
Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P

 , and
, and 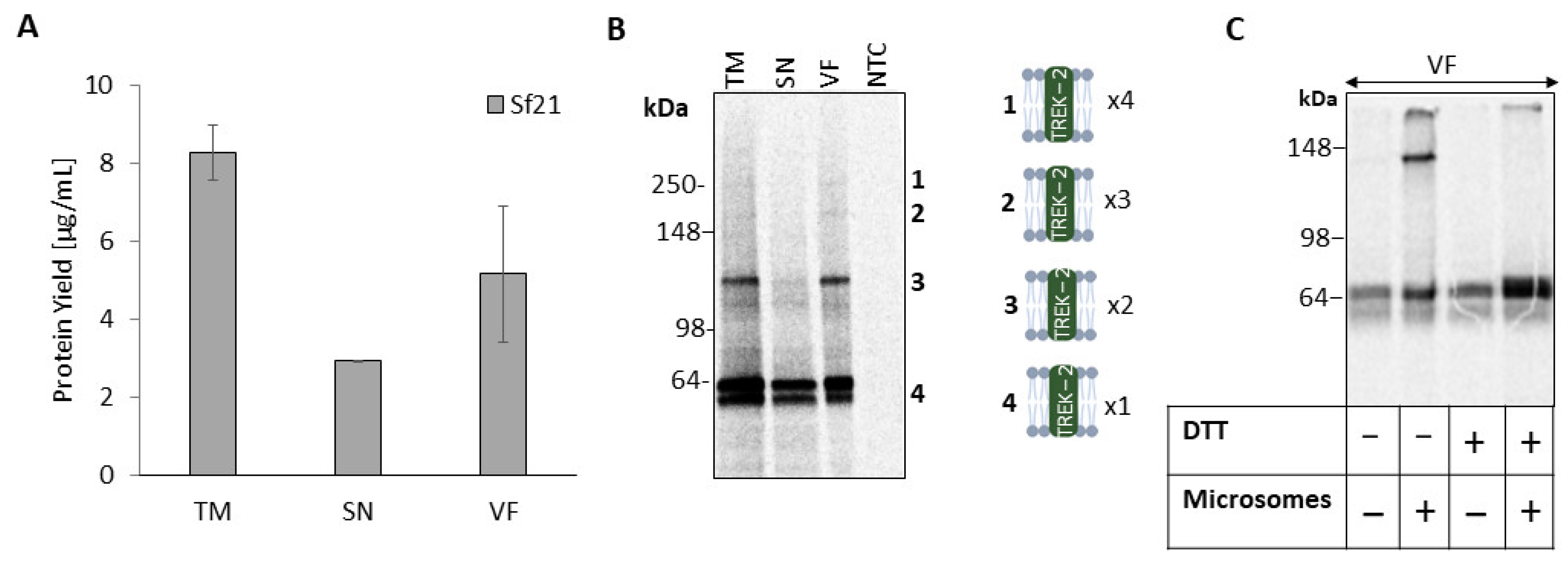{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
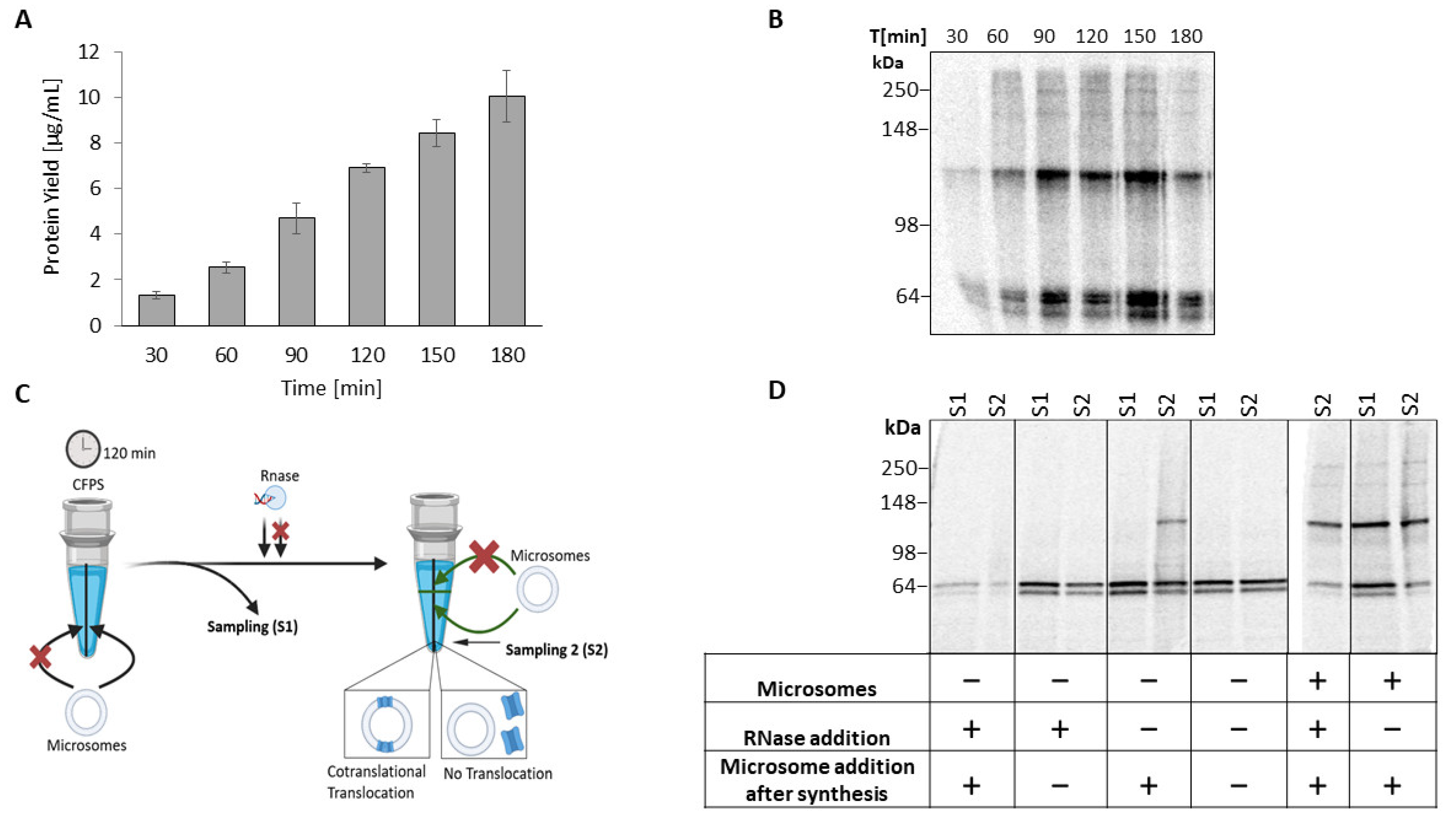
2.1. Cell-Free Synthesis of TREK-2 in a Eukaryotic Vesicle-Based System
2.2. Homomeric TREK-2 Assembly in Endoplasmic-Reticulum-Derived Microsomes
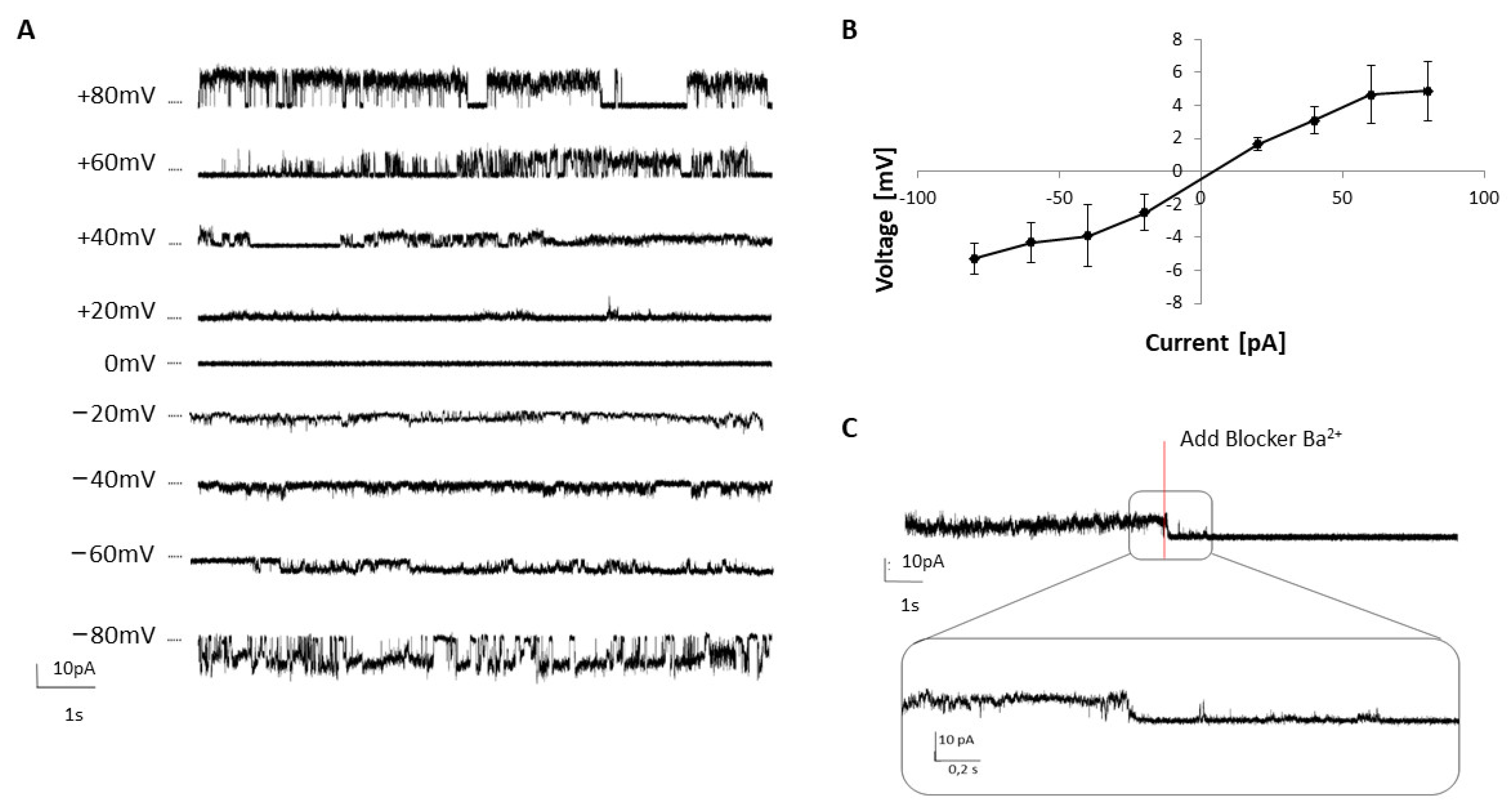
2.3. Analysis of TREK-2 Functionality
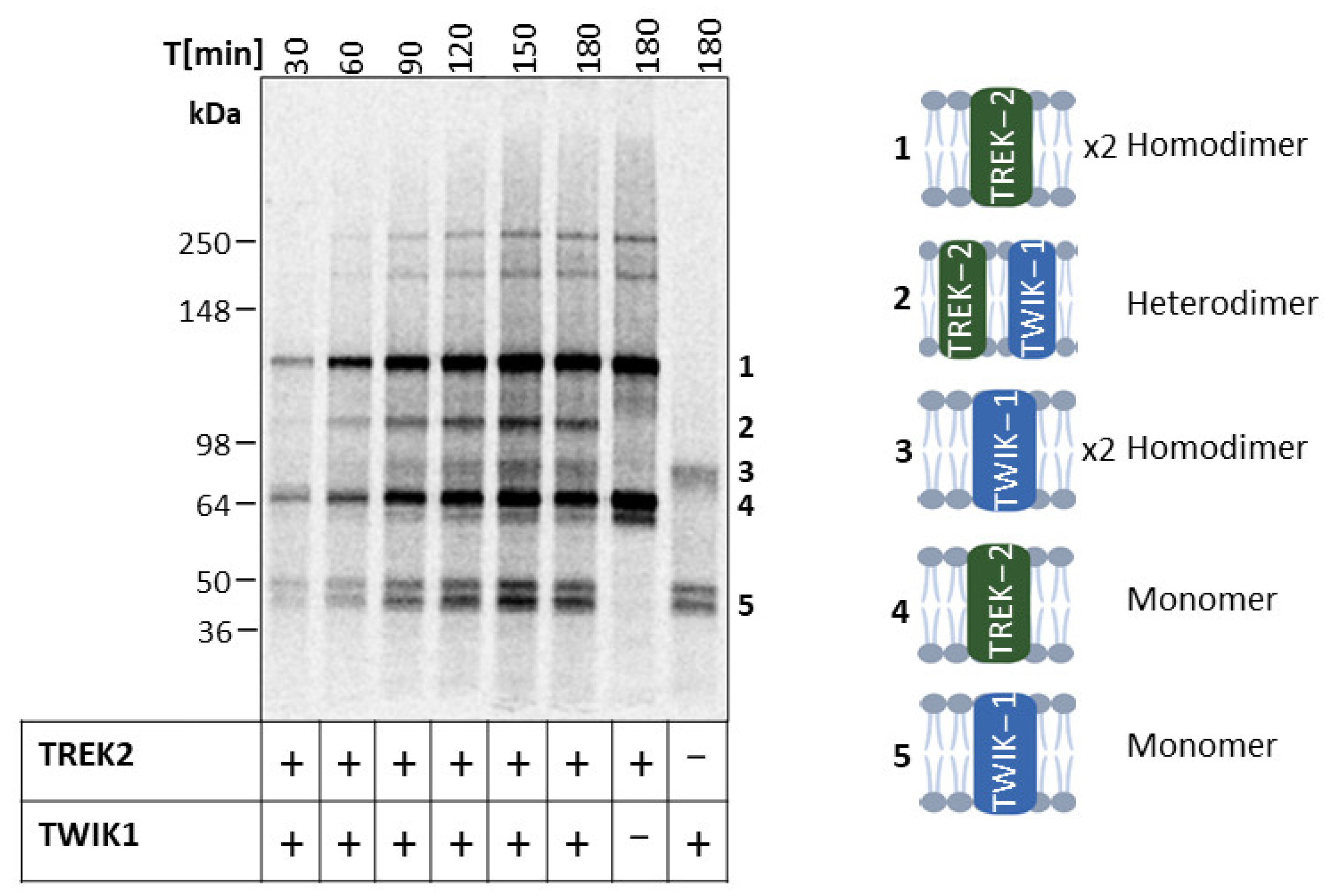
2.4. Heterodimeric Assembly of TREK-2 and TWIK-1
3. Discussion
4. Materials and Methods
4.1. Batch-Based Cell-Free Synthesis
4.2. Quantitative and Qualitative Analyses of Synthesized Proteins
4.3. The Deglycosylation Assay
4.4. Electrophysiological Recordings
4.5. The Biomolecular Fluorescence Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enyedi, P.; Czirják, G. Molecular background of leak K+ currents: Two-pore domain potassium channels. Physiol. Rev. 2010, 90, 559–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.Y.; Pike, A.C.W.; Mackenzie, A.; McClenaghan, C.; Aryal, P.; Dong, L.; Quigley, A.; Grieben, M.; Goubin, S.; Mukhopadhyay, S.; et al. K2P channel gating mechanisms revealed by structures of TREK-2 and a complex with Prozac. Science 2015, 347, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F.; Terrenoire, C.; Romey, G.; Lazdunski, M. Human TREK2, a 2P domain mechano-sensitive K+ channel with multiple regulations by polyunsaturated fatty acids, lysophospholipids, and Gs, Gi, and Gq protein-coupled receptors. J. Biol. Chem. 2000, 275, 28398–28405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brohawn, S.G.; Campbell, E.B.; MacKinnon, R. Physical mechanism for gating and mechanosensitivity of the human TRAAK K+ channel. Nature 2014, 516, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Khoubza, L.; Chatelain, F.C.; Feliciangeli, S.; Lesage, F.; Bichet, D. Physiological roles of heteromerization: Focus on the two-pore domain potassium channels. J. Physiol. 2021, 599, 1041–1055. [Google Scholar] [CrossRef]
- Czirják, G.; Enyedi, P. Formation of functional heterodimers between the TASK-1 and TASK-3 two-pore domain potassium channel subunits. J. Biol. Chem. 2002, 277, 5426–5432. [Google Scholar] [CrossRef] [Green Version]
- Blin, S.; Chatelain, F.C.; Feliciangeli, S.; Kang, D.; Lesage, F.; Bichet, D. Tandem pore domain halothane-inhibited K+ channel subunits THIK1 and THIK2 assemble and form active channels. J. Biol. Chem. 2014, 289, 28202–28212. [Google Scholar] [CrossRef] [Green Version]
- Blin, S.; Ben Soussia, I.; Kim, E.-J.; Brau, F.; Kang, D.; Lesage, F.; Bichet, D. Mixing and matching TREK/TRAAK subunits generate heterodimeric K2P channels with unique properties. Proc. Natl. Acad. Sci. USA 2016, 113, 4200–4205. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, M.; Czirják, G.; Enyedi, P. Formation of Functional Heterodimers by TREK-1 and TREK-2 Two-pore Domain Potassium Channel Subunits. J. Biol. Chem. 2016, 291, 13649–13661. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, M.; Czirják, G.; Jacobson, D.A.; Enyedi, P. TRESK and TREK-2 two-pore-domain potassium channel subunits form functional heterodimers in primary somatosensory neurons. J. Biol. Chem. 2020, 295, 12408–12425. [Google Scholar] [CrossRef]
- Hwang, E.M.; Kim, E.; Yarishkin, O.; Woo, D.H.; Han, K.-S.; Park, N.; Bae, Y.; Woo, J.; Kim, D.; Park, M.; et al. A disulphide-linked heterodimer of TWIK-1 and TREK-1 mediates passive conductance in astrocytes. Nat. Commun. 2014, 5, 3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Yarishkin, O.; Kim, E.; Bae, Y.; Kim, A.; Kim, S.-C.; Ryoo, K.; Cho, C.-H.; Hwang, E.M.; Park, J.-Y. TWIK-1/TASK-3 heterodimeric channels contribute to the neurotensin-mediated excitation of hippocampal dentate gyrus granule cells. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitz, J.; Royal, P.; Comoglio, Y.; Wdziekonski, B.; Schaub, S.; Clemens, D.M.; Isacoff, E.Y.; Sandoz, G. Heterodimerization within the TREK channel subfamily produces a diverse family of highly regulated potassium channels. Proc. Natl. Acad. Sci. USA 2016, 113, 4194–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, V.; Busserolles, J.; Christin, M.; Devilliers, M.; Poupon, L.; Legha, W.; Alloui, A.; Aissouni, Y.; Bourinet, E.; Lesage, F.; et al. Role of the TREK2 potassium channel in cold and warm thermosensation and in pain perception. Pain 2014, 155, 2534–2544. [Google Scholar] [CrossRef]
- Kawasaki, K.; Suzuki, Y.; Yamamura, H.; Imaizumi, Y. Development of a Novel Cell-Based Assay System for High-Throughput Screening of Compounds Acting on Background Two-Pore Domain K+ Channels. SLAS Discov. 2019, 24, 641–652. [Google Scholar] [CrossRef]
- Andréll, J.; Tate, C.G. Overexpression of membrane proteins in mammalian cells for structural studies. Mol. Membr. Biol. 2013, 30, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Şterbuleac, D. Molecular determinants of chemical modulation of two-pore domain potassium channels. Chem. Biol. Drug Des. 2019, 94, 1596–1614. [Google Scholar] [CrossRef]
- Mathie, A.; Veale, E.L.; Cunningham, K.P.; Holden, R.G.; Wright, P.D. Two-Pore Domain Potassium Channels as Drug Targets: Anesthesia and Beyond. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 401–420. [Google Scholar] [CrossRef]
- Stech, M.; Nikolaeva, O.; Thoring, L.; Stöcklein, W.F.M.; Wüstenhagen, D.A.; Hust, M.; Dübel, S.; Kubick, S. Cell-free synthesis of functional antibodies using a coupled in vitro transcription-translation system based on CHO cell lysates. Sci. Rep. 2017, 7, 12030. [Google Scholar] [CrossRef] [Green Version]
- Brödel, A.K.; Sonnabend, A.; Roberts, L.O.; Stech, M.; Wüstenhagen, D.A.; Kubick, S. IRES-mediated translation of membrane proteins and glycoproteins in eukaryotic cell-free systems. PLoS ONE 2013, 8, e82234. [Google Scholar] [CrossRef] [Green Version]
- Dondapati, S.K.; Kreir, M.; Quast, R.B.; Wüstenhagen, D.A.; Brüggemann, A.; Fertig, N.; Kubick, S. Membrane assembly of the functional KcsA potassium channel in a vesicle-based eukaryotic cell-free translation system. Biosens. Bioelectron. 2014, 59, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Zemella, A.; Thoring, L.; Hoffmeister, C.; Šamalíková, M.; Ehren, P.; Wüstenhagen, D.A.; Kubick, S. Cell-free protein synthesis as a novel tool for directed glycoengineering of active erythropoietin. Sci. Rep. 2018, 8, 8514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinküher, J.; Peruzzi, J.A.; Krüger, A.; Jacobs, M.L.; Jewett, M.C.; Kamat, N.P. Improving cell-free expression of membrane proteins by tuning ribosome co-translational membrane association and nascent chain aggregation. boiRxiv 2023. [Google Scholar] [CrossRef]
- Khoubza, L.; Gilbert, N.; Kim, E.-J.; Chatelain, F.C.; Feliciangeli, S.; Abelanet, S.; Kang, D.; Lesage, F.; Bichet, D. Alkaline-sensitive two-pore domain potassium channels form functional heteromers in pancreatic β-cells. J. Biol. Chem. 2022, 298, 102447. [Google Scholar] [CrossRef] [PubMed]
- Lesage, F.; Reyes, R.; Fink, M.; Duprat, F.; Guillemare, E.; Lazdunski, M. Dimerization of TWIK-1 K+ channel subunits via a disulfide bridge. EMBO J. 1996, 15, 6400–6407. [Google Scholar] [CrossRef]
- Thoring, L.; Dondapati, S.K.; Stech, M.; Wüstenhagen, D.A.; Kubick, S. High-yield production of “difficult-to-express” proteins in a continuous exchange cell-free system based on CHO cell lysates. Sci. Rep. 2017, 7, 11710. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, R.; Tanaka, T.; Hayashida, J.; Nakagita, T.; Zhou, W.; Takeda, H. Evaluation of Cell-Free Synthesized Human Channel Proteins for In Vitro Channel Research. Membranes 2022, 13, 48. [Google Scholar] [CrossRef]
- Bernstein, H.D. Cotranslational translocation of proteins into canine rough microsomes. Curr. Protoc. Cell Biol. 2001, 11, 4. [Google Scholar] [CrossRef]
- Bocksteins, E. Kv5, Kv6, Kv8, and Kv9 subunits: No simple silent bystanders. J. Gen. Physiol. 2016, 147, 105–125. [Google Scholar] [CrossRef] [Green Version]
- Guidelli, R. The common features of tetrameric ion channels and the role of electrostatic interactions. Electrochem. Commun. 2020, 121, 106866. [Google Scholar] [CrossRef]
- Mant, A.; Williams, S.; Roncoroni, L.; Lowry, E.; Johnson, D.; O’Kelly, I. N-glycosylation-dependent control of functional expression of background potassium channels K2P3.1 and K2P9.1. J. Biol. Chem. 2013, 288, 3251–3264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullrich, J.; Göhmann, P.J.; Zemella, A.; Kubick, S. Oligomerization of the heteromeric γ-aminobutyric acid receptor GABAB in a eukaryotic cell-free system. Sci. Rep. 2022, 12, 20742. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Choe, C.; Cavanaugh, E.; Kim, D. Properties of single two-pore domain TREK-2 channels expressed in mammalian cells. J. Physiol. 2007, 583, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.; Kim, Y.; Kim, D. TREK-2, a new member of the mechanosensitive tandem-pore K+ channel family. J. Biol. Chem. 2000, 275, 17412–17419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkin, D.; Cavanaugh, E.J.; Kim, D. Control of the single channel conductance of K2P10.1 (TREK-2) by the amino-terminus: Role of alternative translation initiation. J. Physiol. 2008, 586, 5651–5663. [Google Scholar] [CrossRef] [PubMed]
- Plant, L.D.; Zuniga, L.; Araki, D.; Marks, J.D.; Goldstein, S.A.N. SUMOylation silences heterodimeric TASK potassium channels containing K2P1 subunits in cerebellar granule neurons. Sci. Signal. 2012, 5, ra84. [Google Scholar] [CrossRef] [Green Version]
- Clarke, O.B.; Gulbis, J.M. Oligomerization at the membrane: Potassium channel structure and function. Adv. Exp. Med. Biol. 2012, 747, 122–136. [Google Scholar] [CrossRef]
- Fan, J.-Y.; Cui, Z.-Q.; Wei, H.-P.; Zhang, Z.-P.; Zhou, Y.-F.; Wang, Y.-P.; Zhang, X.-E. Split mCherry as a new red bimolecular fluorescence complementation system for visualizing protein-protein interactions in living cells. Biochem. Biophys. Res. Commun. 2008, 367, 47–53. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, E.; Müller, H.; Cui, B. Optical Electrophysiology: Toward the Goal of Label-Free Voltage Imaging. J. Am. Chem. Soc. 2021, 143, 10482–10499. [Google Scholar] [CrossRef]
- Szczesny, R.J.; Kowalska, K.; Klosowska-Kosicka, K.; Chlebowski, A.; Owczarek, E.P.; Warkocki, Z.; Kulinski, T.M.; Adamska, D.; Affek, K.; Jedroszkowiak, A.; et al. Versatile approach for functional analysis of human proteins and efficient stable cell line generation using FLP-mediated recombination system. PLoS ONE 2018, 13, e0194887. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullrich, J.; Ohlhoff, C.; Dondapati, S.K.; Zemella, A.; Kubick, S. Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P. Int. J. Mol. Sci. 2023, 24, 6299. https://doi.org/10.3390/ijms24076299
Ullrich J, Ohlhoff C, Dondapati SK, Zemella A, Kubick S. Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P. International Journal of Molecular Sciences. 2023; 24(7):6299. https://doi.org/10.3390/ijms24076299
Chicago/Turabian StyleUllrich, Jessica, Carsten Ohlhoff, Srujan Kumar Dondapati, Anne Zemella, and Stefan Kubick. 2023. "Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P" International Journal of Molecular Sciences 24, no. 7: 6299. https://doi.org/10.3390/ijms24076299
APA StyleUllrich, J., Ohlhoff, C., Dondapati, S. K., Zemella, A., & Kubick, S. (2023). Evaluation of the Ion Channel Assembly in a Eukaryotic Cell-Free System Focusing on Two-Pore Domain Potassium Channels K2P. International Journal of Molecular Sciences, 24(7), 6299. https://doi.org/10.3390/ijms24076299