
Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research
 , , , , ,
, , , , ,
Abstract
1. Introduction
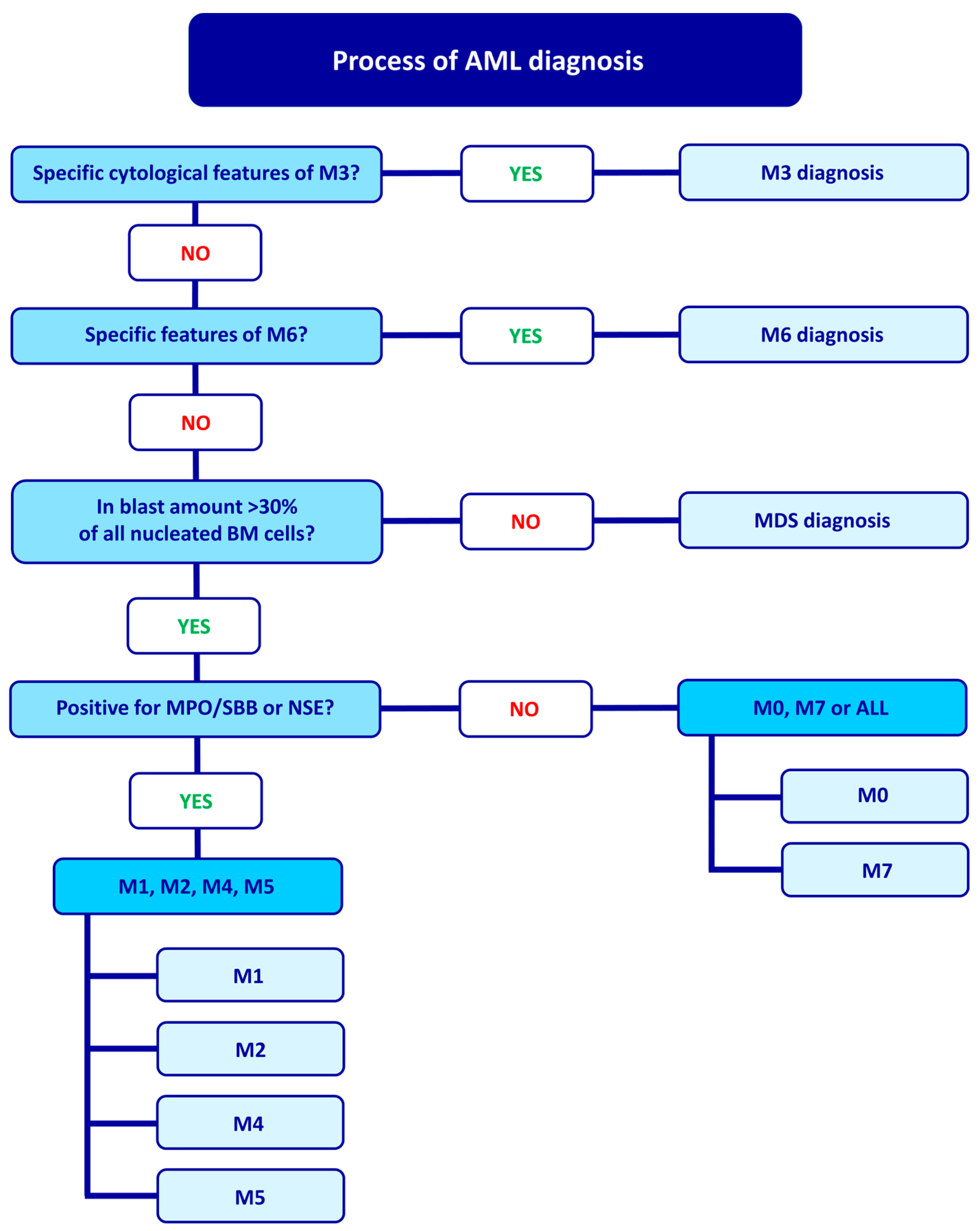
2. FAB Classification and Diagnostic Process
3. WHO Classification
4. AML Cell Lines and Types
4.1. AML with Minimal Differentiation (M0)
4.1.1. Kasumi-3
4.1.2. MOLM-16
4.2. AML without Maturation (M1)
4.2.1. CTS
4.2.2. UoC-M1
4.2.3. KG-1
4.2.4. K-562
4.3. AML with Maturation (M2)
4.3.1. Kasumi-1
4.3.2. Kasumi-6
4.3.3. SKNO-1
4.3.4. HL-60
4.3.5. PLB-985
4.4. Acute Promyelocytic Leukemia (M3)
4.4.1. NB4
4.4.2. PL-21
4.4.3. UF-1
4.4.4. HT93
4.4.5. AP-1060
4.5. Acute Myelomonocytic Leukemia (M4)
4.5.1. OCI-AML2 and OCI-AML3
4.5.2. MUTZ-11
4.5.3. MUTZ-8
4.5.4. MUTZ-3
4.5.5. ME-1
4.6. Acute Monoblastic and Monocytic Leukemia (M5)
4.6.1. THP-1
4.6.2. U-937
4.6.3. MOLM-13 and MV4-11
4.7. Acute Erythroid Leukemia (M6)
4.7.1. HEL
4.7.2. OCI-M1
4.7.3. OCI-M2
4.7.4. F-36P
4.7.5. TF-1
4.7.6. AS-E2
4.8. Acute Megakaryoblastic Leukemia (M7)
4.8.1. CMK
4.8.2. ELF-153
4.8.3. UT-7
4.8.4. M-07
4.8.5. MEG-01
4.8.6. MEGAL
4.9. Cell-Line Markers

{kind=link}
{kind=link}
{kind=link}
| Disease | Cell Line | Markers | Use |
|---|---|---|---|
| M0 | Kasumi-3 | CD2−, cy/smCD3−, CD4+, CD5−, CD7+, CD8−, CD13+, CD14+, CD15−, CD19−, CD20−, CD22+, CD25+, CD33+, CD34+,CD38+, CD56+, cyCD68+, HLA-DR+, c-Kit+ | EVI1 and BET inhibitors research; |
| Drug response in AML; | |||
| Engraftment studies | |||
| MOLM-16 | CD3−, CD9+, CD13+, CD19−, CD22+ CD31+, CD33+, CD34+, CD36+, CD38+, CD41+, CD47+, CD56+, CD61+, CD62P+, CD63+, CD71+, CD110+, CD117+, CD119+ CD151+, CD235A+, thrombospondin+, vWf+, fibrinogen+, HLA-DR- | t(6;8) (q21;q24.3) model; | |
| PIM/FLT3 signaling; | |||
| JAK2 V617F function research; | |||
| PMS2 and RSPH10B2 deletion | |||
| M1 | CTS | CD1+, CD2+, CD3+, CD4+, CD5+, CD7+, CD8+, CD10+, CD13+, CD14+, CD19+, CD20+, CD25+, CD33+, CD34+, HLA-DR+, D2-10+, P2+, HPCA-1+ | t(6;11) (q27;q23) model; |
| KMT2/AF6 research model; | |||
| GM-CSF and G-CSF differentiation; | |||
| Pluripotent stem cell research | |||
| UoC-M1 | CD7+, CD24+, CD34+, CD38+, CD45+, HLA-DR+ CD61+ | Monosomy 7 and 5q loss model; | |
| High KMT2A mRNA level | |||
| KG1 | CD3−, CD13+, CD14−, CD15+, CD19−, CD33+, CD34+, HLA-DR+ | Cell maturation studies; | |
| KMT2A and WT DNMT3A research; | |||
| Toxicology and drug testing; | |||
| Macrophage differentiation; | |||
| K-562 | CD3−, CD14−, CD15+, CD19−, CD33+, CD71+, CD235a+ | BCR-ABL1 fusion, Ph1 chromosome; | |
| Platelet-formation; | |||
| p53-deficient | |||
| M2 | Kasumi-1 | CD3−, CD4+, CD13+, CD14−, CD15+, CD19−, CD33+, CD34+, CD38+, CD71+, HLA-DR+ | t(8;21) model; |
| RUNX1-RUNX1T1 fusion research; | |||
| c-kit, TP53 mutations; | |||
| Granulocytic and macrophage differentiation; | |||
| Il-5 and TPA-induced differentiation; | |||
| Kasumi-6 | CD3−, CD4−, CD13+, CD14−, CD19−, CD33+, CD34−, cyCD68−, HLA-DR+ | FLT3, CEBPA, TP53 mutations; | |
| Model for differentiation research | |||
| TPA-induced differentiation | |||
| SKNO-1 | CD3−, CD4+, CD13+, CD14−, CD15−, CD19-, CD33+ CD1, CD2, CD3, CD4, CD5, CD6, CD7, CD8, CD11, CD13, CD14, CD15, CD19, CD20, CD33, CD34, HLA-DR | t(8;21) (q22;q22) research; | |
| RUNX1/RUNX1T1 fusion; | |||
| Myeloid leukemogenesis studies; | |||
| HL-60 | CD3−, CD4+, CD13+, CD14−, CD15+, CD19−, CD33+, CD34−, HLA-DR- | t(15;17) model; | |
| Granulocytic and mononuclear maturation; Chemotherapeutics influence; | |||
| Proliferation, apoptosis, and cell cycle study; | |||
| Chemotactic response; | |||
| miRNA studies | |||
| PLB-985 | CD3−, CD4+, CD13+, CD14−, CD15+, CD19−, CD33+, CD34− | Granulocytic, monocytic, macrophage maturation, proliferation; | |
| Neutrophil differentiation; | |||
| Maturation studies of cells in early stage | |||
| M3 | NB4 | CD3−, CD4+, CD11b−, CD13+, CD14−, CD15+, CD19−, CD33+, CD34−, CD38+, HLA-DR- | ATRA resistance mechanisms |
| PML-RARAPro900Ser mutation; | |||
| Retinoic acid, DMSO, TPA differentiation; | |||
| Drug screening | |||
| PL-21 | CD3−, CD4 (+), CD14−, CD15+, CD19−, CD33+, cyCD68+, HLA-DR- | Lack of t(15;17) | |
| KRAS, FLT3 mutations and WT P53; | |||
| Kinase inhibitors studies | |||
| UF-1 | CD3−, CD4−, CD5−, CD8−, CD11b−, CD10−, CD7+, CD13+, CD19−, CD20−, CD33+, CD34−, CD38+, CD41− [82] | WT RARA; | |
| PML-RARA research; | |||
| ATRA-resistance studies; | |||
| Multi-drug screening with ATRA | |||
| HT93 | CD3−, CD19−, CD33+, CD34+, cyCD68+, HLA-DR− | t(15;17) and t(1;12) model with PML-RARA and | |
| ETV6-ABL2 fusion; | |||
| TP53 mutation; | |||
| Differentiation, proliferation, cytokine studies | |||
| AP-1060 | CD3−, CD14−, CD15+, CD19−, CD33+, cyCD68+, HLA-DR− | t(15;17) and unique t(3;14) model; | |
| PML-RARAPro900Leu mutation; | |||
| ATRA and ATO resistance; | |||
| Cytokine-dependent growth research; | |||
| ETV6-NTRK3 fusion model; | |||
| Neutrophil maturation | |||
| M4 | OCI-AML2 | CD3−, CD4+, CD13+, CD15+, CD19−, CD33+, CD34−, cyCD68+, HLA-DR+ | Mutated DNMT3A role in leukemogenesis; |
| xenograft models | |||
| OCI-AML3 | CD3−, CD4+, CD13+, CD14−, CD15+, CD19−, cyCD68+, HLA-DR- | Mutated DNMT3A role in leukemogenesis; xenograft models; | |
| NPM1 mutation; | |||
| MUTZ-11 | CD4+, CD7+, CD13+, CD15+, CD33+, CD65+, CD68+ [102] | Response to cytokines; | |
| Dendritic cell myeloid differentiation; | |||
| KMT2A and FLT3 mutations | |||
| MUTZ-8 | CD3−, CD4−, CD13+, CD19−, CD33+, CD34+, HLA-DR+ | t(5;11) model; | |
| JAK2 V617F mutation; | |||
| Cytokine response | |||
| MUTZ-3 | CD3−, CD4+, CD5−, CD7−, CD8−, CD13+, CD14+, CD15+, CD19−, CD34+, HLA-DR+ | Role of FLT3 in leukemia pathogenesis; Cytokine response; | |
| Dendritic cell myeloid differentiation | |||
| M5 | THP-1 | CD3−, CD4+, CD13+, CD15+, CD19−, CD34−, cyCD68+, HLA-DR+ | t(9;11) (p22;q23) with KMT2A/MLLT3(AF9) fusion model; |
| Immune and inflammatory response; inflammation; | |||
| Susceptible to genetic modifications; | |||
| Skin sensitization model; | |||
| Cytokine response; | |||
| U-937 | CD3−, CD4+, CD14−, CD15+, CD19−, CD33+, CD34−, CD54+ | Monocyte and macrophage differentiation; Response to ROS; | |
| Skin sensitization model; | |||
| Model for cell apoptotic disintegration | |||
| MOLM-13 | CD3−, CD4+, CD14−, CD15+, CD19−, CD33+, CD34−, CD68+, HLA-DR− | Drug resistance research; Leukemia xenograft models; | |
| WT TP53, FLT3-ITD+ in AML | |||
| MV4-11 | CD3−, CD4+, CD5−, CD8−, CD10−, CD14−, CD15+, CD19−, CD33+, CD34− | t(4;11) (q21;q23) model; | |
| Mechanisms of FLT3-ITD+ AML Leukemia; Xenograft models; | |||
| WT TP53, FLT3-ITD+ in AML | |||
| M6 | HEL | CD3−, CD13+, CD14−, CD19−, CD33+, CD41a+, CD71+, CD235a+ | JAK/STAT signaling pathway, |
| Differential globins expression | |||
| JAK2 mutation | |||
| OCI-M1 | CD3−, CD4−, CD15+, CD19−, CD33+, CD34−, CD41−, CD42−, CD71+, HLA-DR+ | Model for the EPO receptors studies, | |
| OCI-M2 | CD3+, CD14+, CD19−, CD33 (+), CD71+ | Studying NFkB inhibitors; | |
| NKX2-4 expression | |||
| F-36P | CD3−, CD13+, CD14−, CD15−, CD19−, CD33+, CD34+, CD41−, CD42−, CD71+, CD235a+ | Study of IL-3 and GM-CSF dependence; | |
| Primitive progenitor cell model; | |||
| Myeloid cell differentiation; | |||
| Oncogenesis | |||
| TF-1 | CD3−, CD13+, CD14−, CD15−, CD19−, CD33+, CD34+, CD71+, HLA-DR+ | TNFR2 expression; | |
| TPA macrophage differentiation; | |||
| Cytokines response; | |||
| Anti-IL-6R nanobody ALX-0061 | |||
| AS-E2 | CD2−, CD3−, CD10−, CD11b+, CD13+, CD19−, CD25−, CD33+, CD36+, CD41−, Glycophorin A+, CD71+ [199] | Erythroid progenitor cells; | |
| EPO-dependent growth; | |||
| GATA-1 expression | |||
| M7 | CMK | CD3−, CD13+, CD14−, CD15−, CD19−, CD33+, CD34+, CD71+, CD235a+ | der(17)t(11:17) model; |
| JAK2 V617F mutation; | |||
| Megakaryocytopoiesis research; | |||
| ELF-153 | CD3−, CD4+, CD13+, CD14−, CD15−, CD19−, CD33+, CD34+, HLA-DR+ | High GATA level; | |
| Megakaryocytopoiesis research | |||
| UT-7 | CD3−, CD13+, CD14−, CD15−, CD19−, CD33+, CD34−, cyCD68+ | Response to cytokines; | |
| EPO-response; | |||
| GATA expression, | |||
| M-07 | Non disclosed | Cytokine response | |
| M-07e | CD3−, CD13+, CD14−, CD19−, CD33+, HLA-DR- | Cytokine response | |
| MEG-01 | CD3−, CD13+, CD15+, CD19−, CD33+ | t(9;22) model; | |
| BCR/CBL fusion; | |||
| Megakaryocytic differentiation/maturation | |||
| MEGAL | CD3−, CD13−, CD14−, CD19−, CD33+, CD34+, CD71+, CD235a− | SET-NUP214 fusion |
4.10. Controls from Healthy Donors
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J.M.; Catovsky, D.; Daniel, M.T.; Flandrin, G.; Galton, D.A.; Gralnick, H.R.; Sultan, C. Proposals for the Classification of the Acute Leukaemias. French-American-British (FAB) Co-Operative Group. Br. J. Haematol. 1976, 33, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.M.; Catovsky, D.; Daniel, M.T.; Flandrin, G.; Galton, D.A.; Gralnick, H.R.; Sultan, C. Proposed Revised Criteria for the Classification of Acute Myeloid Leukemia. A Report of the French-American-British Cooperative Group. Ann. Intern. Med. 1985, 103, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Naeim, F.; Rao, P.N. Chapter 11—Acute Myeloid Leukemia. In Hematopathology; Naeim, F., Rao, P.N., Grody, W.W., Eds.; Academic Press: Oxford, UK, 2008; pp. 207–255. [Google Scholar] [CrossRef]
- Ladines-Castro, W.; Barragán-Ibañez, G.; Luna-Pérez, M.A.; Santoyo-Sánchez, A.; Collazo-Jaloma, J.; Mendoza-García, E.; Ramos-Peñafiel, C.O. Morphology of Leukaemias. Rev. Médica Hosp. Gen. México 2016, 79, 107–113. [Google Scholar] [CrossRef]
- Khoury, J.D.; Solary, E.; Abla, O.; Akkari, Y.; Alaggio, R.; Apperley, J.F.; Bejar, R.; Berti, E.; Busque, L.; Chan, J.K.C.; et al. The 5th Edition of the World Health Organization Classification of Haematolymphoid Tumours: Myeloid and Histiocytic/Dendritic Neoplasms. Leukemia 2022, 36, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, H.; Abdullah, W.Z.; Yong, A.C.; Ahmed, S.A.; Abdullah, A.D.; Baba, A.A.; Ankathil, R.; Husin, A.; Hussein, A.R.; Mustaffa, R.; et al. A Review of AML Classification: A Single Institution Experience in a Developing Country. J. Hematopathol. 2014, 7, 3–8. [Google Scholar] [CrossRef]
- Dozzo, A.; Galvin, A.; Shin, J.-W.; Scalia, S.; O’Driscoll, C.M.; Ryan, K.B. Modelling Acute Myeloid Leukemia (AML): What’s New? A Transition from the Classical to the Modern. Drug Deliv. Transl. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Pollak, A.; Leavitt, R.D.; Testa, J.R.; Schiffer, C.A. Minimally Differentiated Acute Nonlymphocytic Leukemia: A Distinct Entity. Blood 1987, 70, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.L.; Hoyer, J.D.; Kurtin, P.J.; Dewald, G.W.; Hanson, C.A. Acute Myeloid Leukemia with Minimal Differentiation. A Multiple Parameter Study. Am. J. Clin. Pathol. 1998, 109, 32–38. [Google Scholar] [CrossRef]
- Shanmugam, V.; Dorfman, D.M. Acute Myeloid Leukemia with Minimal Differentiation (AML M0) Mimicking Acute Lymphoblastic Leukemia. Am. J. Hematol. 2019, 94, 955–956. [Google Scholar] [CrossRef]
- Asou, H.; Tashiro, S.; Hamamoto, K.; Otsuji, A.; Kita, K.; Kamada, N. Establishment of a Human Acute Myeloid Leukemia Cell Line (Kasumi-1) With 8;21 Chromosome Translocation. Blood 1991, 77, 2031–2036. [Google Scholar] [CrossRef]
- Kasai, F.; Asou, H.; Ozawa, M.; Kobayashi, K.; Kuramitsu, H.; Satoh, M.; Kohara, A.; Kaneko, Y.; Kawamura, M. Kasumi Leukemia Cell Lines: Characterization of Tumor Genomes with Ethnic Origin and Scales of Genomic Alterations. Hum. Cell 2020, 33, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Wang, J. EVI1 in Leukemia and Solid Tumors. Cancers 2020, 12, 2667. [Google Scholar] [CrossRef]
- Birdwell, C.; Fiskus, W.; Kadia, T.M.; DiNardo, C.D.; Mill, C.P.; Bhalla, K.N. EVI1 Dysregulation: Impact on Biology and Therapy of Myeloid Malignancies. Blood Cancer J. 2021, 11, 64. [Google Scholar] [CrossRef]
- Chin, D.; Pietsch, C.; McCabe, F.; Chippari, S.; Kaiser, E.; Hanson, R.; Lubomirski, M. Abstract 90: Development of a Systemic Kasumi-3 Acute Myeloid Leukemia Model in NSG Mice. Cancer Res. 2014, 74 (Suppl. S19), 90. [Google Scholar] [CrossRef]
- Matsuo, Y.; Drexler, H.G.; Kaneda, K.; Kojima, K.; Ohtsuki, Y.; Hara, M.; Yasukawa, M.; Tanimoto, M.; Orita, K. Megakaryoblastic Leukemia Cell Line MOLM-16 Derived from Minimally Differentiated Acute Leukemia with Myeloid/NK Precursor Phenotype. Leuk. Res. 2003, 27, 165–171. [Google Scholar] [CrossRef]
- Kaashoek, J.G.; Mout, R.; Falkenburg, J.H.; Willemze, R.; Fibbe, W.E.; Landegent, J.E. Cytokine Production by the Bladder Carcinoma Cell Line 5637: Rapid Analysis of MRNA Expression Levels Using a CDNA-PCR Procedure. Lymphokine Cytokine Res. 1991, 10, 231–235. [Google Scholar] [PubMed]
- Bruyere, H.; Al Moosawi, M. t(6;8) (p21;q24) MYC/SUPT3H. Atlasgeneticsoncology.org. Available online: https://atlasgeneticsoncology.org/haematological/2987/t(6;8)p21;q24) (accessed on 13 December 2022).
- Czardybon, W.; Windak, R.; Dolata, I.; Salwińska, M.; Szydlowski, M.; Sewastianik, T.; Białopiotrowicz, E.; Mądro, E.; Lech-Marańda, E.; Budziszewska, B.K.; et al. Abstract 1749: Preclinical Characterization of SEL24-B489, a Dual PIM/FLT3 Inhibitor for the Treatment of Hematological Malignancies. Cancer Res. 2014, 74 (Suppl. S19), 1749. [Google Scholar] [CrossRef]
- Quentmeier, H.; Reinhardt, J.; Zaborski, M.; Drexler, H.G. FLT3 Mutations in Acute Myeloid Leukemia Cell Lines. Leukemia 2003, 17, 120–124. [Google Scholar] [CrossRef]
- Gonçalves, E.; Poulos, R.C.; Cai, Z.; Barthorpe, S.; Manda, S.S.; Lucas, N.; Beck, A.; Bucio-Noble, D.; Dausmann, M.; Hall, C.; et al. Pan-Cancer Proteomic Map of 949 Human Cell Lines. Cancer Cell 2022, 40, 835–849.e8. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, H.; Sato, T.; Hayashi, Y.; Enomoto, Y.; Takayama, J.; Ohira, M.; Seto, M.; Ueda, R.; Fuse, A.; Niimi, H. A Novel Human Leukaemic Cell Line, CTS, Has a t(6;11) Chromosomal Translocation and Characteristics of Pluripotent Stem Cells. Br. J. Haematol. 1996, 95, 306–318. [Google Scholar] [CrossRef]
- Allen, R.J.; Smith, S.D.; Moldwin, R.L.; Lu, M.M.; Giordano, L.; Vignon, C.; Suto, Y.; Harden, A.; Tomek, R.; Veldman, T.; et al. Establishment and Characterization of a Megakaryoblast Cell Line with Amplification of MLL. Leukemia 1998, 12, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Koeffler, H.P.; Golde, D.W. Acute Myelogenous Leukemia: A Human Cell Line Responsive to Colony-Stimulating Activity. Science 1978, 200, 1153–1154. [Google Scholar] [CrossRef] [PubMed]
- Koya, J.; Kataoka, K.; Sato, T.; Bando, M.; Kato, Y.; Tsuruta-Kishino, T.; Kobayashi, H.; Narukawa, K.; Miyoshi, H.; Shirahige, K.; et al. DNMT3A R882 Mutants Interact with Polycomb Proteins to Block Haematopoietic Stem and Leukaemic Cell Differentiation. Nat. Commun. 2016, 7, 10924. [Google Scholar] [CrossRef]
- Rau, R.E.; Rodriguez, B.A.; Luo, M.; Jeong, M.; Rosen, A.; Rogers, J.H.; Campbell, C.T.; Daigle, S.R.; Deng, L.; Song, Y.; et al. DOT1L as a Therapeutic Target for the Treatment of DNMT3A-Mutant Acute Myeloid Leukemia. Blood 2016, 128, 971–981. [Google Scholar] [CrossRef]
- Mouly, E.; Planquette, C.; Rousseau, E.; Delansorne, R. Abstract 1890: Inecalcitol Respectively Induces or Increases CD38 Expression at the Surface of CD38- or CD38+ AML Cell Lines Representative of All 9 FAB Subtypes except M6. Cancer Res. 2018, 78 (Suppl. S13), 1890. [Google Scholar] [CrossRef]
- Sarde, A.; Eckard, S.; Mei, L.; Ruegg, C.; Chun, P.; Smith, V. Selectivity of T Cell Engager AMV564 Against Different Leukemic Blast Populations and Potential Application for Patient Selection. Blood 2020, 136, 25–26. [Google Scholar] [CrossRef]
- Leung, K.K.; Nguyen, A.; Shi, T.; Tang, L.; Ni, X.; Escoubet, L.; MacBeth, K.J.; DiMartino, J.; Wells, J.A. Multi-Omics Analysis of AML Cells Treated with Azacitidine Reveals Highly Variable Cell Surface Proteome Remodeling. bioRxiv 2018, 369322. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, F. Circular RNA CircCRKL Inhibits the Proliferation of Acute Myeloid Leukemia Cells via the MiR-196a-5p/MiR-196b-5p/P27 Axis. Bioengineered 2021, 12, 7704–7713. [Google Scholar] [CrossRef]
- Koeffler, H.P.; Billing, R.; Lusis, A.J.; Sparkes, R.; Golde, D.W. An Undifferentiated Variant Derived from the Human Acute Myelogenous Leukemia Cell Line (KG-1). Blood 1980, 56, 265–273. [Google Scholar] [CrossRef]
- Li, Y.; Feng, Y.; Si, X.; Zhao, C.; Wang, F.; Niu, X. Genetic Expression Screening of Arsenic Trioxide-Induced Cytotoxicity in KG-1a Cells Based on Bioinformatics Technology. Front. Genet. 2021, 12, 654826. [Google Scholar] [CrossRef]
- Leung, K.K.; Nguyen, A.; Shi, T.; Tang, L.; Ni, X.; Escoubet, L.; MacBeth, K.J.; DiMartino, J.; Wells, J.A. Multiomics of Azacitidine-Treated AML Cells Reveals Variable and Convergent Targets That Remodel the Cell-Surface Proteome. Proc. Natl. Acad. Sci. USA 2019, 116, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Alloush, H.; Smith, M.A.; Hill, P.J.; Salisbury, V. A Rapid Assay of Cytosine Arabinoside Uptake and Metabolism by Acute Myeloid Leukaemic Cells Using a Bioluminescent Bacterial Biosensor. Blood 2007, 110, 4308. [Google Scholar] [CrossRef]
- St Louis, D.C.; Woodcock, J.B.; Franzoso, G.; Blair, P.J.; Carlson, L.M.; Murillo, M.; Wells, M.R.; Williams, A.J.; Smoot, D.S.; Kaushal, S.; et al. Evidence for Distinct Intracellular Signaling Pathways in CD34+ Progenitor to Dendritic Cell Differentiation from a Human Cell Line Model. J. Immunol. 1999, 162, 3237–3248. [Google Scholar] [CrossRef]
- Cejas, P.J.; Carlson, L.M.; Kolonias, D.; Zhang, J.; Lindner, I.; Billadeau, D.D.; Boise, L.H.; Lee, K.P. Regulation of RelB Expression during the Initiation of Dendritic Cell Differentiation. Mol. Cell. Biol. 2005, 25, 7900–7916. [Google Scholar] [CrossRef]
- Andersson, A.; Edén, P.; Lindgren, D.; Nilsson, J.; Lassen, C.; Heldrup, J.; Fontes, M.; Borg, Å.; Mitelman, F.; Johansson, B.; et al. Gene Expression Profiling of Leukemic Cell Lines Reveals Conserved Molecular Signatures among Subtypes with Specific Genetic Aberrations. Leukemia 2005, 19, 1042–1050. [Google Scholar] [CrossRef]
- Jensen, H.A.; Yourish, H.B.; Bunaciu, R.P.; Varner, J.D.; Yen, A. Induced Myelomonocytic Differentiation in Leukemia Cells Is Accompanied by Noncanonical Transcription Factor Expression. FEBS Open Bio. 2015, 5, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Lozzio, C.B.; Lozzio, B.B. Human Chronic Myelogenous Leukemia Cell-Line With Positive Philadelphia Chromosome. Blood 1975, 45, 321–334. [Google Scholar] [CrossRef]
- Raghuvar Gopal, D.V.; Narkar, A.A.; Badrinath, Y.; Mishra, K.P.; Joshi, D.S. Betulinic Acid Induces Apoptosis in Human Chronic Myelogenous Leukemia (CML) Cell Line K-562 without Altering the Levels of Bcr-Abl. Toxicol. Lett. 2005, 155, 343–351. [Google Scholar] [CrossRef]
- Drexler, H.G.; Quentmeier, H.; MacLeod, R.A.F.; Uphoff, C.C.; Hu, Z.-B. Leukemia Cell Lines: In Vitro Models for the Study of Acute Promyelocytic Leukemia. Leuk. Res. 1995, 19, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Kurkowiak, M.; Pępek, M.; Machnicki, M.M.; Solarska, I.; Borg, K.; Rydzanicz, M.; Stawiński, P.; Płoski, R.; Stokłosa, T. Genomic Landscape of Human Erythroleukemia K562 Cell Line, as Determined by next-Generation Sequencing and Cytogenetics. Acta Haematol. Pol. 2017, 48, 343–349. [Google Scholar] [CrossRef]
- Fan, Y.; Lu, H.; An, L.; Wang, C.; Zhou, Z.; Feng, F.; Ma, H.; Xu, Y.; Zhao, Q. Effect of Active Fraction of Eriocaulon Sieboldianum on Human Leukemia K562 Cells via Proliferation Inhibition, Cell Cycle Arrest and Apoptosis Induction. Environ. Toxicol. Pharmacol. 2016, 43, 13–20. [Google Scholar] [CrossRef]
- Duncan, M.T.; DeLuca, T.A.; Kuo, H.-Y.; Yi, M.; Mrksich, M.; Miller, W.M. SIRT1 Is a Critical Regulator of K562 Cell Growth, Survival, and Differentiation. Exp. Cell Res. 2016, 344, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Koeffler, H.P.; Golde, D.W. Human Myeloid Leukemia Cell Lines: A Review. Blood 1980, 56, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kim, S.H.; Zhou, D.-C.; Ding, W.; Paietta, E.; Guidez, F.; Zelent, A.; Ramesh, K.H.; Cannizzaro, L.; Warrell, R.P.; et al. Acute Promyelocytic Leukemia Cell Line AP-1060 Established as a Cytokine-Dependent Culture from a Patient Clinically Resistant to All-Trans Retinoic Acid and Arsenic Trioxide. Leukemia 2004, 18, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Matsuo, Y.; MacLeod, R.A.F. Malignant Hematopoietic Cell Lines: In Vitro Models for the Study of Erythroleukemia. Leuk. Res. 2004, 28, 1243–1251. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Wang, H.; Qin, L.; Feng, A.; Qi, D.; Wang, H.; Zhao, Y.; Kong, L.; Wang, H.; et al. JMJD1C Regulates Megakaryopoiesis in In Vitro Models through the Actin Network. Cells 2022, 11, 3660. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Miyoshi, H.; Shimizu, K.; Kozu, T.; Maseki, N.; Kaneko, Y.; Ohki, M. T(8;21) Breakpoints on Chromosome 21 in Acute Myeloid Leukemia Are Clustered within a Limited Region of a Single Gene, AML1. Proc. Natl. Acad. Sci. USA 1991, 88, 10431–10434. [Google Scholar] [CrossRef]
- Nerlov, C. C/EBPα Mutations in Acute Myeloid Leukaemias. Nat. Rev. Cancer 2004, 4, 394–400. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Chávez-Valencia, V.; Gómez-Guijosa, M.Á.; Cortes-Penagos, C. Acute Myeloid Leukemia—Genetic Alterations and Their Clinical Prognosis. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 328–339. [Google Scholar]
- Chi, Y.; Lindgren, V.; Quigley, S.; Gaitonde, S. Acute Myelogenous Leukemia with t(6;9) (P23;Q34) and Marrow Basophilia: An Overview. Arch. Pathol. Lab. Med. 2008, 132, 1835–1837. [Google Scholar] [CrossRef]
- Thiede, C.; Steudel, C.; Mohr, B.; Schaich, M.; Schäkel, U.; Platzbecker, U.; Wermke, M.; Bornhäuser, M.; Ritter, M.; Neubauer, A.; et al. Analysis of FLT3-Activating Mutations in 979 Patients with Acute Myelogenous Leukemia: Association with FAB Subtypes and Identification of Subgroups with Poor Prognosis. Blood 2002, 99, 4326–4335. [Google Scholar] [CrossRef] [PubMed]
- Slovak, M.L.; Gundacker, H.; Bloomfield, C.D.; Dewald, G.; Appelbaum, F.R.; Larson, R.A.; Tallman, M.S.; Bennett, J.M.; Stirewalt, D.L.; Meshinchi, S.; et al. A Retrospective Study of 69 Patients with t(6;9) (P23;Q34) AML Emphasizes the Need for a Prospective, Multicenter Initiative for Rare “poor Prognosis” Myeloid Malignancies. Leukemia 2006, 20, 1295–1297. [Google Scholar] [CrossRef]
- Larizza, L.; Magnani, I.; Beghini, A. The Kasumi-1 Cell Line: A t(8;21)-Kit Mutant Model for Acute Myeloid Leukemia. Leuk. Lymphoma 2005, 46, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Asou, H.; Gombart, A.F.; Takeuchi, S.; Tanaka, H.; Tanioka, M.; Matsui, H.; Kimura, A.; Inaba, T.; Koeffler, H.P. Establishment of the Acute Myeloid Leukemia Cell Line Kasumi-6 from a Patient with a Dominant-Negative Mutation in the DNA-Binding Region of the C/EBPalpha Gene. Genes Chromosomes Cancer 2003, 36, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Matozaki, S.; Nakagawa, T.; Kawaguchi, R.; Aozaki, R.; Tsutsumi, M.; Murayama, T.; Koizumi, T.; Nishimura, R.; Isobe, T.; Chihara, K. Establishment of a Myeloid Leukaemic Cell Line (SKNO-1) from a Patient with t(8;21) Who Acquired Monosomy 17 during Disease Progression. Br. J. Haematol. 1995, 89, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lv, N.; Zhou, L.; Li, Y.; Yu, L. Chidamide Inhibits t(8;21) AML Cell Proliferation and AMK1/ETO and C-KIT Expression by Inhibiting ERK1/2 Signaling Pathway. Transl. Cancer Res. 2020, 9, 827–839. [Google Scholar] [CrossRef]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; et al. Characterization of the Continuous, Differentiating Myeloid Cell Line (HL-60) from a Patient with Acute Promyelocytic Leukemia. Blood 1979, 54, 713–733. [Google Scholar] [CrossRef]
- Sak, K.; Everaus, H. Established Human Cell Lines as Models to Study Anti-Leukemic Effects of Flavonoids. Curr. Genom. 2017, 18, 3–26. [Google Scholar] [CrossRef]
- Kwa, F.A.A.; Cole-Sinclair, M.F.; Kapuscinski, M.K. Combination Treatment of P53-Null HL-60 Cells with Histone Deacetylase Inhibitors and Chlorambucil Augments Apoptosis and Increases BCL6 and P21 Gene Expression. Curr. Mol. Pharmacol. 2019, 12, 72–81. [Google Scholar] [CrossRef]
- Al-Otaibi, N.A.S.; Cassoli, J.S.; Slater, N.K.H.; Rahmoune, H. Molecular Characterization of Human Leukemia 60 (HL-60) Cells as a Model of Acute Myelogenous Leukemia Post Cryopreservation. Methods Mol. Biol. 2019, 1916, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Newburger, P.E.; Chovaniec, M.E.; Greenberger, J.S.; Cohen, H.J. Functional Changes in Human Leukemic Cell Line HL-60. A Model for Myeloid Differentiation. J. Cell Biol. 1979, 82, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.J. The HL-60 Promyelocytic Leukemia Cell Line: Proliferation, Differentiation, and Cellular Oncogene Expression. Blood 1987, 70, 1233–1244. [Google Scholar] [CrossRef]
- Johnston, J.J.; Rintels, P.; Chung, J.; Sather, J.; Benz, E.J.; Berliner, N. Lactoferrin Gene Promoter: Structural Integrity and Nonexpression in HL60 Cells. Blood 1992, 79, 2998–3006. [Google Scholar] [CrossRef]
- Jian, P.; Li, Z.W.; Fang, T.Y.; Jian, W.; Zhuan, Z.; Mei, L.X.; Yan, W.S.; Jian, N. Retinoic Acid Induces HL-60 Cell Differentiation via the Upregulation of MiR-663. J. Hematol. Oncol. 2011, 4, 20. [Google Scholar] [CrossRef]
- Tucker, K.A.; Lilly, M.B.; Heck, L.; Rado, T.A. Characterization of a New Human Diploid Myeloid Leukemia Cell Line (PLB-985) with Granulocytic and Monocytic Differentiating Capacity. Blood 1987, 70, 372–378. [Google Scholar] [CrossRef]
- Drexler, H.G.; Dirks, W.G.; Matsuo, Y.; MacLeod, R.a.F. False Leukemia–Lymphoma Cell Lines: An Update on over 500 Cell Lines. Leukemia 2003, 17, 416–426. [Google Scholar] [CrossRef]
- Rincón, E.; Rocha-Gregg, B.L.; Collins, S.R. A Map of Gene Expression in Neutrophil-like Cell Lines. BMC Genom. 2018, 19, 573. [Google Scholar] [CrossRef] [PubMed]
- Kamath, G.R.; Tremblay, D.; Coltoff, A.; Caro, J.; Lancman, G.; Bhalla, S.; Najfeld, V.; Mascarenhas, J.; Taioli, E. Comparing the Epidemiology, Clinical Characteristics and Prognostic Factors of Acute Myeloid Leukemia with and without Acute Promyelocytic Leukemia. Carcinogenesis 2019, 40, 651–660. [Google Scholar] [CrossRef]
- Hwang, S.M. Classification of Acute Myeloid Leukemia. Blood Res. 2020, 55 (Suppl. S1), S1–S4. [Google Scholar] [CrossRef]
- de Thé, H.; Lavau, C.; Marchio, A.; Chomienne, C.; Degos, L.; Dejean, A. The PML-RAR Alpha Fusion MRNA Generated by the t(15;17) Translocation in Acute Promyelocytic Leukemia Encodes a Functionally Altered RAR. Cell 1991, 66, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Lo-Coco, F.; Ammatuna, E. The Biology of Acute Promyelocytic Leukemia and Its Impact on Diagnosis and Treatment. Hematol. Am. Soc. Hematol. Educ. Program 2006, 156–161, 514. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, P.P. In Vivo Analysis of the Molecular Genetics of Acute Promyelocytic Leukemia. Oncogene 2001, 20, 5726–5735. [Google Scholar] [CrossRef] [PubMed]
- Khanna-Gupta, A.; Kolibaba, K.; Zibello, T.A.; Berliner, N. NB4 Cells Show Bilineage Potential and an Aberrant Pattern of Neutrophil Secondary Granule Protein Gene Expression. Blood 1994, 84, 294–302. [Google Scholar] [CrossRef]
- Duprez, E.; Ruchaud, S.; Houge, G.; Martin-Thouvenin, V.; Valensi, F.; Kastner, P.; Berger, R.; Lanotte, M. A Retinoid Acid “resistant” t(15;17) Acute Promyelocytic Leukemia Cell Line: Isolation, Morphological, Immunological, and Molecular Features. Leukemia 1992, 6, 1281–1287. [Google Scholar] [PubMed]
- Dermime, S.; Grignani, F.; Rogaia, D.; Liberatore, C.; Marchesi, E.; Gambacorti-Passerini, C. Acute Promyelocytic Leukaemia Cells Resistant to Retinoic Acid Show Further Perturbation of the RARα Signal Transduction System. Leuk. Lymphoma 1995, 16, 289–295. [Google Scholar] [CrossRef]
- Kubonishi, I.; Machida, K.; Sonobe, H.; Ohtsuki, Y.; Akagi, T.; Miyoshi, I. Two New Human Myeloid Cell Lines Derived from Acute Promyelocytic Leukemia and Chronic Myelocytic Leukemia. GANN Jpn. J. Cancer Res. 1983, 74, 319–322. [Google Scholar] [CrossRef]
- Sugimoto, K.; Toyoshima, H.; Sakai, R.; Miyagawa, K.; Hagiwara, K.; Ishikawa, F.; Takaku, F.; Yazaki, Y.; Hirai, H. Frequent Mutations in the P53 Gene in Human Myeloid Leukemia Cell Lines. Blood 1992, 79, 2378–2383. [Google Scholar] [CrossRef]
- Reiter, K.; Polzer, H.; Krupka, C.; Maiser, A.; Vick, B.; Rothenberg-Thurley, M.; Metzeler, K.H.; Dörfel, D.; Salih, H.R.; Jung, G.; et al. Tyrosine Kinase Inhibition Increases the Cell Surface Localization of FLT3-ITD and Enhances FLT3-Directed Immunotherapy of Acute Myeloid Leukemia. Leukemia 2018, 32, 313–322. [Google Scholar] [CrossRef]
- Kizaki, M.; Matsushita, H.; Takayama, N.; Muto, A.; Ueno, H.; Awaya, N.; Kawai, Y.; Asou, H.; Kamada, N.; Ikeda, Y. Establishment and Characterization of a Novel Acute Promyelocytic Leukemia Cell Line (UF-1) with Retinoic Acid-Resistant Features. Blood 1996, 88, 1824–1833. [Google Scholar] [CrossRef]
- Sato, A.; Imaizumi, M.; Hoshi, Y.; Rikiishi, T.; Fujii, K.; Kizaki, M.; Kagechika, H.; Kakizuka, A.; Hayashi, Y.; Iinuma, K. Alteration in the Cellular Response to Retinoic Acid of a Human Acute Promyelocytic Leukemia Cell Line, UF-1, Carrying a Patient-Derived Mutant PML-RARα Chimeric Gene. Leuk. Res. 2004, 28, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G. (Ed.) Front Matter. In The Leukemia-Lymphoma Cell Line FactsBook; Factsbook; Academic Press: London, UK, 2001; p. iii. [Google Scholar] [CrossRef]
- Takayama, N.; Kizaki, M.; Hida, T.; Kinjo, K.; Ikeda, Y. Novel Mutation in the PML/RARα Chimeric Gene Exhibits Dramatically Decreased Ligand-Binding Activity and Confers Acquired Resistance to Retinoic Acid in Acute Promyelocytic Leukemia. Exp. Hematol. 2001, 29, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T.; Kizaki, M.; Omine, M. Induction of Differentiation of Retinoic Acid-Resistant Acute Promyelocytic Leukemia Cells by the Combination of All-Trans Retinoic Acid and Granulocyte Colony-Stimulating Factor. Leuk. Res. 2004, 28, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Witcher, M.; Shiu, H.Y.; Guo, Q.; Miller, W.H. Combination of Retinoic Acid and Tumor Necrosis Factor Overcomes the Maturation Block in a Variety of Retinoic Acid-Resistant Acute Promyelocytic Leukemia Cells. Blood 2004, 104, 3335–3342. [Google Scholar] [CrossRef]
- Nakazato, T.; Ito, K.; Miyakawa, Y.; Kinjo, K.; Yamada, T.; Hozumi, N.; Ikeda, Y.; Kizaki, M. Catechin, a Green Tea Component, Rapidly Induces Apoptosis of Myeloid Leukemic Cells via Modulation of Reactive Oxygen Species Production in Vitro and Inhibits Tumor Growth in Vivo. Haematologica 2005, 90, 317–325. [Google Scholar]
- Komura, N.; Ikeda, Y.; Masuda, N.; Umezawa, Y.; Ito, K.; Kizaki, M.; Umezawa, K. Designed ATRA Analogue Active against ATRA-Resistant Acute Promyelocytic Leukemia Cells Having a Single Nucleotide Substitution in Their Retinoic Acid Receptor. Leuk. Res. 2007, 31, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Cassinat, B.; Zassadowski, F.; Ferry, C.; Llopis, L.; Bruck, N.; Lainey, E.; Duong, V.; Cras, A.; Despouy, G.; Chourbagi, O.; et al. New Role for Granulocyte Colony-Stimulating Factor-Induced Extracellular Signal-Regulated Kinase 1/2 in Histone Modification and Retinoic Acid Receptor α Recruitment to Gene Promoters: Relevance to Acute Promyelocytic Leukemia Cell Differentiation. Mol. Cell. Biol. 2011, 31, 1409–1418. [Google Scholar] [CrossRef]
- Iriyama, N.; Hatta, Y.; Takei, M. ETV6/ARG Oncoprotein Confers Autonomous Cell Growth by Enhancing c-Myc Expression via Signal Transducer and Activator of Transcription 5 Activation in the Acute Promyelocytic Leukemia Cell Line HT93A. Leuk. Lymphoma 2015, 56, 2416–2423. [Google Scholar] [CrossRef] [PubMed]
- Kishi, K.; Toba, K.; Azegami, T.; Tsukada, N.; Uesugi, Y.; Masuko, M.; Niwano, H.; Hashimoto, S.; Sakaue, M.; Furukawa, T.; et al. Hematopoietic Cytokine-Dependent Differentiation to Eosinophils and Neutrophils in a Newly Established Acute Promyelocytic Leukemia Cell Line with t(15;17). Exp. Hematol. 1998, 26, 135–142. [Google Scholar]
- Kitamura, K.; Kiyoi, H.; Yoshida, H.; Saito, H.; Ohno, R.; Naoe, T. Mutant AF-2 Domain of PML-RARalpha in Retinoic Acid-Resistant NB4 Cells: Differentiation Induced by RA Is Triggered Directly through PML-RARalpha and Its down-Regulation in Acute Promyelocytic Leukemia. Leukemia 1997, 11, 1950–1956. [Google Scholar] [CrossRef]
- Chen, S.; Nagel, S.; Schneider, B.; Dai, H.; Geffers, R.; Kaufman, M.; Meyer, C.; Pommerenke, C.; Thress, K.S.; Li, J.; et al. A New ETV6-NTRK3 Cell Line Model Reveals MALAT1 as a Novel Therapeutic Target—A Short Report. Cell. Oncol. 2018, 41, 93–101. [Google Scholar] [CrossRef]
- Corces-Zimmerman, M.R.; Hong, W.-J.; Weissman, I.L.; Medeiros, B.C.; Majeti, R. Preleukemic Mutations in Human Acute Myeloid Leukemia Affect Epigenetic Regulators and Persist in Remission. Proc. Natl. Acad. Sci. USA 2014, 111, 2548–2553. [Google Scholar] [CrossRef]
- Sano, H.; Shimada, A.; Taki, T.; Murata, C.; Park, M.; Sotomatsu, M.; Tabuchi, K.; Tawa, A.; Kobayashi, R.; Horibe, K.; et al. RAS Mutations Are Frequent in FAB Type M4 and M5 of Acute Myeloid Leukemia, and Related to Late Relapse: A Study of the Japanese Childhood AML Cooperative Study Group. Int. J. Hematol. 2012, 95, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Tiacci, E.; Spanhol-Rosseto, A.; Martelli, M.P.; Pasqualucci, L.; Quentmeier, H.; Grossmann, V.; Drexler, H.G.; Falini, B. The NPM1 Wild-Type OCI-AML2 and the NPM1-Mutated OCI-AML3 Cell Lines Carry DNMT3A Mutations. Leukemia 2012, 26, 554–557. [Google Scholar] [CrossRef]
- Falini, B.; Brunetti, L.; Sportoletti, P.; Martelli, M.P. NPM1-Mutated Acute Myeloid Leukemia: From Bench to Bedside. Blood 2020, 136, 1707–1721. [Google Scholar] [CrossRef] [PubMed]
- Banella, C.; Catalano, G.; Travaglini, S.; Pelosi, E.; Ottone, T.; Zaza, A.; Guerrera, G.; Angelini, D.F.; Niscola, P.; Divona, M.; et al. Ascorbate Plus Buformin in AML: A Metabolic Targeted Treatment. Cancers 2022, 14, 2565. [Google Scholar] [CrossRef] [PubMed]
- Grenier, A.; Poulain, L.; Mondesir, J.; Jacquel, A.; Bosc, C.; Stuani, L.; Mouche, S.; Larrue, C.; Sahal, A.; Birsen, R.; et al. AMPK-PERK Axis Represses Oxidative Metabolism and Enhances Apoptotic Priming of Mitochondria in Acute Myeloid Leukemia. Cell Rep. 2022, 38, 110197. [Google Scholar] [CrossRef]
- Zheng, A.; Castren, K.; Säily, M.; Savolainen, E.-R.; Koistinen, P.; Vähäkangas, K. P53 Status of Newly Established Acute Myeloid Leukaemia Cell Lines. Br. J. Cancer 1999, 79, 407–415. [Google Scholar] [CrossRef]
- Hu, Z.-B.; Quentmeier, H.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. New Cytokine-Dependent Acute Myeloid Leukemia Cell Line MUTZ-11 with Disomic Chromosome Rearrangement t(16;17). Leuk. Res. 2004, 28, 509–515. [Google Scholar] [CrossRef] [PubMed]
- El Fitori, J.; Su, Y.; Büchler, P.; Ludwig, R.; Giese, N.A.; Büchler, M.W.; Quentmeier, H.; Hines, O.J.; Herr, I.; Friess, H. PKC 412 Small-Molecule Tyrosine Kinase Inhibitor: Single-Compound Therapy for Pancreatic Cancer. Cancer 2007, 110, 1457–1468. [Google Scholar] [CrossRef]
- Liu, W.; Deng, L.; Song, Y.; Redell, M. DOT1L Inhibition Sensitizes MLL-Rearranged AML to Chemotherapy. PLoS ONE 2014, 9, e98270. [Google Scholar] [CrossRef] [PubMed]
- Kühn, M.W.M.; Hadler, M.J.; Daigle, S.R.; Koche, R.P.; Krivtsov, A.V.; Olhava, E.J.; Caligiuri, M.A.; Huang, G.; Bradner, J.E.; Pollock, R.M.; et al. MLL Partial Tandem Duplication Leukemia Cells Are Sensitive to Small Molecule DOT1L Inhibition. Haematologica 2015, 100, e190–e193. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-B.; MacLeod, R.a.F.; Meyer, C.; Quentmeier, H.; Drexler, H.G. New Acute Myeloid Leukemia-Derived Cell Line: MUTZ-8 with 5q-. Leukemia 2002, 16, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-B.; Quentmeier, H.; Meyer, C.; MacLeod, R.A.F.; Drexler, H.G. Establishment of the Two Cytokine-Dependent Acute Myeloid Leukemia-Derived Cell Lines MUTZ-8 and MUTZ-11. In Acute Leukemias IX; Hiddemann, W., Haferlach, T., Unterhalt, M., Büchner, T., Ritter, J., Eds.; Haematology and Blood Transfusion Hämatologie und Bluttransfusion; Springer: Berlin/Heidelberg, Germany, 2003; pp. 122–127. [Google Scholar] [CrossRef]
- Quentmeier, H.; MacLeod, R.a.F.; Zaborski, M.; Drexler, H.G. JAK2 V617F Tyrosine Kinase Mutation in Cell Lines Derived from Myeloproliferative Disorders. Leukemia 2006, 20, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Geffers, R.; Jost, E.; MacLeod, R.a.F.; Nagel, S.; Röhrs, S.; Romani, J.; Scherr, M.; Zaborski, M.; Drexler, H.G. SOCS2: Inhibitor of JAK2V617F-Mediated Signal Transduction. Leukemia 2008, 22, 2169–2175. [Google Scholar] [CrossRef]
- Hu, Z.B.; Ma, W.; Zaborski, M.; MacLeod, R.; Quentmeier, H.; Drexler, H.G. Establishment and Characterization of Two Novel Cytokine-Responsive Acute Myeloid and Monocytic Leukemia Cell Lines, MUTZ-2 and MUTZ-3. Leukemia 1996, 10, 1025–1040. [Google Scholar]
- Masterson, A.J.; Sombroek, C.C.; De Gruijl, T.D.; Graus, Y.M.F.; van der Vliet, H.J.J.; Lougheed, S.M.; van den Eertwegh, A.J.M.; Pinedo, H.M.; Scheper, R.J. MUTZ-3, a Human Cell Line Model for the Cytokine-Induced Differentiation of Dendritic Cells from CD34+ Precursors. Blood 2002, 100, 701–703. [Google Scholar] [CrossRef]
- Santegoets, S.J.A.M.; Schreurs, M.W.J.; Masterson, A.J.; Liu, Y.P.; Goletz, S.; Baumeister, H.; Kueter, E.W.M.; Lougheed, S.M.; van den Eertwegh, A.J.M.; Scheper, R.J.; et al. In Vitro Priming of Tumor-Specific Cytotoxic T Lymphocytes Using Allogeneic Dendritic Cells Derived from the Human MUTZ-3 Cell Line. Cancer Immunol. Immunother. 2006, 55, 1480–1490. [Google Scholar] [CrossRef]
- Santegoets, S.J.A.M.; van den Eertwegh, A.J.M.; van de Loosdrecht, A.A.; Scheper, R.J.; de Gruijl, T.D. Human Dendritic Cell Line Models for DC Differentiation and Clinical DC Vaccination Studies. J. Leukoc. Biol. 2008, 84, 1364–1373. [Google Scholar] [CrossRef]
- Quentmeier, H.; Drexler, H.G.; Fleckenstein, D.; Zaborski, M.; Armstrong, A.; Sims, J.E.; Lyman, S.D. Cloning of Human Thymic Stromal Lymphopoietin (TSLP) and Signaling Mechanisms Leading to Proliferation. Leukemia 2001, 15, 1286–1292. [Google Scholar] [CrossRef] [PubMed]
- Rasaiyaah, J.; Noursadeghi, M.; Kellam, P.; Chain, B. Transcriptional and Functional Defects of Dendritic Cells Derived from the MUTZ-3 Leukaemia Line. Immunology 2009, 127, 429–441. [Google Scholar] [CrossRef]
- Gimeno, M.; San José-Enériz, E.; Villar, S.; Agirre, X.; Prosper, F.; Rubio, A.; Carazo, F. Explainable Artificial Intelligence for Precision Medicine in Acute Myeloid Leukemia. Front. Immunol. 2022, 13, 977358. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami, O.; Friedman, D.; Leshkowitz, D.; Goldenberg, D.; Orlovsky, K.; Pencovich, N.; Lotem, J.; Tanay, A.; Groner, Y. Addiction of t(8;21) and Inv(16) Acute Myeloid Leukemia to Native RUNX1. Cell Rep. 2013, 4, 1131–1143. [Google Scholar] [CrossRef]
- Wilkinson, A.C.; Ballabio, E.; Geng, H.; North, P.; Tapia, M.; Kerry, J.; Biswas, D.; Roeder, R.G.; Allis, C.D.; Melnick, A.; et al. RUNX1 Is a Key Target in t(4;11) Leukemias That Contributes to Gene Activation through an AF4-MLL Complex Interaction. Cell Rep 2013, 3, 116–127. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Y.; Williams, J.; Hang, Y.; Richter, L.; Becker, M.; Amador, C.; Oupický, D.; Hyde, R.K. Use of Polymeric CXCR4 Inhibitors as SiRNA Delivery Vehicles for the Treatment of Acute Myeloid Leukemia. Cancer Gene Ther. 2019, 27, 45–55. [Google Scholar] [CrossRef]
- Gilby, D.C.; Sung, H.Y.; Winship, P.R.; Goodeve, A.C.; Reilly, J.T.; Kiss-Toth, E. Tribbles-1 and -2 Are Tumour Suppressors, down-Regulated in Human Acute Myeloid Leukaemia. Immunol. Lett. 2010, 130, 115–124. [Google Scholar] [CrossRef]
- Cripe, L.D.; Gelfanov, V.M.; Smith, E.A.; Spigel, D.R.; Phillips, C.A.; Gabig, T.G.; Jung, S.-H.; Fyffe, J.; Hartman, A.D.; Kneebone, P.; et al. Role for C-Jun N-Terminal Kinase in Treatment-Refractory Acute Myeloid Leukemia (AML): Signaling to Multidrug-Efflux and Hyperproliferation. Leukemia 2002, 16, 799–812. [Google Scholar] [CrossRef]
- Tallman, M.S.; Kim, H.T.; Paietta, E.; Bennett, J.M.; Dewald, G.; Cassileth, P.A.; Wiernik, P.H.; Rowe, J.M. Acute Monocytic Leukemia (French-American-British Classification M5) Does Not Have a Worse Prognosis Than Other Subtypes of Acute Myeloid Leukemia: A Report From the Eastern Cooperative Oncology Group. JCO 2004, 22, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and Characterization of a Human Acute Monocytic Leukemia Cell Line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Humeniuk-Polaczek, R.; Marcinkowska, E. Impaired Nuclear Localization of Vitamin D Receptor in Leukemia Cells Resistant to Calcitriol-Induced Differentiation. J. Steroid Biochem. Mol. Biol. 2004, 88, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.; Cotter, P.; López-Expósito, I.; Kleiveland, C.; Lea, T.; Mackie, A.; Requena, T.; Swiatecka, D.; Wichers, H. (Eds.) The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Schnoor, M.; Buers, I.; Sietmann, A.; Brodde, M.F.; Hofnagel, O.; Robenek, H.; Lorkowski, S. Efficient Non-Viral Transfection of THP-1 Cells. J. Immunol. Methods 2009, 344, 109–115. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Savelkoul, H.F.J.; Wichers, H.J. Transcription Profiles of LPS-Stimulated THP-1 Monocytes and Macrophages: A Tool to Study Inflammation Modulating Effects of Food-Derived Compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Ashikaga, T.; Yoshida, Y.; Hirota, M.; Yoneyama, K.; Itagaki, H.; Sakaguchi, H.; Miyazawa, M.; Ito, Y.; Suzuki, H.; Toyoda, H. Development of an in Vitro Skin Sensitization Test Using Human Cell Lines: The Human Cell Line Activation Test (h-CLAT): I. Optimization of the h-CLAT Protocol. Toxicol. In Vitro 2006, 20, 767–773. [Google Scholar] [CrossRef]
- Sakaguchi, H.; Ashikaga, T.; Miyazawa, M.; Yoshida, Y.; Ito, Y.; Yoneyama, K.; Hirota, M.; Itagaki, H.; Toyoda, H.; Suzuki, H. Development of an in Vitro Skin Sensitization Test Using Human Cell Lines; Human Cell Line Activation Test (h-CLAT) II. An Inter-Laboratory Study of the h-CLAT. Toxicol. Vitr. 2006, 20, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Helal, R.; Melzig, M.F. New Aspects in the Synthesis and Secretion of Lysozyme by Cultured Human Monocyte Cell Lines. Vitr. Cell. Dev. Biol. Anim. 2010, 46, 492–496. [Google Scholar] [CrossRef]
- Helal, R.; Melzig, M.F. In Vitro Effects of Selected Saponins on the Production and Release of Lysozyme Activity of Human Monocytic and Epithelial Cell Lines. Sci. Pharm. 2011, 79, 337–350. [Google Scholar] [CrossRef]
- Helal, R.; Bader, G.; Melzig, M.F. Stimulation of Lysozyme Release by Selected Microbial Preparations. Die Pharm.-Int. J. Pharm. Sci. 2012, 67, 564–566. [Google Scholar] [CrossRef]
- Pandur, E.; Micalizzi, G.; Mondello, L.; Horváth, A.; Sipos, K.; Horváth, G. Antioxidant and Anti-Inflammatory Effects of Thyme (Thymus vulgaris L.) Essential Oils Prepared at Different Plant Phenophases on Pseudomonas Aeruginosa LPS-Activated THP-1 Macrophages. Antioxidants 2022, 11, 1330. [Google Scholar] [CrossRef] [PubMed]
- Bosshart, H.; Heinzelmann, M. Lipopolysaccharide-Mediated Cell Activation without Rapid Mobilization of Cytosolic Free Calcium. Mol. Immunol. 2004, 41, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Bruckmeier, M.; Kuehnl, A.; Culmes, M.; Pelisek, J.; Eckstein, H.-H. Impact of OxLDL and LPS on C-Type Natriuretic Peptide System Is Different between THP-1 Cells and Human Peripheral Blood Monocytic Cells. CPB 2012, 30, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Bosshart, H.; Heinzelmann, M. Arginine-Rich Cationic Polypeptides Amplify Lipopolysaccharide-Induced Monocyte Activation. Infect. Immun. 2002, 70, 6904–6910. [Google Scholar] [CrossRef]
- Bosshart, H.; Heinzelmann, M. THP-1 Cells as a Model for Human Monocytes. Ann. Transl. Med. 2016, 4, 438. [Google Scholar] [CrossRef]
- Sundström, C.; Nilsson, K. Establishment and Characterization of a Human Histiocytic Lymphoma Cell Line (U-937). Int. J. Cancer 1976, 17, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Aggerholm, A.; Grønbaek, K.; Guldberg, P.; Hokland, P. Mutational Analysis of the Tumour Suppressor Gene MMAC1/PTEN in Malignant Myeloid Disorders. Eur. J. Haematol. 2000, 65, 109–113. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. ClinVar, VCV000449416.13. Available online: https://www.ncbi.nlm.nih.gov/clinvar/variation/VCV000449416.13 (accessed on 29 December 2022).
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 Cell Line: An in Vitro Cell Model for Immune Modulation Approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, K.; Tauber, S.; Goelz, N.; Simmet, D.M.; Engeli, S.; Birlem, M.; Dumrese, C.; Karer, A.; Hunziker, S.; Biskup, J.; et al. Severe Disruption of the Cytoskeleton and Immunologically Relevant Surface Molecules in a Human Macrophageal Cell Line in Microgravity—Results of an in Vitro Experiment on Board of the Shenzhou-8 Space Mission. Acta Astronaut. 2014, 94, 277–292. [Google Scholar] [CrossRef]
- Olsson, I.L.; Breitman, T.R. Induction of Differentiation of the Human Histiocytic Lymphoma Cell Line U-937 by Retinoic Acid and Cyclic Adenosine 3’:5’-Monophosphate-Inducing Agents. Cancer Res. 1982, 42, 3924–3927. [Google Scholar]
- Hattori, T.; Pack, M.; Bougnoux, P.; Chang, Z.L.; Hoffman, T. Interferon-Induced Differentiation of U937 Cells. Comparison with Other Agents That Promote Differentiation of Human Myeloid or Monocytelike Cell Lines. J. Clin. Investig. 1983, 72, 237–244. [Google Scholar] [CrossRef]
- Olsson, I.; Gullberg, U.; Ivhed, I.; Nilsson, K. Induction of Differentiation of the Human Histiocytic Lymphoma Cell Line U-937 by 1 Alpha,25-Dihydroxycholecalciferol. Cancer Res. 1983, 43 Pt 1, 5862–5867. [Google Scholar] [PubMed]
- Stöckbauer, P.; Malasková, V.; Soucek, J.; Chudomel, V. Differentiation of Human Myeloid Leukemia Cell Lines Induced by Tumor-Promoting Phorbol Ester (TPA). I. Changes of the Morphology, Cytochemistry and the Surface Differentiation Antigens Analyzed with Monoclonal Antibodies. Neoplasma 1983, 30, 257–272. [Google Scholar]
- Minafra, L.; Di Cara, G.; Albanese, N.N.; Cancemi, P. Proteomic Differentiation Pattern in the U937 Cell Line. Leuk. Res. 2011, 35, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.R.; Um, H.-D. Hydrogen Peroxide Suppresses U937 Cell Death by Two Different Mechanisms Depending on Its Concentration. Exp. Cell Res. 1999, 248, 430–438. [Google Scholar] [CrossRef]
- Kim, D.K.; Cho, E.S.; Lee, B.R.; Um, H.-D. NF-ΚB Mediates the Adaptation of Human U937 Cells to Hydrogen Peroxide. Free. Radic. Biol. Med. 2001, 30, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Cho, E.S.; Yang, H.S.; Kim, H.; Um, H.-D. Serum Withdrawal Kills U937 Cells by Inducing a Positive Mutual Interaction between Reactive Oxygen Species and Phosphoinositide 3-Kinase. Cell. Signal. 2005, 17, 197–204. [Google Scholar] [CrossRef]
- Song, M.; Ryoo, I.; Choi, H.; Choi, B.; Kim, S.-T.; Heo, T.-H.; Lee, J.Y.; Park, P.-H.; Kwak, M.-K. NRF2 Signaling Negatively Regulates Phorbol-12-Myristate-13-Acetate (PMA)-Induced Differentiation of Human Monocytic U937 Cells into Pro-Inflammatory Macrophages. PLoS ONE 2015, 10, e0134235. [Google Scholar] [CrossRef]
- Vu, M.; Kassouf, N.; Appiah, S. Betulinic Acid–Doxorubicin-Drug Combination Induced Apoptotic Death via ROS Stimulation in a Relapsed AML MOLM-13 Cell Model. Antioxidants 2021, 10, 1456. [Google Scholar] [CrossRef] [PubMed]
- Greco, G.; Schnekenburger, M.; Catanzaro, E.; Turrini, E.; Ferrini, F.; Sestili, P.; Diederich, M.; Fimognari, C. Discovery of Sulforaphane as an Inducer of Ferroptosis in U-937 Leukemia Cells: Expanding Its Anticancer Potential. Cancers 2022, 14, 76. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Balukova, A.; Ovsii, A.; Rác, M.; Křupka, M.; Kasai, S.; Pospíšil, P. Reactive Oxygen Species Imaging in U937 Cells. Front. Physiol. 2020, 11, 552569. [Google Scholar] [CrossRef]
- Nathan, I.; Dizdaroglu, M.; Bernstein, L.; Junker, U.; Lee, C.-K.; Muegge, K.; Durum, S.K. Induction of oxidative DNA damage in U937 cells by TNF or anti-Fas stimulation. Cytokine 2000, 12, 881–887. [Google Scholar] [CrossRef]
- Pina-Jiménez, E.; Calzada, F.; Bautista, E.; Ordoñez-Razo, R.M.; Velázquez, C.; Barbosa, E.; García-Hernández, N. Incomptine A Induces Apoptosis, ROS Production and a Differential Protein Expression on Non-Hodgkin’s Lymphoma Cells. Int. J. Mol. Sci. 2021, 22, 10516. [Google Scholar] [CrossRef]
- del Rosario, H.; Saavedra, E.; Brouard, I.; González-Santana, D.; García, C.; Spínola-Lasso, E.; Tabraue, C.; Quintana, J.; Estévez, F. Structure-Activity Relationships Reveal a 2-Furoyloxychalcone as a Potent Cytotoxic and Apoptosis Inducer for Human U-937 and HL-60 Leukaemia Cells. Bioorg. Chem. 2022, 127, 105926. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-S.; Jeong, S.-I.; Hwang, B.-S.; Lee, Y.-E.; Kang, S.-H.; Lee, H.-C.; Oh, C.-H. Gallic Acid Inhibits Cell Viability and Induces Apoptosis in Human Monocytic Cell Line U937. J. Med. Food 2011, 14, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Endo, M.; Matsui, T.; Katsuda, I.; Emi, N.; Kawamoto, Y.; Koike, T.; Beppu, H. Agaritine from Agaricus Blazei Murrill Induces Apoptosis in the Leukemic Cell Line U937. Biochim. Biophys. Acta-Gen. Subj. 2011, 1810, 519–525. [Google Scholar] [CrossRef]
- Kaba, S.I.; Egorova, E.M. In Vitro Studies of the Toxic Effects of Silver Nanoparticles on HeLa and U937 Cells. Nanotechnol. Sci. Appl. 2015, 8, 19–29. [Google Scholar] [CrossRef]
- Barbasz, A.; Kreczmer, B.; Skórka, M.; Czyżowska, A. Toxicity of Pesticides toward Human Immune Cells U-937 and HL-60. J. Environ. Sci. Health B 2020, 55, 719–725. [Google Scholar] [CrossRef]
- OECD. U-SENSTM—Myeloid U937 Skin Sensitisation Test; OECD: Paris, France, 2013; Available online: https://tsar.jrc.ec.europa.eu/test-method/tm2013-02 (accessed on 29 December 2022).
- Passmore, J.S.; Lukey, P.T.; Ress, S.R. The Human Macrophage Cell Line U937 as an in Vitro Model for Selective Evaluation of Mycobacterial Antigen-Specific Cytotoxic T-Cell Function. Immunology 2001, 102, 146–156. [Google Scholar] [CrossRef]
- Kaszubowska, L.; Engelmann, H.; Gotartowska, M.; Iliszko, M.; Bigda, J. Identification of two U937 cell sublines exhibiting different patterns of response to tumour necrosis factor. Cytokine 2001, 13, 365–370. [Google Scholar] [CrossRef]
- Stasiłojć, G.; Pinto, S.; Wyszkowska, R.; Wejda, M.; Słomińska, E.M.; Filipska, M.; Koszałka, P.; Swierczyński, J.; O’Connor, J.E.; Bigda, J.J. U937 Variant Cells as a Model of Apoptosis without Cell Disintegration. Cell. Mol. Biol. Lett. 2013, 18, 249–262. [Google Scholar] [CrossRef]
- Reid, Y.A.; McGuire, L.; O’Neill, K.; Macy, M.; Chen, T.R.; McClintock, P.; Dorotinsky, C.; Hay, R. Cell Line Cross-Contamination of U-937 [Correction of U-397]. J. Leukoc. Biol. 1995, 57, 804. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; MacLeod, R.A.; Uphoff, C.C.; Drexler, H.G.; Nishizaki, C.; Katayama, Y.; Kimura, G.; Fujii, N.; Omoto, E.; Harada, M.; et al. Two Acute Monocytic Leukemia (AML-M5a) Cell Lines (MOLM-13 and MOLM-14) with Interclonal Phenotypic Heterogeneity Showing MLL-AF9 Fusion Resulting from an Occult Chromosome Insertion, Ins(11;9) (Q23;P22p23). Leukemia 1997, 11, 1469–1477. [Google Scholar] [CrossRef]
- Lange, B.; Valtieri, M.; Santoli, D.; Caracciolo, D.; Mavilio, F.; Gemperlein, I.; Griffin, C.; Emanuel, B.; Finan, J.; Nowell, P. Growth Factor Requirements of Childhood Acute Leukemia: Establishment of GM-CSF-Dependent Cell Lines. Blood 1987, 70, 192–199. [Google Scholar] [CrossRef]
- Stong, R.C.; Kersey, J.H. In Vitro Culture of Leukemic Cells in t(4;11) Acute Leukemia. Blood 1985, 66, 439–443. [Google Scholar] [CrossRef]
- Parkin, J.L.; Arthur, D.C.; Abramson, C.S.; McKenna, R.W.; Kersey, J.H.; Heideman, R.L.; Brunning, R.D. Acute Leukemia Associated with the t(4;11) Chromosome Rearrangement: Ultrastructural and Immunologic Characteristics. Blood 1982, 60, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Chen, Q.; Xu, J.; Li, W.; Xu, B.; Qiu, Y. Low-Frequency TP53 Hotspot Mutation Contributes to Chemoresistance through Clonal Expansion in Acute Myeloid Leukemia. Leukemia 2020, 34, 1816–1827. [Google Scholar] [CrossRef]
- Long, J.; Jia, M.-Y.; Fang, W.-Y.; Chen, X.-J.; Mu, L.-L.; Wang, Z.-Y.; Shen, Y.; Xiang, R.-F.; Wang, L.-N.; Wang, L.; et al. FLT3 Inhibition Upregulates HDAC8 via FOXO to Inactivate P53 and Promote Maintenance of FLT3-ITD+ Acute Myeloid Leukemia. Blood 2020, 135, 1472–1483. [Google Scholar] [CrossRef] [PubMed]
- Kojima, K.; McQueen, T.; Chen, Y.; Jacamo, R.; Konopleva, M.; Shinojima, N.; Shpall, E.; Huang, X.; Andreeff, M. P53 Activation of Mesenchymal Stromal Cells Partially Abrogates Microenvironment-Mediated Resistance to FLT3 Inhibition in AML through HIF-1α–Mediated down-Regulation of CXCL12. Blood 2011, 118, 4431–4439. [Google Scholar] [CrossRef] [PubMed]
- Medina-Medina, I.; Martínez-Sánchez, M.; Hernández-Monge, J.; Fahraeus, R.; Muller, P.; Olivares-Illana, V. P53 Promotes Its Own Polyubiquitination by Enhancing the HDM2 and HDMX Interaction. Protein Sci. 2018, 27, 976–986. [Google Scholar] [CrossRef]
- Zimmerman, E.I.; Turner, D.C.; Buaboonnam, J.; Hu, S.; Orwick, S.; Roberts, M.S.; Janke, L.J.; Ramachandran, A.; Stewart, C.F.; Inaba, H.; et al. Crenolanib Is Active against Models of Drug-Resistant FLT3-ITD−positive Acute Myeloid Leukemia. Blood 2013, 122, 3607–3615. [Google Scholar] [CrossRef]
- Lu, J.-W.; Wang, A.-N.; Liao, H.-A.; Chen, C.-Y.; Hou, H.-A.; Hu, C.-Y.; Tien, H.-F.; Ou, D.-L.; Lin, L.-I. Cabozantinib Is Selectively Cytotoxic in Acute Myeloid Leukemia Cells with FLT3-Internal Tandem Duplication (FLT3-ITD). Cancer Lett. 2016, 376, 218–225. [Google Scholar] [CrossRef]
- Capelli, D.; Menotti, D.; Fiorentini, A.; Saraceni, F.; Olivieri, A. Overcoming Resistance: FLT3 Inhibitors Past, Present, Future and the Challenge of Cure. Cancers 2022, 14, 4315. [Google Scholar] [CrossRef]
- Yu, Z.; Du, J.; Hui, H.; Kan, S.; Huo, T.; Zhao, K.; Wu, T.; Guo, Q.; Lu, N. LT-171-861, a Novel FLT3 Inhibitor, Shows Excellent Preclinical Efficacy for the Treatment of FLT3 Mutant Acute Myeloid Leukemia. Theranostics 2021, 11, 93–106. [Google Scholar] [CrossRef]
- Hart, S.; Goh, K.C.; Novotny-Diermayr, V.; Tan, Y.C.; Madan, B.; Amalini, C.; Ong, L.C.; Kheng, B.; Cheong, A.; Zhou, J.; et al. Pacritinib (SB1518), a JAK2/FLT3 Inhibitor for the Treatment of Acute Myeloid Leukemia. Blood Cancer J. 2011, 1, e44. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Kaneko, N.; Ueno, Y.; Yamada, M.; Tanaka, R.; Saito, R.; Shimada, I.; Mori, K.; Kuromitsu, S. Gilteritinib, a FLT3/AXL Inhibitor, Shows Antileukemic Activity in Mouse Models of FLT3 Mutated Acute Myeloid Leukemia. Investig. New Drugs 2017, 35, 556–565. [Google Scholar] [CrossRef]
- Cervera, N.; Carbuccia, N.; Garnier, S.; Guille, A.; Adélaïde, J.; Murati, A.; Vey, N.; Mozziconacci, M.-J.; Chaffanet, M.; Birnbaum, D.; et al. Molecular Characterization of Acute Erythroid Leukemia (M6-AML) Using Targeted next-Generation Sequencing. Leukemia 2016, 30, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Naeim, F.; Nagesh Rao, P.; Song, S.X.; Grody, W.W. 21—Acute Myeloid Leukemia, Not Otherwise Specified. In Atlas of Hematopathology; Naeim, F., Nagesh Rao, P., Song, S.X., Grody, W.W., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 259–281. [Google Scholar] [CrossRef]
- Martin, P.; Papayannopoulou, T. HEL Cells: A New Human Erythroleukemia Cell Line with Spontaneous and Induced Globin Expression. Science 1982, 216, 1233–1235. [Google Scholar] [CrossRef] [PubMed]
- Petiti, J.; Rosso, V.; Lo Iacono, M.; Panuzzo, C.; Calabrese, C.; Signorino, E.; Pironi, L.; Cartellà, A.; Bracco, E.; Pergolizzi, B.; et al. Curcumin Induces Apoptosis in JAK2-mutated Cells by the Inhibition of JAK2/STAT and MTORC1 Pathways. J. Cell. Mol. Med. 2019, 23, 4349–4357. [Google Scholar] [CrossRef] [PubMed]
- Boddu, P.; Benton, C.B.; Wang, W.; Borthakur, G.; Khoury, J.D.; Pemmaraju, N. Erythroleukemia-Historical Perspectives and Recent Advances in Diagnosis and Management. Blood Reviews 2018, 32, 96–105. [Google Scholar] [CrossRef]
- Santos, F.P.S.; Verstovsek, S. JAK2 Inhibitors: Are They the Solution? Clin. Lymphoma Myeloma Leuk. 2011, 11, S28–S36. [Google Scholar] [CrossRef]
- Venugopal, S.; Bar-Natan, M.; Mascarenhas, J.O. JAKs to STATs: A Tantalizing Therapeutic Target in Acute Myeloid Leukemia. Blood Rev. 2020, 40, 100634. [Google Scholar] [CrossRef]
- Broudy, V.C.; Lin, N.; Egrie, J.; de Haën, C.; Weiss, T.; Papayannopoulou, T.; Adamson, J.W. Identification of the Receptor for Erythropoietin on Human and Murine Erythroleukemia Cells and Modulation by Phorbol Ester and Dimethyl Sulfoxide. Proc. Natl. Acad. Sci. USA 1988, 85, 6513–6517. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Pommerenke, C.; Meyer, C.; MacLeod, R.A.F. NKL Homeobox Genes NKX2-3 and NKX2-4 Deregulate Megakaryocytic-Erythroid Cell Differentiation in AML. Int. J. Mol. Sci. 2021, 22, 11434. [Google Scholar] [CrossRef] [PubMed]
- Estrov, Z.; Shishodia, S.; Faderl, S.; Harris, D.; Van, Q.; Kantarjian, H.M.; Talpaz, M.; Aggarwal, B.B. Resveratrol Blocks Interleukin-1beta-Induced Activation of the Nuclear Transcription Factor NF-KappaB, Inhibits Proliferation, Causes S-Phase Arrest, and Induces Apoptosis of Acute Myeloid Leukemia Cells. Blood 2003, 102, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulou, T.; Nakamoto, B.; Kurachi, S.; Tweeddale, M.; Messner, H. Surface Antigenic Profile and Globin Phenotype of Two New Human Erythroleukemia Lines: Characterization and Interpretations. Blood 1988, 72, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Takaku, F.; Tange, T.; Shibuya, K.; Misawa, C.; Sasaki, K.; Miyagawa, K.; Yazaki, Y.; Hirai, H. Establishment and Erythroid Differentiation of a Cytokine-Dependent Human Leukemic Cell Line F-36: A Parental Line Requiring Granulocyte- Macrophage Colony-Stimulating Factor or Interleukin-3, and a Subline Requiring Erythropoietin. Blood 1991, 78, 2261–2268. [Google Scholar] [CrossRef]
- Drexler, H.G.; Zaborski, M.; Quentmeier, H. Cytokine Response Profiles of Human Myeloid Factor-Dependent Leukemia Cell Lines. Leukemia 1997, 11, 701–708. [Google Scholar] [CrossRef]
- Kitamura, T.; Tojo, A.; Kuwaki, T.; Chiba, S.; Miyazono, K.; Urabe, A.; Takaku, F. Identification and Analysis of Human Erythropoietin Receptors on a Factor-Dependent Cell Line, TF-1. Blood 1989, 73, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, T.; Tange, T.; Terasawa, T.; Chiba, S.; Kuwaki, T.; Miyagawa, K.; Piao, Y.-F.; Miyazono, K.; Urabe, A.; Takaku, F. Establishment and Characterization of a Unique Human Cell Line That Proliferates Dependently on GM-CSF, IL-3, or Erythropoietin. J. Cell. Physiol. 1989, 140, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Chrétien, S.; Varlet, P.; Verdier, F.; Gobert, S.; Cartron, J.P.; Gisselbrecht, S.; Mayeux, P.; Lacombe, C. Erythropoietin-Induced Erythroid Differentiation of the Human Erythroleukemia Cell Line TF-1 Correlates with Impaired STAT5 Activation. EMBO J. 1996, 15, 4174–4181. [Google Scholar] [CrossRef] [PubMed]
- Steube, K.G.; Meyer, C.; Tachibana, M.; Murai, M.; Drexler, H.G. Bladder Carcinoma Cell Line KU-19-19-Derived Cytokines Support Proliferation of Growth Factor-Dependent Hematopoietic Cell Lines: Modulation by Phorbol Ester, Interferon-γ and Interleukin-1β. Biochem. Biophys. Res. Commun. 1998, 242, 497–501. [Google Scholar] [CrossRef]
- Tucker, S.J.; Rae, C.; Littlejohn, A.F.; Paul, A.; MacEwan, D.J. Switching Leukemia Cell Phenotype between Life and Death. Proc. Natl. Acad. Sci. USA 2004, 101, 12940–12945. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Kuriyama, K.; Higuchi, M.; Tsushima, H.; Sohda, H.; Imai, N.; Saito, M.; Kondo, T.; Tomonaga, M. Establishment and Characterization of a New Erythropoietin-Dependent Acute Myeloid Leukemia Cell Line, AS-E2. Leukemia 1997, 11, 1941–1949. [Google Scholar] [CrossRef]
- Vicente, C.; Conchillo, A.; García-Sánchez, M.A.; Odero, M.D. The Role of the GATA2 Transcription Factor in Normal and Malignant Hematopoiesis. Crit. Rev. Oncol./Hematol. 2012, 82, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wu, W.; Wang, X.; Gu, J. Clinical Diagnosis of Adult Patients with Acute Megakaryocytic Leukemia. Oncol. Lett. 2018, 16, 6988–6997. [Google Scholar] [CrossRef]
- De Marchi, F.; Araki, M.; Komatsu, N. Molecular Features, Prognosis, and Novel Treatment Options for Pediatric Acute Megakaryoblastic Leukemia. Expert Rev. Hematol. 2019, 12, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.W.; Li, B.; Prouet, P.; Giri, S.; Pathak, R.; Martin, M.G. Acute Megakaryocytic Leukemia: What Have We Learned. Blood Rev. 2016, 30, 49–53. [Google Scholar] [CrossRef]
- Yoshida, K.; Toki, T.; Okuno, Y.; Kanezaki, R.; Shiraishi, Y.; Sato-Otsubo, A.; Sanada, M.; Park, M.; Terui, K.; Suzuki, H.; et al. The Landscape of Somatic Mutations in Down Syndrome–Related Myeloid Disorders. Nat. Genet. 2013, 45, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Hitzler, J.K.; Cheung, J.; Li, Y.; Scherer, S.W.; Zipursky, A. GATA1 Mutations in Transient Leukemia and Acute Megakaryoblastic Leukemia of Down Syndrome. Blood 2003, 101, 4301–4304. [Google Scholar] [CrossRef]
- Hitzler, J.K. Acute Megakaryoblastic Leukemia in Down Syndrome. Pediatr. Blood Cancer 2007, 49 (Suppl. S7), 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, J.D.E.; Branstetter, C.; Ma, J.; Li, Y.; Walsh, M.P.; Cheng, J.; Obulkasim, A.; Dang, J.; Easton, J.; Verboon, L.J.; et al. Pediatric Non-Down Syndrome Acute Megakaryoblastic Leukemia Is Characterized by Distinct Genomic Subsets with Varying Outcomes. Nat. Genet. 2017, 49, 451–456. [Google Scholar] [CrossRef]
- Komatsu, N.; Suda, T.; Moroi, M.; Tokuyama, N.; Sakata, Y.; Okada, M.; Nishida, T.; Hirai, Y.; Sato, T.; Fuse, A. Growth and Differentiation of a Human Megakaryoblastic Cell Line, CMK. Blood 1989, 74, 42–48. [Google Scholar] [CrossRef]
- Sato, T.; Fuse, A.; Eguchi, M.; Hayashi, Y.; Ryo, R.; Adachi, M.; Kishimoto, Y.; Teramura, M.; Mizoguchi, H.; Shima, Y.; et al. Establishment of a Human Leukaemic Cell Line (CMK) with Megakaryocytic Characteristics from a Down’s Syndrome Patient with Acute Megakaryoblastic Leukaemia. Br. J. Haematol. 1989, 72, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Mouthon, M.A.; Freund, M.; Titeux, M.; Katz, A.; Guichard, J.; Breton-Gorius, J.; Vainchenker, W. Growth and Differentiation of the Human Megakaryoblastic Cell Line (ELF-153): A Model for Early Stages of Megakaryocytopoiesis. Blood 1994, 84, 1085–1097. [Google Scholar] [CrossRef]
- Saito, H.; Hayakawa, M.; Kamoshita, N.; Yasumoto, A.; Suzuki-Inoue, K.; Yatomi, Y.; Ohmori, T. Establishment of a Megakaryoblastic Cell Line for Conventional Assessment of Platelet Calcium Signaling. Int. J. Hematol. 2020, 111, 786–794. [Google Scholar] [CrossRef]
- Avanzi, G.C.; Lista, P.; Giovinazzo, B.; Miniero, R.; Saglio, G.; Benetton, G.; Coda, R.; Cattoretti, G.; Pegoraro, L. Selective Growth Response to IL-3 of a Human Leukaemic Cell Line with Megakaryoblastic Features. Br. J. Haematol. 1988, 69, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Brizzi, M.F.; Avanzi, G.C.; Veglia, F.; Clark, S.C.; Pegoraro, L. Expression and Modulation of IL-3 and GM-CSF Receptors in Human Growth Factor Dependent Leukaemic Cells. Br. J. Haematol. 1990, 76, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Avanzi, G.C.; Brizzi, M.F.; Giannotti, J.; Ciarletta, A.; Yang, Y.-C.; Pegoraro, L.; Clark, S.C. M-07e Human Leukemic Factor-Dependent Cell Line Provides a Rapid and Sensitive Bioassay for the Human Cytokines GM-CSF and IL-3. J. Cell. Physiol. 1990, 145, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Ogura, M.; Morishima, Y.; Ohno, R.; Kato, Y.; Hirabayashi, N.; Nagura, H.; Saito, H. Establishment of a Novel Human Megakaryoblastic Leukemia Cell Line, MEG-01, with Positive Philadelphia Chromosome. Blood 1985, 66, 1384–1392. [Google Scholar] [CrossRef]
- Drexler, H.G.; MacLeod, R.A.; Uphoff, C.C. Leukemia Cell Lines: In Vitro Models for the Study of Philadelphia Chromosome-Positive Leukemia. Leuk. Res. 1999, 23, 207–215. [Google Scholar] [CrossRef]
- Wertheim, J.A.; Forsythe, K.; Druker, B.J.; Hammer, D.; Boettiger, D.; Pear, W.S. BCR-ABL-Induced Adhesion Defects Are Tyrosine Kinase-Independent. Blood 2002, 99, 4122–4130. [Google Scholar] [CrossRef]
- Takeuchi, K.; Ogura, M.; Saito, H.; Satoh, M.; Takeuchi, M. Production of Platelet-like Particles by a Human Megakaryoblastic Leukemia Cell Line (MEG-01). Exp. Cell Res. 1991, 193, 223–226. [Google Scholar] [CrossRef]
- Murate, T.; Hotta, T.; Tsushita, K.; Suzuki, M.; Yoshida, T.; Saga, S.; Saito, H.; Yoshida, S. Aphidicolin, an Inhibitor of DNA Replication, Blocks the TPA-Induced Differentiation of a Human Megakaryoblastic Cell Line, MEG-O1. Blood 1991, 78, 3168–3177. [Google Scholar] [CrossRef] [PubMed]
- Quentmeier, H.; Pommerenke, C.; Dirks, W.G.; Eberth, S.; Koeppel, M.; MacLeod, R.A.F.; Nagel, S.; Steube, K.; Uphoff, C.C.; Drexler, H.G. The LL-100 Panel: 100 Cell Lines for Blood Cancer Studies. Sci. Rep. 2019, 9, 8218. [Google Scholar] [CrossRef]
- Quentmeier, H.; Schneider, B.; Röhrs, S.; Romani, J.; Zaborski, M.; Macleod, R.A.F.; Drexler, H.G. SET-NUP214 Fusion in Acute Myeloid Leukemia- and T-Cell Acute Lymphoblastic Leukemia-Derived Cell Lines. J. Hematol. Oncol. 2009, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Koblitz, J.; Dirks, W.G.; Eberth, S.; Nagel, S.; Steenpass, L.; Pommerenke, C. DSMZCellDive: Diving into High-Throughput Cell Line Data. F1000Research 2022, 11, 420. [Google Scholar] [CrossRef] [PubMed]
- Mihara, K.; Imai, C.; Coustan-Smith, E.; Dome, J.S.; Dominici, M.; Vanin, E.; Campana, D. Development and Functional Characterization of Human Bone Marrow Mesenchymal Cells Immortalized by Enforced Expression of Telomerase. Br. J. Haematol. 2003, 120, 846–849. [Google Scholar] [CrossRef]
- Böcker, W.; Yin, Z.; Drosse, I.; Haasters, F.; Rossmann, O.; Wierer, M.; Popov, C.; Locher, M.; Mutschler, W.; Docheva, D.; et al. Introducing a Single-Cell-Derived Human Mesenchymal Stem Cell Line Expressing HTERT after Lentiviral Gene Transfer. J. Cell. Mol. Med. 2008, 12, 1347–1359. [Google Scholar] [CrossRef]
- Galarza Torre, A.; Shaw, J.E.; Wood, A.; Gilbert, H.T.J.; Dobre, O.; Genever, P.; Brennan, K.; Richardson, S.M.; Swift, J. An Immortalised Mesenchymal Stem Cell Line Maintains Mechano-Responsive Behaviour and Can Be Used as a Reporter of Substrate Stiffness. Sci. Rep. 2018, 8, 8981. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skopek, R.; Palusińska, M.; Kaczor-Keller, K.; Pingwara, R.; Papierniak-Wyglądała, A.; Schenk, T.; Lewicki, S.; Zelent, A.; Szymański, Ł. Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research. Int. J. Mol. Sci. 2023, 24, 5377. https://doi.org/10.3390/ijms24065377
Skopek R, Palusińska M, Kaczor-Keller K, Pingwara R, Papierniak-Wyglądała A, Schenk T, Lewicki S, Zelent A, Szymański Ł. Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research. International Journal of Molecular Sciences. 2023; 24(6):5377. https://doi.org/10.3390/ijms24065377
Chicago/Turabian StyleSkopek, Rafał, Małgorzata Palusińska, Katarzyna Kaczor-Keller, Rafał Pingwara, Anna Papierniak-Wyglądała, Tino Schenk, Sławomir Lewicki, Artur Zelent, and Łukasz Szymański. 2023. "Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research" International Journal of Molecular Sciences 24, no. 6: 5377. https://doi.org/10.3390/ijms24065377
APA StyleSkopek, R., Palusińska, M., Kaczor-Keller, K., Pingwara, R., Papierniak-Wyglądała, A., Schenk, T., Lewicki, S., Zelent, A., & Szymański, Ł. (2023). Choosing the Right Cell Line for Acute Myeloid Leukemia (AML) Research. International Journal of Molecular Sciences, 24(6), 5377. https://doi.org/10.3390/ijms24065377