AT2 | 32.78 ab ± 3.29 | 44.92 c ± 1.29 | 7.89 a ± 1.16 | 39.56 ab ± 0.36 | 23.41 a ± 2.11 | 82.31 |
| CR2BT2 | 31.59 bc ± 1.03 | 47.62 bc ± 1.12 | 8.22 a ± 0.35 | 38.59 b ± 1.69 | 22.73 ab ± 1.10 | 75.70 |
| CR2CT2 | 32.16 b ± 2.40 | 48.15 b± 0.73 | 8.13 a ± 0.52 | 38.42 b ± 0.39 | 21.57 b ± 0.58 | 78.87 |
| LSD0.05 | 1.61 | 1.88 | 0.49 | 0.76 | 0.95 | - |
| Type of Editing | Oleic Acid (%) | Linoleic Acid (%) | Linolenic Acid (%) | Protein (%) | Fat (%) | Oleic Acid Increase Percentage (%) |
|---|---|---|---|---|---|---|
| JN18-CK | 17.98 d ± 1.16 | 55.25 a ± 0.58 | 8.17 ab ± 0.29 | 38.50 b ± 0.83 | 21.85 b ± 1.67 | - |
| D > 2 | 36.95 a ± 0.85 | 41.22 d ± 1.74 | 8.55 a ± 0.33 | 39.92 a ± 0.57 | 22.30 a ± 1.46 | 105.51% |
| RA | 34.82 bc ± 0.32 | 47.17 bc ± 0.99 | 7.92 b ± 0.51 | 39.52 ab ± 0.39 | 21.61 b ± 0.74 | 93.66% |
| RT | 34.19 c ± 0.82 | 46.82 b ± 0.85 | 7.81 b ± 0.12 | 38.37 b ± 1.47 | 21.03 b ± 0.77 | 90.16% |
| RG | 35.20 b ± 0.56 | 47.53 b ± 1.34 | 8.12 b ± 0.32 | 38.51 b ± 0.93 | 22.47 ab ± 3.06 | 95.77% |
| DT | 31.61 d ± 0.48 | 47.67 b ± 0.29 | 8.20 ab ± 0.17 | 38.42 b ± 0.62 | 22.94 ab ± 1.06 | 75.81% |
| DG | 31.33 d ± 0.60 | 46.33 c ± 0.49 | 7.72 b ± 0.33 | 38.76 b ± 0.99 | 23.03 a ± 0.69 | 74.25% |
| IA | 33.24 d ± 0.64 | 47.65 b ± 1.23 | 8.40 ab ± 0.71 | 39.00 b ± 1.02 | 21.71 b ± 0.82 | 84.87% |
| IG | 32.35 d ± 0.48 | 47.79 b ± 1.50 | 7.89 b ± 0.24 | 38.45 b ± 0.52 | 21.43 b ± 1.42 | 79.92% |
| LSD0.05 | 0.84 | 0.92 | 0.41 | 0.71 | 1.19 | - |
| Materials | Plant Height (cm) | Number of Branches | Number of Nodes | Number of Pods | Total Number of Grains | Single Plant Weight (g) | 100 Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| JN18-CK | 83.28 a ± 1.89 | 5.92 ab ± 0.33 | 18.60 b ± 0.78 | 35.85 c ± 4.69 | 245.86 c ± 3.01 | 41.45 c ± 1.19 | 20.98 ab ± 0.48 |
| CR1AT2 | 84.30 a ± 2.19 | 6.38 a ± 1.40 | 21.50 a ± 1.28 | 47.44 b ± 1.99 | 306.57 ab ± 4.95 | 50.37 b ± 2.14 | 20.78 b ± 0.62 |
| CR1BT2 | 85.13 a ± 1.39 | 6.27 ab ± 0.96 | 20.27 ab ± 0.71 | 50.83 a ± 3.69 | 300.10 b ± 10.13 | 54.90 a ± 1.18 | 21.35 ab ± 0.36 |
| CR2AT2 | 83.60 a ± 3.20 | 6.52 a ± 2.55 | 22.07 a ± 2.18 | 53.57 a ± 4.51 | 310.40 a ± 16.37 | 51.59 b ± 5.10 | 21.47 a ± 0.65 |
| CR2BT2 | 83.13 a ± 0.61 | 6.47 a ± 1.44 | 18.47 b ± 0.85 | 52.67 a ± 4.55 | 298.57 b ± 5.42 | 52.39 b ± 2.93 | 20.61 b ± 0.70 |
| CR2CT2 | 83.18 a ± 1.81 | 5.70 b ± 0.46 | 19.11 b ± 0.64 | 52.23 a ± 2.61 | 303.23 ab ± 5.92 | 52.30 b ± 2.98 | 20.74 b ± 0.85 |
| LSD0.05 | 2.01 | 0.67 | 1.85 | 2.77 | 7.32 | 2.36 | 0.51 |
| Materials | Plant Height (cm) | Number of Branches | Number of Nodes | Number of Pods | Total Number of Grains | Single Plant Weight (g) | 100 Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| JN18-CK | 83.28 b ± 1.89 | 5.92 ab ± 0.33 | 18.60 b ± 0.78 | 35.85 e ± 4.69 | 245.86 e ± 3.01 | 41.45 b ± 1.19 | 20.98 b ± 0.48 |
| D > 2 | 85.13 ab ± 2.85 | 6.03 ab ± 1.70 | 21.26 a ± 2.37 | 49.50 bc ± 5.48 | 315.40 a ± 7.99 | 51.85 a ± 4.89 | 21.15 b ± 0.61 |
| RA | 82.65 b ± 0.81 | 5.65 b ± 0.47 | 20.62 ab ± 1.03 | 46.03 c ± 3.21 | 298.51 bc ± 2.64 | 52.24 a ± 0.52 | 20.56 b ± 0.72 |
| RT | 83.53 b ± 1.40 | 5.85 ab ± 1.44 | 20.96 ab ± 1.99 | 40.95 d ± 1.89 | 290.93 c ± 18.06 | 44.15 b ± 4.09 | 19.61 c ± 0.57 |
| RG | 84.28 ab ± 2.81 | 5.50 b ± 0.72 | 19.86 ab ± 2.47 | 46.62 bc ± 1.86 | 279.80 d ± 17.91 | 50.31 a ± 3.74 | 22.07 a ± 0.61 |
| DT | 83.10 b ± 0.95 | 5.73 b ± 0.41 | 20.23 ab ± 1.51 | 53.73 a ± 1.06 | 299.86 bc ± 7.74 | 52.80 a ± 3.22 | 20.92 b ± 0.75 |
| DG | 85.41 ab ± 1.40 | 6.58 a ± 0.78 | 19.53 b ± 0.83 | 43.56 cd ± 8.55 | 307.12 ab ± 9.20 | 51.63 a ± 9.03 | 21.16 b ± 0.61 |
| IA | 85.62 a ± 3.83 | 4.99 b ± 0.75 | 19.57 b ± 2.46 | 49.56 b ± 1.07 | 297.46 bc ± 16.49 | 52.04 a ± 1.68 | 20.58 b ± 1.53 |
| IG | 85.70 a ± 2.74 | 5.39 b ± 1.13 | 20.25 ab ± 1.91 | 48.38 bc ± 1.42 | 302.78 b ± 4.37 | 44.23 b ± 0.94 | 20.51 b ± 0.89 |
| LSD0.05 | 1.97 | 0.79 | 1.49 | 3.53 | 9.29 | 3.34 | 0.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Li, Z.; Li, Y.; Zhao, Q.; Luan, X.; Wang, L.; Liu, Y.; Liu, H.; Zhang, J.; Yao, D. Effects of Different Gene Editing Modes of CRISPR/Cas9 on Soybean Fatty Acid Anabolic Metabolism Based on GmFAD2 Family. Int. J. Mol. Sci. 2023, 24, 4769. https://doi.org/10.3390/ijms24054769
Zhou J, Li Z, Li Y, Zhao Q, Luan X, Wang L, Liu Y, Liu H, Zhang J, Yao D. Effects of Different Gene Editing Modes of CRISPR/Cas9 on Soybean Fatty Acid Anabolic Metabolism Based on GmFAD2 Family. International Journal of Molecular Sciences. 2023; 24(5):4769. https://doi.org/10.3390/ijms24054769
Chicago/Turabian StyleZhou, Junming, Zeyuan Li, Yue Li, Qiuzhu Zhao, Xinchao Luan, Lixue Wang, Yixuan Liu, Huijing Liu, Jun Zhang, and Dan Yao. 2023. "Effects of Different Gene Editing Modes of CRISPR/Cas9 on Soybean Fatty Acid Anabolic Metabolism Based on GmFAD2 Family" International Journal of Molecular Sciences 24, no. 5: 4769. https://doi.org/10.3390/ijms24054769
APA StyleZhou, J., Li, Z., Li, Y., Zhao, Q., Luan, X., Wang, L., Liu, Y., Liu, H., Zhang, J., & Yao, D. (2023). Effects of Different Gene Editing Modes of CRISPR/Cas9 on Soybean Fatty Acid Anabolic Metabolism Based on GmFAD2 Family. International Journal of Molecular Sciences, 24(5), 4769. https://doi.org/10.3390/ijms24054769