
Generation of a p21 Reporter Mouse and Its Use to Identify and Eliminate p21high Cells In Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation of the p21-3MR Mouse Model
2.2. GCV Treatment Selectively Kills p21high Cells In Vivo
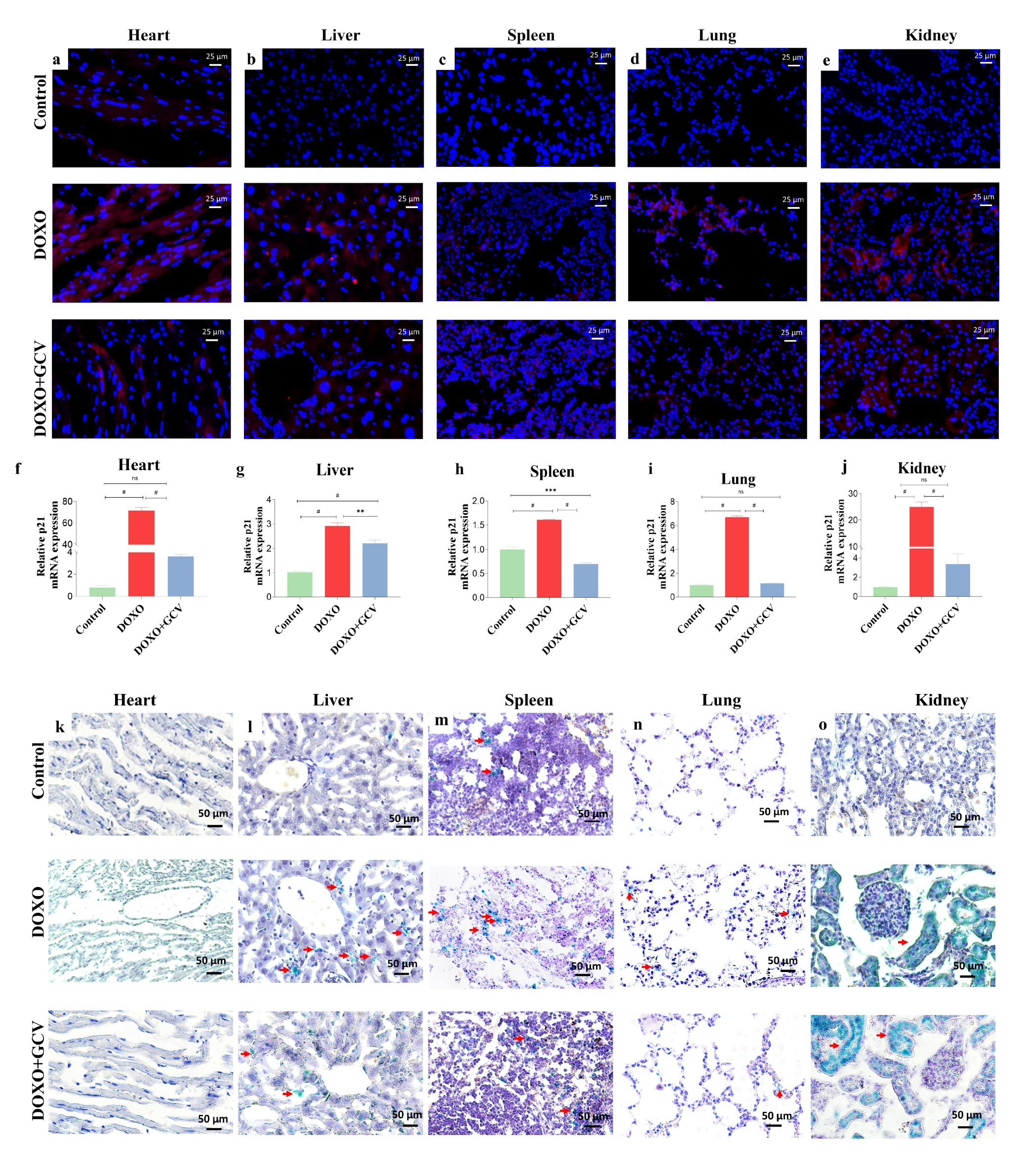
2.3. GCV Counteracts the Expression of Senescence Markers in Vital Organs
2.4. Clearance of p21high Cells Improves Histological Changes in Vital Organs
2.5. SASP Factor Changes in Vital Organs following GCV Administration
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug Treatments
4.3. Fluorescence Microscopy in Tissue Section
4.4. Immunofluorescence Staining
4.5. RNA Extraction and Quantitative Real-Time PCR
4.6. Bioluminescence Imaging
4.7. Senescence-Associated β-Galactosidase Activity
4.8. Histological Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Childs, B.G.; Durik, M.; Baker, D.J.; Van Deursen, J.M. Cellular senescence in aging and age-related disease: From mechanisms to therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Hickson, L.J.; Eirin, A.; Kirkland, J.L.; Lerman, L.O. Cellular Senescence: The Good, the Bad and the Unknown. Nat. Rev. Nephrol. 2022, 18, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M. Cellular Aging beyond Cellular Senescence: Markers of Senescence Prior to Cell Cycle Arrest in Vitro and in Vivo. Aging Cell 2021, 20, e13338. [Google Scholar] [CrossRef]
- Cohn, R.L.; Gasek, N.S.; Kuchel, G.A.; Xu, M. The Heterogeneity of Cellular Senescence: Insights at the Single-Cell Level. Trends Cell Biol. 2023, 33, 9–17. [Google Scholar] [CrossRef]
- Birch, J.; Gil, J. Senescence and the SASP: Many Therapeutic Avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef]
- Aguayo Mazzucato, C.; Andle, J.; Lee, T.B., Jr.; Midha, A.; Talemal, L.; Chipashvili, V.; Hollister-Lock, J.; van Deursen, J.; Weir, G.; Bonner-Weir, S. Acceleration of β Cell Aging Determines Diabetes and Senolysis Improves Disease Outcomes. Cell Metab. 2019, 30, 129–142. [Google Scholar] [CrossRef]
- Starling, S. Targeting β-Cell Senescence. Nat. Rev. Endocrinol. 2019, 15, 438–439. [Google Scholar] [CrossRef]
- Wang, L.; Wang, B.; Gasek, N.S.; Zhou, Y.; Cohn, R.L.; Martin, D.E.; Zuo, W.; Flynn, W.F.; Guo, C.; Jellison, E.R.; et al. Targeting P21Cip1 Highly Expressing Cells in Adipose Tissue Alleviates Insulin Resistance in Obesity. Cell Metab. 2022, 34, 75–89. [Google Scholar] [CrossRef]
- Grosse, L.; Wagner, N.; Emelyanov, A.; Molina, C.; Lacas-Gervais, S.; Wagner, K.-D.; Bulavin, D.V. Defined P16High Senescent Cell Types Are Indispensable for Mouse Healthspan. Cell Metab. 2020, 32, 87–99. [Google Scholar] [CrossRef]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Kondoh, H.; Hara, E. Targeting P21 for Diabetes: Another Choice of Senotherapy. Cell Metab. 2022, 34, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Inagaki, N.; Kondoh, H. Cellular Senescence in Diabetes Mellitus: Distinct Senotherapeutic Strategies for Adipose Tissue and Pancreatic β Cells. Front. Endocrinol. 2022, 13, 869414. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.K.; Huang, C.Y.; Chen, Y.W.; Sun, Y.T. Hyperglycemia Compromises the Ischemia-Provoked Dedifferentiation of Cerebral Pericytes through P21-SOX2 Signaling in High-Fat Diet-Induced Murine Model. Diabetes Vasc. Dis. Res. 2021, 18, 1479164121990641. [Google Scholar] [CrossRef]
- Chandra, A.; Lagnado, A.B.; Farr, J.N.; Doolittle, M.; Tchkonia, T.; Kirkland, J.L.; LeBrasseur, N.K.; Robbins, P.D.; Niedernhofer, L.J.; Ikeno, Y.; et al. Targeted Clearance of P21- but Not P16-positive Senescent Cells Prevents Radiation-induced Osteoporosis and Increased Marrow Adiposity. Aging Cell 2022, 21, e13602. [Google Scholar] [CrossRef]
- Faust, H.J.; Zhang, H.; Han, J.; Wolf, M.T.; Jeon, O.H.; Sadtler, K.; Peña, A.N.; Chung, L.; Maestas, D.R.; Tam, A.J.; et al. IL-17 and Immunologically Induced Senescence Regulate Response to Injury in Osteoarthritis. J. Clin. Investig. 2020, 130, 5493–5507. [Google Scholar] [CrossRef] [PubMed]
- Diekman, B.O.; Sessions, G.A.; Collins, J.A.; Knecht, A.K.; Strum, S.L.; Mitin, N.K.; Carlson, C.S.; Loeser, R.F.; Sharpless, N.E. Expression of P16INK4a Is a Biomarker of Chondrocyte Aging but Does Not Cause Osteoarthritis. Aging Cell 2018, 17, e12771. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.K.; Gustafson, B.; Kirkland, J.L.; Smith, U. Cellular senescence: At the nexus between ageing and diabetes. Diabetologia 2019, 62, 1835–1841. [Google Scholar] [CrossRef] [Green Version]
- Wahlmueller, M.; Narzt, M.S.; Missfeldt, K.; Arminger, V.; Krasensky, A.; Lämmermann, I.; Schaedl, B.; Mairhofer, M.; Suessner, S.; Wolbank, S.; et al. Establishment of in Vitro Models by Stress-Induced Premature Senescence for Characterizing the Stromal Vascular Niche in Human Adipose Tissue. Life 2022, 12, 1459. [Google Scholar] [CrossRef]
- Ohtani, N.; Imamura, Y. Visualizing the Dynamics of P21(Waf1/Cip1) Cyclin-Dependent Kinase Inhibitor Expression in Living Animals. Proc. Natl. Acad. Sci. USA 2007, 104, 15034–15039. [Google Scholar] [CrossRef] [Green Version]
- Laberge, R.M.; Adler, D.; DeMaria, M.; Mechtouf, N.; Teachenor, R.; Cardin, G.B.; Desprez, P.-Y.; Campisi, J.; Rodier, F. Mitochondrial DNA Damage Induces Apoptosis in Senescent Cells. Cell Death Dis. 2013, 4, e727. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, L.; Gasek, N.S.; Zhou, Y.; Kim, T.; Guo, C.; Jellison, E.R.; Haynes, L.; Yadav, S.; Tchkonia, T.; et al. An Inducible P21-Cre Mouse Model to Monitor and Manipulate P21-Highly-Expressing Senescent Cells in Vivo. Nat. Aging 2021, 1, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Carver, J.R.; Shapiro, C.L.; ASCO Cancer Survivorship Expert Panel. American Society of Clinical Oncology Clinical Evidence Review on the Ongoing Care of Adult Cancer Survivors: Cardiac and Pulmonary Late Effects. J. Clin. Oncol. 2007, 25, 3991–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, M.; Della Sala, A.; Tocchetti, C.G.; Porporato, P.E.; Ghigo, A. Metabolic Aspects of Anthracycline Cardiotoxicity. Curr. Treat. Options Oncol. 2021, 22, 18. [Google Scholar] [CrossRef]
- Margier, M.; Kuehnemann, C.; Hulo, N.; Morales, J.; Kumaar, P.V.A.; Cros, C.; Cannelle, H.; Charmetant, J.; Verdin, E.; Canault, M.; et al. Nicotinamide Mononucleotide Administration Prevents Doxorubicin-Induced Cardiotoxicity and Loss in Physical Activity in Mice. Cells 2022, 12, 108. [Google Scholar] [CrossRef]
- Xiong, Y.; Hannon, G.J.; Zhang, H.; Casso, D.; Kobayashi, R.; Beach, D. p21 is a universal inhibitor of cyclin kinases. Nature 1993, 366, 701–704. [Google Scholar] [CrossRef]
- Waga, S.; Hannon, G.J.; Beach, D.; Stillman, B. The p21 inhibitor of cyclin-dependent kinases controls DNA replication by interaction with PCNA. Nature 1994, 369, 574–578. [Google Scholar] [CrossRef]
- Oros-Pantoja, R.; Córdoba-Adaya, J.C.; Torres-García, E.; Morales-Avila, E.; Aranda-Lara, L.; Santillán-Benítez, J.G.; Sánchez-Holguín, M.; Hernández-Herrera, N.O.; Otero, G.; Isaac-Olivé, K. Preclinical Evaluation of Early Multi-Organ Toxicity Induced by Liposomal Doxorubicin Using 67Ga-Citrate. Nanotoxicology 2022, 16, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Abdelghffar, E.A.; Obaid, W.A.; Elgamal, A.M.; Daoud, R.; Sobeh, M.; El Raey, M.A. Pea (Pisum Sativum) Peel Extract Attenuates DOX-Induced Oxidative Myocardial Injury. Biomed. Pharmacother. 2021, 143, 112120. [Google Scholar] [CrossRef] [PubMed]
- Sturmlechner, I.; Zhang, C.; Sine, C.C.; van Deursen, E.-J.; Jeganathan, K.B.; Hamada, N.; Grasic, J.; Friedman, D.; Stutchman, J.T.; Can, I.; et al. P21 Produces a Bioactive Secretome That Places Stressed Cells under Immunosurveillance. Science 2021, 374, eabb3420. [Google Scholar] [CrossRef]
- Abd El-Aziz, T.A.; Mohamed, R.H.; Pasha, H.F.; Abdel-Aziz, H.R. Catechin Protects against Oxidative Stress and Inflammatory-Mediated Cardiotoxicity in Adriamycin-Treated Rats. Clin. Exp. Med. 2012, 12, 233–240. [Google Scholar] [CrossRef]
- Bin Jardan, Y.A.; Ansari, M.A. Sinapic Acid Ameliorates Oxidative Stress, Inflammation, and Apoptosis in Acute Doxorubicin-Induced Cardiotoxicity via the NF-κB-Mediated Pathway. BioMed Res. Int. 2020, 2020, e3921796. [Google Scholar] [CrossRef] [Green Version]
- Owumi, S.E.; Nwozo, S.O.; Arunsi, U.O.; Oyelere, A.K.; Odunola, O.A. Co-Administration of Luteolin Mitigated Toxicity in Rats’ Lungs Associated with Doxorubicin Treatment. Toxicol. Appl. Pharmacol. 2021, 411, 115380. [Google Scholar] [CrossRef] [PubMed]
- Brand, K.; Arnold, W.; Bartels, T.; Lieber, A.; A Kay, M.; Strauss, M.; Dörken, B. Liver-Associated Toxicity of the HSV-Tk/GCV Approach and Adenoviral Vectors. Cancer Gene Ther. 1997, 4, 9–16. [Google Scholar] [PubMed]
- Basu, A. The Interplay between Apoptosis and Cellular Senescence: Bcl-2 Family Proteins as Targets for Cancer Therapy. Pharmacol. Ther. 2022, 230, 107943. [Google Scholar] [CrossRef]
- Fletcher-Sananikone, E.; Kanji, S.; Tomimatsu, N.; Di Cristofaro, L.F.M.; Kollipara, R.K.; Saha, D.; Floyd, J.R.; Sung, P.; Hromas, R.; Burns, T.C.; et al. Elimination of Radiation-Induced Senescence in the Brain Tumor Microenvironment Attenuates Glioblastoma Recurrence. Cancer Res. 2021, 81, 5935–5947. [Google Scholar] [CrossRef]
- Prasanna, P.G.; Citrin, D.E.; Hildesheim, J.; Ahmed, M.M.; Venkatachalam, S.; Riscuta, G.; Xi, D.; Zheng, G.; van Deursen, J.; Goronzy, J.; et al. Therapy-Induced Senescence: Opportunities to Improve Anticancer Therapy. J. Natl. Cancer Inst. 2021, 113, 1285–1298. [Google Scholar] [CrossRef]
- Blokland, K.E.C.; Habibie, H.; Borghuis, T.; Teitsma, G.; Schuliga, M.; Melgert, B.; Knight, D.; Brandsma, C.-A.; Pouwels, S.; Burgess, J. Regulation of Cellular Senescence Is Independent from Profibrotic Fibroblast-Deposited ECM. Cells 2021, 10, 1628. [Google Scholar] [CrossRef]
- Nosrati, F.; Grillari, J.; Azarnia, M.; Nabiuni, M.; Moghadasali, R.; Karimzadeh, L.; Lämmermann, I. The Expression of Fibrosis-Related Genes Is Elevated in Doxorubicin-Induced Senescent Human Dermal Fibroblasts, but Their Secretome Does Not Trigger a Paracrine Fibrotic Response in Non-Senescent Cells. Biogerontology, 2023; online ahead of print. [Google Scholar]
- Meng, X.; Wang, H.; Song, X.; Clifton, A.C.; Xiao, J. The Potential Role of Senescence in Limiting Fibrosis Caused by Aging. J. Cell. Physiol. 2020, 235, 4046–4059. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J.; Baker, J.; Donnelly, L.E. Cellular Senescence as a Mechanism and Target in Chronic Lung Diseases. Am. J. Respir. Crit. Care Med. 2019, 200, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Robles, S.J.; Adami, G.R. Agents That Cause DNA Double Strand Breaks Lead to P16INK4a Enrichment and the Premature Senescence of Normal Fibroblasts. Oncogene 1998, 16, 1113–1123. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.H.; Altschuler, S.J. Patterns of Early P21 Dynamics Determine Proliferation-Senescence Cell Fate after Chemotherapy. Cell 2019, 178, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Shen, W.H. Senescence and Cancer. Cancer Transl. Med. 2018, 4, 70–74. [Google Scholar] [PubMed]
- Demaria, M.; O’Leary, M.N.; Chang, J.; Shao, L.; Liu, S.; Alimirah, F.; Koenig, K.; Le, C.; Mitin, N.; Deal, A.M.; et al. Cellular Senescence Promotes Adverse Effects of Chemotherapy and Cancer Relapse. Cancer Discov. 2017, 7, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, Z.; Ren, L.; Wei, Y.; Chen, S.; Zhao, J.; Zhu, J.; Wu, J. Generation of a p21 Reporter Mouse and Its Use to Identify and Eliminate p21high Cells In Vivo. Int. J. Mol. Sci. 2023, 24, 5565. https://doi.org/10.3390/ijms24065565
Yi Z, Ren L, Wei Y, Chen S, Zhao J, Zhu J, Wu J. Generation of a p21 Reporter Mouse and Its Use to Identify and Eliminate p21high Cells In Vivo. International Journal of Molecular Sciences. 2023; 24(6):5565. https://doi.org/10.3390/ijms24065565
Chicago/Turabian StyleYi, Zimei, Le Ren, Yu Wei, Siyi Chen, Jiechen Zhao, Jiayu Zhu, and Junhua Wu. 2023. "Generation of a p21 Reporter Mouse and Its Use to Identify and Eliminate p21high Cells In Vivo" International Journal of Molecular Sciences 24, no. 6: 5565. https://doi.org/10.3390/ijms24065565