Abstract
Silicosis, characterized by irreversible pulmonary fibrosis, remains a major global public health problem. Nowadays, cumulative studies are focusing on elucidating the pathogenesis of silicosis in order to identify preventive or therapeutic antifibrotic agents. However, the existing research on the mechanism of silica-dust-induced pulmonary fibrosis is only the tip of the iceberg and lags far behind clinical needs. Idiopathic pulmonary fibrosis (IPF), as a pulmonary fibrosis disease, also has the same problem. In this study, we examined the relationship between silicosis and IPF from the perspective of their pathogenesis and fibrotic characteristics, further discussing current drug research and limitations of clinical application in silicosis. Overall, this review provided novel insights for clinical treatment of silicosis with the hope of bridging the gap between research and practice in silicosis.
1. Introduction
1.1. Silicosis
The prevalence of crystalline silicon dioxide dust is widespread in many areas [1]. As a result of the continuous inhalation of silica particles, many workers develop silicosis, an irreversible and incurable disease [2]. Silicosis is a chronic interstitial lung disease characterized by fibrosis, inflammation, and destruction of the pulmonary structures. It causes pulmonary hypertension, progressive dyspnoea and death from respiratory insufficiency [3]. Many measures have been taken in recent decades to protect workers in the workplace, but millions of workers still suffer from silicosis [4].
Silicosis is a severe concern in construction and mining workers, particularly young workers, who are exposed to quartz conglomerates during sandblasting, bolting, cutting, shaping, and installing kitchen countertops [5,6,7]. Recent studies have also indicated that exposure to nanosilica can cause inflammation and fibrosis in the lungs. The risk of nanosilica exposure in this emerging industry is noteworthy, despite the lack of reported cases [8]. The increasing number of silicosis cases worldwide presents new challenges for prevention in many countries [4]. The present circumstance underscores the significance of exercising caution in authorizing development of emerging industries and implementing early identification and control measures from a public health perspective. Furthermore, it is crucial to accelerate the discovery of remedies for silicosis from a clinical treatment standpoint.
1.2. Animal Models of Silicosis
Mice and rats have become popular choices for experimentation due to their diminutive size, ease of breeding and maintenance in large numbers, rapid life cycle and convenient sample collection. Among the commonly used strains, C57BL/6 mice and Wistar rats are frequently employed [9]. Interestingly, no overall difference was found in progressive fibrosis between female and male mice with silicosis [10]. Notably, the most effective volume of silica suspension instilled into the noses of inbred mice C57BL/6J, which are highly susceptible to silicosis, was 80 μL via repeated inhalation. After repeated exposure to 20 mg/mL, this mode of inhalation resulted in a high rate of successful entry into the lungs and a high survival rate [11].
Replicating and simulating the entire process of silicosis resulting from prolonged exposure to industrial dust poses significant challenges due to the physiological limitations of animals. Additionally, the combined toxic consequences of silica and any mineral, metal or bacterial material on its surface cannot be assessed in animal models compared to the actual inhalation of silicosis patients [12]. Both silica aerosols and suspensions must be disinfected prior to modeling, and the environments for establishing animal models of silicosis are different from the reality of workers inhaling silica particles, so there may be significant limitations between basic experiments and clinical drugs in the treatment of silicosis. Therefore, exploring and establishing a dynamic animal model highly similar to human silica inhalation is also a key direction for future research.
1.3. Pathogenesis of Silicosis
The pathogenesis of silicosis is not fully understood, and the disease is complex [12]. Although further research is required to clarify the role of intricate signaling pathways [13], multiple pathways are thought to be involved in the development of silicosis (Figure 1). Silica-induced lung injury is characterized by various mechanisms, including direct cytotoxic effects on macrophages, activation of macrophage surface receptors, lysosomal rupture, reactive oxygen species (ROS) production, inflammasome activation, cytokine and chemokine production, apoptosis/softening, and lung fibrosis [14].
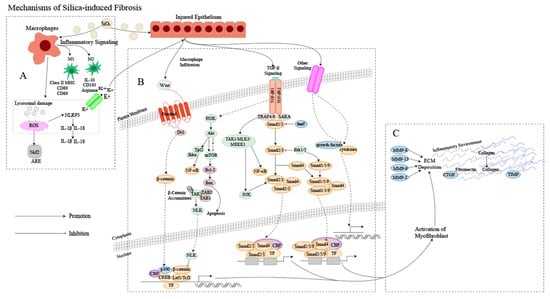
Figure 1.
Mechanism of silica-induced fibrosis. (A) Alveolar macrophages (AMs) engulf silica dust, causing them to turn into dust cells. Subsequently, AMs may synergize with alveolar epithelial cells (AECs) to release a large amount of ROS to participate in oxidative stress reactions, activate NOD-like receptor thermal protein domain associated protein 3 (NLRP3) inflammatory bodies through lysosomal damage and potassium outflux, and activate the release of inflammatory mediator interleukin (IL) -1β, IL-18 and other cytokines inducing epithelial–mesenchymal transition (EMT). Meanwhile, AMs can polarize into M1 and M2 types, playing a role in promoting inflammation, fibrosis, and antigen presentation, increasing the proliferation of lung fibroblasts and collagen synthesis and secretion, and promoting the formation of fibrosis through apoptosis and autophagy. (B) Ongoing damage and damage to lung cells by silica lead to pathological overdeposition of extracellular matrix (ECM) proteins accompanied by upregulation of myofibroblast activity, resulting in a chronic inflammatory environment of macrophage and immune cell infiltration. In this cellular environment, cytokines and growth factors are released in large quantities, activating many signaling cascades, including members of the transforming growth factor-beta (TGF-β) family and Wingless/Int (Wnt) 1, the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway and other pathways. (C) Fibroblasts then aggregate in the area of injury, and the combination of ECM degradation by MMPs and excessive collagen deposition leads to granuloma formation and lung tissue remodeling.
2. Characteristics of Pulmonary Fibrosis
It is generally accepted that the pathogenesis of pulmonary fibrosis is characterized by persistent microdamage to the alveolar epithelium with an abnormal repair process. This process is characterized by abnormal activation of myofibroblasts, excessive accumulation of ECM, and lung scar formation, ultimately leading to structural destruction and loss of function in the lungs. Apart from silicosis, idiopathic pulmonary fibrosis (IPF) is also a prevalent disease that results in pulmonary fibrosis [15].
2.1. Characteristics of IPF
The origin of IPF, much like silicosis, remains unclear, with a convoluted pathogenesis likely involving multiple interconnected signaling pathways, including the TGF-β/Smad signaling pathway, Wnt/β-catenin signaling pathway, platelet-derived growth factor (PDGF) signaling pathway, PI3K/AKT signaling pathway and other signaling pathways [16]. IPF is thought to be a consequence of damage to the alveolar epithelium and abnormal wound healing, but it has also been shown that both genetic factors and environmental conditions can significantly contribute to the development of the disease [17,18]. The condition is characterized by subpleural basal fibrosis, honeycomb changes, and collagen and ECM deposition, which ultimately result in life-threatening structural changes in lung tissue and loss of pulmonary ventilation and diffusion function.
Alveolar epithelial damage, caused by external factors (infection, toxins, smoke) or internal factors (inflammation, oxidative stress, abnormal immune response), leads to the release of fibrogenic cytokines, including TGF and tumor necrosis factor (TNF), as well as growth factors such as PDGF and connective tissue growth factor (CTGF) [19]. Elevated levels of these fibrogenic cytokines and growth factors, both locally and systemically, stimulate the activation and proliferation of lung fibroblasts to some extent. Upon activation, fibroblasts differentiate into pulmonary myofibroblasts, which are responsible for the excessive production of ECM proteins in fibrotic lung tissue. These myofibroblasts also regulate the balance between MMP and tissue inhibitors of metalloproteinases (TIMPs), thereby facilitating the process of IPF [20].
2.2. The Relationship between Silicosis and IPF
2.2.1. Cause of Disease
The inhalation of free silica dust in the air is the main cause of silicosis. The emergence of silicosis is closely linked to the volume, structure and dimensions of the silica particles inhaled [21], whereas IPF is a particular type of interstitial pneumonia that is fibrosing, chronic and progressive, but for which the origin is yet to be determined [22].
Despite the fact that occupational exposure can induce IPF to some extent, a plethora of epidemiological investigations have demonstrated a significant link between smoking, chronic viral infections, and the genetics of IPF [22,23,24].
2.2.2. Pathogenesis
Extensive research has revealed that the development of silicosis fibrosis is not solely due to one factor, but rather a complex outcome resulting from various factors and links [25]. The primary pathogenic mechanisms of silicosis involve direct cytotoxic effects, the generation of ROS and reactive nitrogen radicals, the release of inflammatory chemokines, the initiation of fibrotic pathways and cell death [4]. The cellular molecule and gene transcription regulation fields are also being explored in relation to silicosis [26]. Similarly, the pathogenesis of IPF has also not been fully elaborated, but relevant studies have shown that oxidative stress, inflammatory response, a series of cytokines and their related signal transduction pathways are also involved in the disease process [27].
Cellular senescence, which includes molecular changes such as telomere shortening, is involved in the pathogenesis of various chronic diseases, including lung diseases [28]. Telomere shortening may be a common causative feature of the development of IPF and silicosis [29,30]. Moreover, epigenetic regulation is a newly discovered mechanism of silicosis and IPF, but further research is necessary to fully understand this process [31,32].
Silicosis and IPF are respiratory diseases that cause damage to the lungs. In response to the inflammatory process, fibroblasts proliferate and produce excessive collagen fibers, leading to the deposition of ECM lung tissue remodeling, ultimately resulting in impaired lung function [33,34]. Moreover, the two diseases share similarities in the upregulation of TGF-β and extracellular signal-regulated kinase (ERK) signaling pathways in cytokine and growth factor pathways, and a relationship with autophagy [35]. While the immediate causes of silicosis and IPF may differ, the overall mechanisms of the subsequent profibrotic reaction are comparable, being characterized by ECM deposition and fibroblast proliferation [36]. Therefore, potential therapeutic drugs for the treatment of silicosis may be sought from IPF.
2.2.3. Symptoms and Complications
Silicosis and IPF share certain symptoms, such as dyspnoea and loss of appetite. However, patients with silicosis may experience additional manifestations, such as chest pain, pulmonary dysfunction, low-grade fever, night sweats, and active shortness of breath, while IPF typically presents as cough and sputum production [37]. In some severe cases, patients with IPF may also exhibit general discomfort, including weakness and joint pain.
The development of silicosis can result in various complications, including tuberculosis, chronic obstructive pneumonia, and rheumatoid arthritis [38]. Similarly, individuals with IPF may experience pulmonary hypertension, acute exacerbation of pulmonary fibrosis, respiratory tract infections, acute coronary syndrome, and thromboembolic disease [39]. However, as the diseases advance, both silicosis and IPF can increase the likelihood of developing lung cancer [40,41,42]. Ultimately, respiratory failure is the primary cause of mortality in patients with these conditions [43]. Consequently, we can investigate the symptomatic treatment drugs used in the treatment of IPF to identify potential therapeutic agents that can alleviate silicosis.
3. Treatment of Silicosis and IPF
The clinical management of silicosis remains a challenge, as there is no definitive index for assessing the extent of pulmonary fibrosis and no breakthrough drugs or targeted treatments are available. Whole lung lavage (WLL) has been demonstrated to control the symptoms of silicosis and improve the quality of life, particularly in the early stages of the ailment. Moreover, lung transplant is a well-established treatment for end-stage silicosis [44,45]. Unfortunately, the scarcity of donor lungs, numerous contraindications, and the high risks associated with the surgery all contribute to the difficulty of carrying out transplantation [46]. In recent years, researchers have developed new drugs and new approaches for the treatment of silicosis, but most of the research is still in the basic research stage, including animal experiments and cell experiments.
3.1. Drug Research Targeting Mechanisms of Silicosis
The study findings have demonstrated the considerable scope of pharmaceuticals in the treatment of silicosis fibrosis. These drugs can address one or more fundamental mechanisms of silicosis, mitigating the inflammation and/or fibrosis triggered by silica.
3.1.1. Oxidative Stress Response
The oxidative stress encountered during pulmonary fibrosis is closely associated with both the nuclear factor kappa-B (NF-κB) signaling pathway, the kelch-like ECH-associated protein 1 (Keap1)/NF-E2 p45-related factor 2 (Nrf2)/antioxidant-responsive element (ARE) pathway and the NADPH oxidase (NOX) 4-Nrf2 signaling cascade [47]. According to available evidence, it has been suggested that the administration of dioscin, dihydroquercetin, and quercetin may have a beneficial effect on pulmonary fibrosis by impeding the infiltration of macrophages, B lymphocytes, and T lymphocytes into the lung tissue [48,49,50]. Oleanolic acid has been found to be advantageous in the management of pulmonary fibrosis, most likely due to its capacity to diminish serum TNF-α levels, decrease collagen content in lung tissues, and hinder oxidative stress and NF-κB activation [51]. Moreover, the inhibition of nuclear translocation of NF-κB p65 by compounds such as quercetin, bletilla striata polysaccharide, holly and ursolic acid can effectively suppress inflammation and provide some relief from lung fibrosis [51,52,53]. An investigation has uncovered that tanshinone IIA (tan IIA), earthworm extract and emodin possess the ability to effectively suppress the EMT and TGF-β1/Smad signaling pathways. Furthermore, they can reduce silica-induced oxidative stress by activating the Nrf2 signaling pathway [54,55,56,57,58]. Oxidation, and imbalance in oxidation, can be attributed to the fluctuations in the levels of antioxidant enzymes linked to pulmonary fibrosis, as well as ROS catalyzed by NOX2/4. Studies have revealed that white tea extract and astragalus could enhance the level of antioxidant enzymes in lung tissue, strengthen the antioxidant capacity of lung tissue, and exhibit antifibrotic properties [59].
The fibrosis process in silicosis may be impacted by the use of traditional Chinese medicine (TCM) compound preparations that target oxidative stress. A recent study has revealed that the low-dose nebulized inhalation of Chinese herbal preparations can reduce the levels of malondialdehyde (MDA) and interferon γ, while also improving pulmonary fibrosis and inflammation in silicosis [60]. Other compounds, such as panicolin, bletilla striata polysaccharide, small molecule components, and N-acetylcysteine (NAC) have also been found to be effective in controlling the progression of silicosis. These drugs work by modulating the hydroxyproline levels and regulating factors such as superoxide dismutase (SOD) and MDA in the oxidation system [61]. Studies have also provided evidence that hesperetin and panicolin can enhance the activity of antioxidant enzymes including SOD and glutathione peroxidase (GPx) in animals. Furthermore, these compounds can reduce the levels of lipid peroxidation and TNF-α and slow down the progression of pulmonary fibrosis [62]. Moreover, fullerene nanoparticles (FNs) possess an impressive ability to clear ROS, enabling them to effectively inhibit the secretion of mature IL-1β and the influx of neutrophils [63]. It is worth noting that lung administration of FNs does not cause significant toxicity [63]. Research has also shown that combination therapy with astragalus, huangjin and other drugs for pneumoconiosis complicated with chronic obstructive pulmonary disease can effectively reduce SOD, MDA, glutathione peroxidase, and pulmonary fibrosis in patients, thereby impeding the advancement of pulmonary fibrosis [59]. However, in most cases, fibrosis has already begun at the time of diagnosis, so controlling early inflammatory indicators has little effect on reversing the fibrosis.
3.1.2. Autophagy and Apoptosis
Currently, research has shown that apoptosis of AM induced by SiO2 can be regulated by various intracellular pathways, including the mitochondria-mediated intracellular apoptosis program [64], NF-κB signaling pathway [65], factor-related apoptosis (Fas)-mediated exogenous pathway [66], p53 signaling pathway [67], endoplasmic reticulum stress [68], PI3K/AKT/mTOR signaling pathway [69], Janus kinase (JAK)2/signal transducer and activator of transcription (STAT)3 signaling pathway [70], and others. Studies have demonstrated that emodin can inhibit silica-induced apoptosis and exert antifibrotic effects by increasing the expression of anti-apoptotic protein B-cell lymphoma-2 (BCL2) and decreasing the expression of pro-apoptotic protein-BCL2-associated X(Bax) protein [58]. Similarly, dioscin can promote AM autophagy, enhance the clearance of mitochondria damaged by silica dust, and reduce the activation of mitochondria-mediated apoptosis pathway, so that AM resists apoptosis caused by silica dust and reduces the secretion of pro-inflammatory and profibrotic factors [48]. Moreover, AKEX0011, which is based on the phenylpyridone scaffold of pirfenidone, has been found to inhibit the Arabidopsis Serine/Threonine Kinase1 (ASK-1)/P38 signaling pathway and regulate the polarization of macrophages by reducing the phosphorylation of NF-κB/peroxisome proliferator-activated receptor gamma (PPAR-γ) proteins [71]. Atractylenolide III has also been shown to inhibit autophagy through an mTOR-dependent mechanism, improve the blockade of AM autophagy degradation, and reduce silicon-induced AM apoptosis [72]. Additionally, kaempferol has been found to reduce the phosphorylation levels of PI3K, AKT and mTOR proteins in lung tissues of mice with pulmonary fibrosis, thereby inhibiting the PI3K/AKT/mTOR signaling pathway and increasing the level of autophagy to exert antifibrotic effects [73]. Pirfenidone has also been found to reduce pulmonary fibrosis in silicosis rats by reducing IL-17A secretion through the JAK2/STAT3 signaling pathway and inhibiting macrophage polarization [74].
3.1.3. Regulation of Signaling Pathways Related to EMT
TGF-β is a crucial regulator of EMT, which reduces the expression of epithelial markers such as E-cadherin and α-catenin while increasing the expression of mesenchymal markers such as N-cadherin, vimentin, and alpha-smooth muscle actin(α-SMA) [75]. Certain natural compounds such as emodin, quercetin, tadalafil, and sodium ferulate have been found to increase E-cadherin levels and reduce the expression of Vimentin, α-SMA, Col-I, TNF-α, IL-1β, and pro-inflammatory factors TGF-β1, thereby regulating EMT to relieve fibrosis in silicosis [76,77]. Tamoxifen, on the other hand, reduces the serum TGF-β1 content in rats in the model group in a dose-dependent manner, effectively inhibiting the process of silicosis [78], but also exhibits certain hepatotoxic effects. Interestingly, AKEX0011 has also been found to reduce the infiltration of neutrophils and macrophages in lung tissue and decrease the protein levels and mRNA expression of fibrosis-associated proteins [71]. Moreover, natural compounds such as ursolic acid and astragaloside IV can deactivate the mitogen-activated protein kinase (MAPK) signaling pathway, regulate PI3K/AKT/mTOR, inhibit the hyperphosphorylation of forkhead box O3 (FOXO3a) to inhibit EMT, and slow the progression of pulmonary fibrosis [79,80]. Furthermore, bone morphogenetic protein (BMP)-7 can attenuate silica-induced silicosis fibrosis by modulating the balance between TGF-β/Smad and BMP-7/Smad signaling pathways, but its preventive effect is more significant than its therapeutic effect [81]. It is worth noting that TGF-β1-mediated Smad-dependent pathways and Smad-independent pathways can independently regulate the fibrotic response or exert their effects through interaction with Smad proteins. Buyang Huanwu Tang has been found to regulate multiple signaling pathways, including TGF-β1/Smad2/3 and PIK3/AKT, resulting in a reduction of pulmonary fibrosis and inflammatory changes in silicosis [82]. Similarly, Dahuang Zhechong pills have been shown to effectively inhibit the p38 MAPK/NF-κB/TGF-β1 pathway and TGF-β1/Smad pathway, thereby reducing and eliminating the persistent inflammatory stimulation caused by SiO2 in the lungs [83]. Natural medicines work through these pathways for different periods and have different antifibrotic effects. However, ensuring that a single drug or combination of drugs works in the body without damaging other functions remains a complex problem that requires further investigation.
3.1.4. Blocking Silicosis Fibrosis by Targeting Fibroblasts
Pulmonary fibrosis is characterized by excessive secretion of collagen, fibronectin, and elastin by myofibroblasts, leading to an imbalance in MMP/TIMP regulation and accumulation of the ECM [84]. Quercetin has been found to reduce the expression of MMP-2 and MMP-9, thereby inhibiting the formation of the ECM and reducing pulmonary fibrosis [85,86,87]. Schisandrin B has also been shown to inhibit the expression of MMP-2, slowing the onset of pulmonary fibrosis in rats induced by silica [88]. Additionally, gallus domesticus extract has been found to reduce the content of FNs, Col-I, MMP-9 and MMP-12 in bronchoalveolar lavage fluid (BALF) in rats [89]. Dasatinib has been found to induce macrophage bias towards the M2 macrophages phenotype, improving lung mechanics in mouse models of acute silicosis by downregulating the expression of IL-1β, TNF-α and TGF-β proteins in lung tissue and upregulating the expression of arginase and MMP-9 [90]. While drugs can regulate fibroblast activation and ECM degradation through multiple signal transduction pathways, they cannot reverse transformed myofibroblasts and cure silicosis.
3.1.5. Other Mechanisms to Prevent and Treat Silicosis Fibrosis
In addition to the aforementioned studies, researchers have been investigating alternative mechanisms through which drugs can affect silicosis fibrosis. For instance, kaempferol has been found to restore silica-induced microtubule-associated protein 1A/1B-light chain 3 (LC3) lipidation without increasing p62 levels [73]. Similarly, metformin has been shown to inhibit the inflammatory response in macrophages, reduce TGF-β1-stimulated fibroblast activation in lung fibroblasts via the AMPK-dependent pathway, increase the expression levels of p-AMPK, LC3B and Beclin1 proteins, reduce the levels of phosphorylated mammalian rapamycin (p-mTOR) and p62 proteins in vivo, and activate autophagy to inhibit silica-induced pulmonary fibrosis [91]. Quercetin reduces the accumulation of myofibroblasts and restores fibroblast apoptosis sensitivity by upregulating the Fas ligand receptor and the expression of caveolin-1 [52]. Dihydroquercetin inhibits ferritin phagocytosis-mediated iron death in cells and improves silica-induced pulmonary fibrosis [49], while lysophosphatidylcholine acyltransferase 1 (LPCAT1) alters the balance between phosphatidylcholine and lysophosphatidylcholine and inhibits the development of silicosis in mice [92].
Moreover, the combination of tetrandrine tablets and matrine injection has been shown to have a low incidence of adverse reactions and to be able to improve lung ventilation function, alleviate symptoms, and exhibit significant clinical value [93]. The combined use of desipramine and NAC has been found to effectively suppress the inflammatory response and delay the progression of silicosis fibrosis in a synergistic manner [61]. This synergy has also been observed with the combination of NAC and tetrandrine [94]. However, drug combinations not only show synergistic effects, but also are accompanied by the side effects of multiple drugs.
In summary, it has been discovered that numerous drug components and TCM compound preparations can hinder the development of silicosis fibrosis by intervening in the silicosis TGF-β/Smad signaling pathway, oxidative stress mechanism, apoptosis, and autophagy. Additionally, various pathways have been observed to interact with each other, such as emodin, tan IIA, curcumin, and other drugs, which can exert anti-inflammatory and antifibrotic effects through different intervention mechanisms [95]. In the future, apart from conducting more in-depth research on the pathogenesis and progression mechanism of silicosis, it is vital to focus on the interaction between signaling pathways and drug intervention targets, and to prioritize the transformation of basic research into clinical practice. Furthermore, it is essential to conduct further research on the diagnosis and treatment technology of TCM. A specific and standardized diagnosis and treatment plan for the combination of traditional and modern medicine for the treatment of silicosis should be established as soon as possible. Individualized treatment plans should be formulated based on the patients’ specific conditions to improve their quality of life, reduce economic pressure, and promote their return to social life faster and more effectively.
3.2. Antifibrosis Treatment Drugs for IPF
Numerous studies have been found that by inhibiting the NF-κB signaling pathway, parthenolide, hesperidin and dehydrocostus can inhibit the early inflammatory response, thereby exerting an antifibrotic effect [96,97,98]. Moreover, sulforaphane, dihydroartemisinin, melatonin and ginkgo biloba extract have been found to reduce oxidative stress to exert anti-pulmonary-fibrosis effects [99,100,101,102,103]. Similarly, the total extract of Yupingfeng and the combination of tan IIA and puerarin can exert antifibrotic effects by regulating the PI3K/AKT/mTOR signaling pathway and JAK/STAT signaling pathway [104,105,106]. As for regulating signaling pathways related to EMT, asiatic acid and oridonin inhibit the expression of TGF-β1 in lung tissue, accompanied by a decrease in Col-I, Col-III, α-SMA and TIMP-1, as well as inactivation of Smads and ERK1/2 [107,108]. Moreover, juglanin and Rhodiola rosea L. significantly attenuate bleomycin (BLM)-induced pulmonary fibrosis by reducing the expression of fibrotic features such as TGF-β1, fibronectin, MMP-9, α-SMA, and Col-I [109], particularly in a dose-dependent manner in the case of Rhodiola rosea L [110]. Further investigation indicated that tannic acid treatment suppresses BLM-induced phosphorylation of ERK1/2 in lungs [111]. Moreover, nimbolide regulates autophagy signaling by inhibiting LC3 and p62 expression and increasing Beclin-1 expression [112]. Studies have found that treatment with Olanpingensis polysaccharides can alleviate pulmonary fibrosis progression, mainly by reducing the recruitment of macrophages to the lungs [113].
Numerous studies have demonstrated the potential of various TCMs, including Xin Jia Xuan Bai Cheng Qi Decoction extract, Renshen Pingfei Decoction, Yangyin Yiqi Mixture, and Qing-Xuan Granules, to ameliorate the progression of pulmonary fibrosis. The mechanism may involve the suppression of the TGF-β1/Smad signal pathway and EMT in BLM-induced pulmonary fibrosis [114,115,116,117]. Other drugs, such as Feifukang, Modified Kushen Gancao Formula, Jinshui Huanxian Formula, and Yifei Sanjie Formula, target different pathways to alleviate pulmonary fibrosis [118,119,120,121]. Combination therapies involving traditional Chinese and modern medicines have also been studied. For example, Salvia Miltiorrhiza and Ligustrazine have been found to be safe and effective in repressing BLM-induced pulmonary fibrosis, possibly by modulating the expression of TNF-α and TGF-β1 [122]. Dexamethasone combined with berberine has also shown antifibrotic effects, which involve inhibiting C-X-C motif chemokine ligand (CXCL)-14 and MMP-2/MMP-9 expression and preventing the activation of Smad2/3 and hedgehog signaling pathways [123].
Since there are similarities in the mechanisms of silicosis and IPF, and some drugs have been shown to have a role in both silicosis and IPF animal trials (Table 1), it is worth considering the option of obtaining treatment drugs for silicosis from those used for IPF.

Table 1.
Drug candidates for the treatment of silicosis.
3.3. Other Potential Therapies
3.3.1. Stem Cell Therapy
Mesenchymal stem cells (MSCs) are considered relatively safe due to their abundant sources, ease of isolation and culture, low immunogenicity, and secretion factors that can reduce inflammation [130]. Therefore, stem cell therapy has shown significant promise in the treatment of silicosis. Preclinical studies have shown that the administration of MSCs through endotracheal or tail vein effectively inhibits inflammation and fibrosis in mouse models of silicosis, leading to therapeutic benefits [131,132]. Moreover, previous research suggests that N-acetyl-seryl-aspartyl-lysyl-proline (Ac-SDKP) may alleviate the fibrotic symptoms of silicosis by regulating endoplasmic reticulum stress [133]. Averyanov’s study demonstrated that transplantation of bone-marrow-derived MSCs at high cumulative doses (1.6 × 109/mL) is well tolerated and safe in patients with pulmonary fibrosis, with only minor side effects such as fever. These findings confirm the safety, tolerability, and potential benefits of high-dose MSCs, and may pave the way for future stem cell transplantation trials [134]. In recent years, induced pluripotent stem cells have gained popularity in the research community. These cells can be used to cultivate numerous mature cell types and create lung organoids that can aid in the development of personalized treatments for silicosis [135].
Although this approach has potential, there are several issues that need to be resolved. Firstly, ongoing assessment of the clinical safety and effectiveness of these pharmacotherapies is necessary. Secondly, stem cell therapy is still in its early stages and faces numerous technical challenges. Thirdly, the process of conducting clinical trials and addressing ethical considerations for new drug targets can be lengthy. Lastly, the exorbitant cost of personalized and experimental therapies limits their widespread clinical application due to financial constraints.
3.3.2. Antifibrotic Target Therapy
The advent of high-throughput omics technology and its amalgamation with bioinformatics has brought about a paradigm shift in gene editing technology, presenting a viable remedy for investigating the causation and management of silicosis.
Recent genomic analyses have revealed that certain single nucleotide polymorphisms (SNPs) are connected to an increased risk of silicosis, particularly rs73329476 and rs12812500, which are linked to the development of pneumoconiosis [136]. By conducting weighted gene co-expression network analysis, Jiaqi Lv et al. identified silicosis-related modules and pivot genes and found that the Hippo signaling pathway plays a beneficial role in silicosis fibrosis. This discovery has helped to elucidate the precise mechanisms of silica-induced pulmonary fibrosis and identify the molecular initiation events and adverse outcome pathways of silicosis [137]. In the field of epigenomics, research has demonstrated that certain miRNAs are significantly linked to the development and progression of pneumoconiosis and may serve as non-invasive biomarkers and prognostic indicators for early pneumoconiosis. For example, miRNA-29, which regulates the Wnt/β-catenin pathway, and miRNA-326, which targets tumor necrosis factor superfamily-14 and polypyrimidine bundle-binding proteins, have been shown to inhibit fibrosis in silicosis [138,139]. Furthermore, miRNAs have serum-detectable, morphologically stable, and reusable freeze-thaw properties, making them promising biomarkers for the early diagnosis of silicosis [140]. Finally, Lv-shβ-catenin has been found to reduce the expression of MMP-2 and MMP-9, reducing pulmonary fibrosis and inhibiting the formation of ECM [85,87]. These findings have important implications for the prevention and treatment of silicosis and highlight the potential of genomic and epigenomic research to improve our understanding of this debilitating disease.
The use of transcriptomics and proteomics has led to the identification of a variety of protein-coding genes and proteins that are differentially expressed in silicosis [141]. These findings suggest potential targets and signaling pathways that may play a significant role in lung disease. Bo C and Mingyao Wang et al. conducted a study using different clusters for pathway enrichment. They classified differentially significant proteins and found that although differential expression features in the omics datasets are involved in different pathways, these features are found in some key signaling pathways, such as inflammatory response and interstitial fibrosis regulation [26,142]. Moreover, according to the findings of a functional analysis conducted by Yingdie Zhang et al., 18 m6A-mediated mRNAs were observed to regulate pathways that were closely related to “phagosomes”, “antigen processing and presentation”, and “apoptosis”. This suggests that m6A methylation plays an essential role in the development of silicosis [143]. Multiomics analysis can help identify potential drug targets for the treatment of silicosis fibrosis, and this approach can be used to design and develop targeted drugs. However, these targets are still in the preclinical stage and require further clinical experimental studies before they can be applied in the treatment of silicosis.
In the future, the utilization of novel omics techniques in the study of silicosis, including the integration of omics and spatomics, may establish a more comprehensive and objective approach, significantly enhancing our capacity to explore the pathology and molecular mechanisms of silicosis and offering a prospect for discovering effective diagnostic and therapeutic interventions for silicosis. Nonetheless, it is imperative to subject the targets identified by this method to rigorous testing and validation before implementing them in clinical practice. It is essential to note that the pathological mechanisms of silicosis are intricate, and the biological effects of cytokines produced by inflammatory cells interact in a complex manner, leading to the formation of a network of cells. Consequently, blocking a single target may not suffice to alter the degree of inflammation and fibrosis and effectively treat the disease. Thus, a comprehensive understanding of the pathological mechanisms of silicosis is indispensable.
3.4. Clinical Drugs of Silicosis and IPF
To date, there is no specific remedy for silicosis. The efficacious treatments available for patients with silicosis are WLL and lung transplantation. We should not only attach importance to basic research, but also accelerate the process of clinical trials of basic research results.
3.4.1. Silicosis
Management of silicosis consists of using bronchodilators and cough medication. However, symptomatic treatment may only ameliorate symptoms rather than restoring health. Most drugs that have shown positive effects in animal models, especially reducing lung fibrosis, have not been yet translated into clinically approved drugs in many countries, including Europe and the USA [144]. Tetrandrine stands as the sole drug sanctioned for silicosis treatment in China, as per the approval of regulatory authorities [145]. Recent clinical trials have demonstrated that the combined administration of tetrandrine with other drugs yields more pronounced therapeutic outcomes than conventional treatment modalities [94]. Despite the minimal adverse effects of tetrandrine therapy for pulmonary fibrosis, it remains incapable of reversing fibrosis and curing patients afflicted with silicosis.
3.4.2. IPF
Since many of the signaling pathways of silicosis and IPF overlap, it is possible that clinical medications used in the treatment of IPF could hold promise in the treatment of silicosis. For instance, antifibrotic drugs like pirfenidone and nintedanib, which are commonly prescribed for IPF [146], have demonstrated efficacy in reducing lung inflammation and fibrotic changes in animal models of silicosis [147,148,149]. Due to the significant side effects of nintedanib, it has not been an option for silicosis treatment, but recently an experiment engineered a nanocrystal-based suspension formulation of nintedanib possessing specific physicochemical properties to enhance drug retention in the lung for localized treatment of silicosis [150], bringing new hope to nintedanib for treating silicosis and highlighting that overcoming the side effects of existing clinical drugs can provide a new direction for the treatment of silicosis. Therefore, insights gained from antifibrosis drugs utilized in IPF could offer potential avenues for treating silicosis.
While these drug studies may only serve to delay the progression or alleviate the clinical symptoms of silicosis, they nevertheless provide a glimmer of hope for a potential cure. The treatment of silicosis is a protracted process that necessitates persistent basic research, comprehensive exploration of pathogenesis, implementation of advanced biological science technology, and other related approaches. In this manner, we can discover more drugs that can be utilized in clinical practice, thereby increasing the likelihood of curing silicosis.
4. Conclusions
The treatment of silicosis fibrosis has been a challenging topic due to the differences between animal models and human bodies and other difficulties, and there is currently no effective treatment available. In recent years, scholars have made some progress in the treatment of silicosis pulmonary fibrosis, which has brought hope for further exploration of the therapeutic effect of drugs and the clinical treatment of silicosis. However, the mechanism of action and side effects of some drugs for silicosis pulmonary fibrosis remain unclear, which limits their clinical application. The direction of future research may involve delving into innovative approaches to tackling silicosis through the implementation of combination administration or multitarget therapy, supplemented by state-of-the-art biological science technology. Additionally, drawing inspiration from comparable diseases like IPF may prove to be a valuable avenue of exploration. This approach can help to bridge the gap between basic research and clinical application and shorten the distance between research and practice for silicosis.
Author Contributions
Conceptualization, R.L. and S.C.; writing—original draft preparation, R.L. and S.C.; writing—review and editing, H.K. and S.C.; visualization, R.L. and S.C.; supervision, R.L. and S.C.; project administration, R.L. and S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by grants from the National Natural Science Foundation of China (No.82173493), the Health Commission Program of Hunan Province (No.D202312019523), and the Hunan Normal University undergraduate innovative experiment project and entrepreneurship program (No.2019116).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Ac-SDKP | N-acetyl-seryl-aspartyl-lysyl-proline |
| AEC | Alveolar epithelial cell |
| AKT | Protein kinase B |
| AM | Alveolar macrophage |
| ARE | Antioxidant-responsive element |
| ASK-1 | Arabidopsis Serine/Threonine Kinase1 |
| BALF | Broncho alveolar lavage fluid |
| Bax | BCL2-Associated X |
| BCL2 | B-cell lymphoma-2 |
| BLM | Bleomycin |
| BMP | Bone morphogenetic protein |
| Col | Collagen |
| COX | Cyclooxygenase |
| CTGF | Connective tissue growth factor |
| CXCL | C-X-C motif chemokine ligand |
| ECM | Extracellular Matrix |
| EMT | Epithelial-Mesenchymal Transition |
| ERK | Extracellular signal-regulated kinase |
| Fas | Factor-related Apoptosis |
| FNs | Fullerene nanoparticles |
| FOXO3 | Forkhead box O3 |
| GPx | Glutathione peroxidase |
| IL | Interleukins |
| iNOS | Inducible nitric oxide synthase |
| IPF | Idiopathic pulmonary fibrosis |
| JAK | Janus kinase |
| JNK | The c-Jun NH(2)-terminal kinases |
| Keap1 | Kelch-like ECH-associated protein 1 |
| LC3 | Microtubule-associated protein 1A/1B-light chain 3 |
| LPCAT1 | Lysophosphatidylcholine acyltransferase 1 |
| MAPK | Mitogen-activated protein kinase |
| MDA | Malondialdehyde |
| MMP | Metallomatous proteases |
| MSCs | Mesenchymal stem cells |
| mTOR | Mammalian target of rapamycin |
| NAC | N-Acetyl-L-cysteine |
| NALP3 | Neutrophil Alkaline Phosphatase 3 |
| NF-κB | Nuclear factor kappa-B |
| NLRP3 | NOD-like receptor thermal protein domain associated protein 3 |
| NOX | NADPH oxidase |
| Nrf2 | NF-E2 p45-related factor 2 |
| PDGF | Platelet-derived growth factor |
| PI3K | Phosphatidylinositol 3-kinase |
| PPAR-γ | Peroxisome proliferator-activated receptor gamma |
| ROS | Reactive oxygen species |
| SNPs | Single nucleotide polymorphisms |
| SOD | Superoxide dismutase |
| STAT | Signal transducer and activator of transcription |
| Tan IIA | Tanshinone IIA |
| TCM | Traditional Chinese medicine |
| TGF-β | Transforming growth factor beta |
| TIMPs | Tissue inhibitor of metalloproteinases |
| TLR4 | Toll-like receptor 4 |
| TNF | Tumor necrosis factor |
| WLL | Whole lung lavage |
| Wnt | Wingless/Integrated |
| α-SMA | Alpha-smooth muscle actin |
References
- Barnes, H.; Goh, N.S.L.; Leong, T.L.; Hoy, R. Silica-associated lung disease: An old-world exposure in modern industries. Respirology 2019, 24, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Hessel, P.A.; Gamble, J.F.; Gee, J.B.; Gibbs, G.; Green, F.H.; Morgan, W.K.; Mossman, B.T. Silica, silicosis, and lung cancer: A response to a recent working group report. J. Occup. Environ. Med. 2000, 42, 704–720. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Remy-Jardin, M.; Myers, J.L.; Richeldi, L.; Ryerson, C.J.; Lederer, D.J.; Behr, J.; Cottin, V.; Danoff, S.K.; Morell, F.; et al. Diagnosis of Idiopathic Pulmonary Fibrosis. An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline. Am. J. Respir. Crit. Care Med. 2018, 198, e44–e68. [Google Scholar] [CrossRef] [PubMed]
- Hoy, R.F.; Jeebhay, M.F.; Cavalin, C.; Chen, W.; Cohen, R.A.; Fireman, E.; Go, L.H.T.; Leon-Jimenez, A.; Menendez-Navarro, A.; Ribeiro, M.; et al. Current global perspectives on silicosis-Convergence of old and newly emergent hazards. Respirology 2022, 27, 387–398. [Google Scholar] [CrossRef]
- León-Jiménez, A.; Mánuel, J.M.; García-Rojo, M.; Pintado-Herrera, M.G.; López-López, J.A.; Hidalgo-Molina, A.; García, R.; Muriel-Cueto, P.; Maira-González, N.; Del Castillo-Otero, D.; et al. Compositional and structural analysis of engineered stones and inorganic particles in silicotic nodules of exposed workers. Part. Fibre. Toxicol. 2021, 18, 41. [Google Scholar] [CrossRef]
- Leon-Jimenez, A.; Hidalgo-Molina, A.; Conde-Sanchez, M.A.; Perez-Alonso, A.; Morales-Morales, J.M.; Garcia-Gamez, E.M.; Cordoba-Dona, J.A. Artificial Stone Silicosis: Rapid Progression Following Exposure Cessation. Chest 2020, 158, 1060–1068. [Google Scholar] [CrossRef]
- Leso, V.; Fontana, L.; Romano, R.; Gervetti, P.; Iavicoli, I. Artificial Stone Associated Silicosis: A Systematic Review. Int. J. Environ. Res. Public Health 2019, 16, 568. [Google Scholar] [CrossRef]
- Marques Da Silva, V.; Benjdir, M.; Montagne, P.; Pairon, J.C.; Lanone, S.; Andujar, P. Pulmonary Toxicity of Silica Linked to Its Micro- or Nanometric Particle Size and Crystal Structure: A Review. Nanomaterials 2022, 12, 2392. [Google Scholar] [CrossRef]
- Honnons, S.; Porcher, J.M. In vivo experimental model for silicosis. J. Environ. Pathol. Toxicol. Oncol. 2000, 19, 391–400. [Google Scholar]
- Jin, F.; Li, Y.; Wang, X.; Yang, X.; Li, T.; Xu, H.; Wei, Z.; Liu, H. Effect of Sex Differences in Silicotic Mice. Int. J. Mol. Sci. 2022, 23, 14203. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Tian, H.T.; Liu, H.; Xie, R. The role of macrophage-derived TGF-β1 on SiO(2)-induced pulmonary fibrosis: A review. Toxicol. Ind. Health 2021, 37, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Chen, S. Macrophage Autophagy and Silicosis: Current Perspective and Latest Insights. Int. J. Mol. Sci. 2021, 22, 453. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Ma, J.; Yang, M.; Fan, L.; Chen, W. Extracellular signal-regulated kinase signaling pathway and silicosis. Toxicol. Res. 2021, 10, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Sayan, M.; Mossman, B.T. The NLRP3 inflammasome in pathogenic particle and fibre-associated lung inflammation and diseases. Part. Fibre Toxicol. 2016, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Xaubet, A.; Serrano-Mollar, A.; Ancochea, J. Pirfenidone for the treatment of idiopathic pulmonary fibrosis. Expert Opin. Pharmacother. 2014, 15, 275–281. [Google Scholar] [CrossRef]
- Raghu, G.; Remy-Jardin, M.; Richeldi, L.; Thomson, C.C.; Inoue, Y.; Johkoh, T.; Kreuter, M.; Lynch, D.A.; Maher, T.M.; Martinez, F.J.; et al. Idiopathic Pulmonary Fibrosis (an Update) and Progressive Pulmonary Fibrosis in Adults: An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline. Am. J. Respir. Crit. Care Med. 2022, 205, e18–e47. [Google Scholar] [CrossRef]
- Spagnolo, P.; Cottin, V. Genetics of idiopathic pulmonary fibrosis: From mechanistic pathways to personalised medicine. J. Med. Genet. 2017, 54, 93–99. [Google Scholar] [CrossRef]
- Enomoto, N.; Suda, T.; Kono, M.; Kaida, Y.; Hashimoto, D.; Fujisawa, T.; Inui, N.; Nakamura, Y.; Imokawa, S.; Funai, K.; et al. Amount of elastic fibers predicts prognosis of idiopathic pulmonary fibrosis. Respir. Med. 2013, 107, 1608–1616. [Google Scholar] [CrossRef]
- Sgalla, G.; Iovene, B.; Calvello, M.; Ori, M.; Varone, F.; Richeldi, L. Idiopathic pulmonary fibrosis: Pathogenesis and management. Respir. Res. 2018, 19, 32. [Google Scholar] [CrossRef]
- Mei, Q.; Liu, Z.; Zuo, H.; Yang, Z.; Qu, J. Idiopathic Pulmonary Fibrosis: An Update on Pathogenesis. Front. Pharmacol. 2021, 12, 797292. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Xu, H.; Liu, H. Early Identification, Accurate Diagnosis, and Treatment of Silicosis. Can. Respir. J. 2022, 2022, 3769134. [Google Scholar] [CrossRef]
- Luppi, F.; Kalluri, M.; Faverio, P.; Kreuter, M.; Ferrara, G. Idiopathic pulmonary fibrosis beyond the lung: Understanding disease mechanisms to improve diagnosis and management. Respir. Res. 2021, 22, 109. [Google Scholar] [CrossRef]
- Samarelli, A.V.; Masciale, V.; Aramini, B.; Coló, G.P.; Tonelli, R.; Marchioni, A.; Bruzzi, G.; Gozzi, F.; Andrisani, D.; Castaniere, I.; et al. Molecular Mechanisms and Cellular Contribution from Lung Fibrosis to Lung Cancer Development. Int. J. Mol. Sci. 2021, 22, 12179. [Google Scholar] [CrossRef] [PubMed]
- Effendi, W.I.; Nagano, T. The Crucial Role of NLRP3 Inflammasome in Viral Infection-Associated Fibrosing Interstitial Lung Diseases. Int. J. Mol. Sci. 2021, 22, 10447. [Google Scholar] [CrossRef] [PubMed]
- Adamcakova, J.; Mokra, D. New Insights into Pathomechanisms and Treatment Possibilities for Lung Silicosis. Int. J. Mol. Sci. 2021, 22, 4162. [Google Scholar] [CrossRef] [PubMed]
- Bo, C.; Zhang, J.; Sai, L.; Du, Z.; Yu, G.; Li, C.; Li, M.; Peng, C.; Jia, Q.; Shao, H. Integrative transcriptomic and proteomic analysis reveals mechanisms of silica-induced pulmonary fibrosis in rats. BMC Pulm. Med. 2022, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, P.; Kropski, J.A.; Jones, M.G.; Lee, J.S.; Rossi, G.; Karampitsakos, T.; Maher, T.M.; Tzouvelekis, A.; Ryerson, C.J. Idiopathic pulmonary fibrosis: Disease mechanisms and drug development. Pharmacol. Ther. 2021, 222, 107798. [Google Scholar] [CrossRef]
- Matsubayashi, S.; Ito, S.; Araya, J.; Kuwano, K. Drugs against metabolic diseases as potential senotherapeutics for aging-related respiratory diseases. Front. Endocrinol. 2023, 14, 1079626. [Google Scholar] [CrossRef]
- McDonough, J.E.; Martens, D.S.; Tanabe, N.; Ahangari, F.; Verleden, S.E.; Maes, K.; Verleden, G.M.; Kaminski, N.; Hogg, J.C.; Nawrot, T.S.; et al. A role for telomere length and chromosomal damage in idiopathic pulmonary fibrosis. Respir. Res. 2018, 19, 132. [Google Scholar] [CrossRef]
- Fan, Y.; Zheng, C.; Wu, N.; Li, Y.; Huang, X.; Ye, Q. Telomerase gene variants and telomere shortening in patients with silicosis or asbestosis. Occup. Environ. Med. 2020, 78, 342–348. [Google Scholar] [CrossRef]
- Yin, H.; Xie, Y.; Gu, P.; Li, W.; Zhang, Y.; Yao, Y.; Chen, W.; Ma, J. The emerging role of epigenetic regulation in the progression of silicosis. Clin. Epigenetics 2022, 14, 169. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Bernard, K.; Tang, X.; Hu, M.; Horowitz, J.C.; Thannickal, V.J.; Sanders, Y.Y. Glutaminolysis Epigenetically Regulates Antiapoptotic Gene Expression in Idiopathic Pulmonary Fibrosis Fibroblasts. Am. J. Respir. Cell Mol. Biol. 2019, 60, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Dees, C.; Chakraborty, D.; Distler, J.H.W. Cellular and molecular mechanisms in fibrosis. Exp. Dermatol. 2021, 30, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Moss, B.J.; Ryter, S.W.; Rosas, I.O. Pathogenic Mechanisms Underlying Idiopathic Pulmonary Fibrosis. Annu. Rev. Pathol. 2022, 17, 515–546. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Y.; Qiu, T.; Liu, W.; Yao, P. Autophagy, an important therapeutic target for pulmonary fibrosis diseases. Clin. Chim. Acta Int. J. Clin. Chem. 2020, 502, 139–147. [Google Scholar] [CrossRef]
- Li, S.Y.; Li, C.; Zhang, Y.T.; He, X.; Chen, X.; Zeng, X.N.; Liu, F.W.; Chen, Y.; Chen, J. Targeting Mechanics-Induced Fibroblast Activation through CD44-RhoA-YAP Pathway Ameliorates Crystalline Silica-Induced Silicosis. Theranostics 2019, 9, 4993–5008. [Google Scholar] [CrossRef]
- Cottin, V.; Crestani, B.; Valeyre, D.; Wallaert, B.; Cadranel, J.; Dalphin, J.C.; Delaval, P.; Israel-Biet, D.; Kessler, R.; Reynaud-Gaubert, M.; et al. Diagnosis and management of idiopathic pulmonary fibrosis: French practical guidelines. Eur. Respir. Rev. 2014, 23, 193–214. [Google Scholar] [CrossRef]
- Krefft, S.; Wolff, J.; Rose, C. Silicosis: An Update and Guide for Clinicians. Clin. Chest Med. 2020, 41, 709–722. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.J.; Kim, S.; Kim, Y.J.; Kim, H.C. Epidemiology and comorbidities in idiopathic pulmonary fibrosis: A nationwide cohort study. BMC Pulm. Med. 2023, 23, 54. [Google Scholar] [CrossRef]
- Ge, C.; Peters, S.; Olsson, A.; Portengen, L.; Schüz, J.; Almansa, J.; Behrens, T.; Pesch, B.; Kendzia, B.; Ahrens, W.; et al. Respirable Crystalline Silica Exposure, Smoking, and Lung Cancer Subtype Risks. A Pooled Analysis of Case-Control Studies. Am. J. Respir. Crit. Care Med. 2020, 202, 412–421. [Google Scholar] [CrossRef]
- Sato, T.; Shimosato, T.; Klinman, D.M. Silicosis and lung cancer: Current perspectives. Lung Cancer 2018, 9, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Abu Qubo, A.; Numan, J.; Snijder, J.; Padilla, M.; Austin, J.H.M.; Capaccione, K.M.; Pernia, M.; Bustamante, J.; O’Connor, T.; Salvatore, M.M. Idiopathic pulmonary fibrosis and lung cancer: Future directions and challenges. Breathe 2022, 18, 220147. [Google Scholar] [CrossRef] [PubMed]
- Cottin, V.; Spagnolo, P.; Bonniaud, P.; Nolin, M.; Dalon, F.; Kirchgässler, K.U.; Kamath, T.V.; Van Ganse, E.; Belhassen, M. Mortality and Respiratory-Related Hospitalizations in Idiopathic Pulmonary Fibrosis Not Treated With Antifibrotics. Front. Med. 2021, 8, 802989. [Google Scholar] [CrossRef] [PubMed]
- McEwen, K.; Brodie, L. Lung transplantation for silicosis and recovery: An Australian case study. Br. J. Nurs 2021, 30, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Q.; Yao, R.; Guo, N. Experimental studies on the therapeutic effects of lung lavage with large volume of saline on silicosis. J. Hyg. Res. 1997, 26, 77–79. [Google Scholar]
- Rosengarten, D.; Fox, B.D.; Fireman, E.; Blanc, P.D.; Rusanov, V.; Fruchter, O.; Raviv, Y.; Shtraichman, O.; Saute, M.; Kramer, M.R. Survival following lung transplantation for artificial stone silicosis relative to idiopathic pulmonary fibrosis. Am. J. Ind. Med. 2017, 60, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Traver, G.; Mont, S.; Gius, D.; Lawson, W.E.; Ding, G.X.; Sekhar, K.R.; Freeman, M.L. Loss of Nrf2 promotes alveolar type 2 cell loss in irradiated, fibrotic lung. Free. Radic. Biol. Med. 2017, 112, 578–586. [Google Scholar] [CrossRef]
- Du, S.; Li, C.; Lu, Y.; Lei, X.; Zhang, Y.; Li, S.; Liu, F.; Chen, Y.; Weng, D.; Chen, J. Dioscin Alleviates Crystalline Silica-Induced Pulmonary Inflammation and Fibrosis through Promoting Alveolar Macrophage Autophagy. Theranostics 2019, 9, 1878–1892. [Google Scholar] [CrossRef]
- Yuan, L.; Sun, Y.; Zhou, N.; Wu, W.; Zheng, W.; Wang, Y. Dihydroquercetin Attenuates Silica-Induced Pulmonary Fibrosis by Inhibiting Ferroptosis Signaling Pathway. Front. Pharmacol. 2022, 13, 845600. [Google Scholar] [CrossRef]
- Liu, N.; Cao, F.; Li, Q.; Zhang, Y.; Zhang, Z.; Guan, W. Study of quercetin on pulmonary fibrosis by silica particles. J. Hyg. Res. 2014, 43, 814–818. [Google Scholar]
- Peng, H.B.; Wang, R.X.; Deng, H.J.; Wang, Y.H.; Tang, J.D.; Cao, F.Y.; Wang, J.H. Protective effects of oleanolic acid on oxidative stress and the expression of cytokines and collagen by the AKT/NF-κB pathway in silicotic rats. Mol. Med. Rep. 2017, 15, 3121–3128. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Xu, M.; Zhao, L.; Zhang, H.; Li, J.; Jin, F.; Li, Y.; Li, T.; Yang, X.; Li, S.; et al. Quercetin Alleviates Pulmonary Fibrosis in Mice Exposed to Silica by Inhibiting Macrophage Senescence. Front. Pharmacol. 2022, 13, 912029. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Shi, Z.Z.; Shu, L.F.; Wang, J.; Li, M.Y.; Ding, Z.S.; Jiang, F.S. Research on the Anti-Pulmonary Fibrosis Effect of the Bletilla striata Polysaccharide in Rat Silicosis Model. J. Chin. Med. Mater. 2016, 39, 1638–1642. [Google Scholar]
- Zhu, Z.; Li, Q.; Xu, C.; Zhao, J.; Li, S.; Wang, Y.; Tian, L. Sodium tanshinone IIA sulfonate attenuates silica-induced pulmonary fibrosis in rats via activation of the Nrf2 and thioredoxin system. Environ. Toxicol. Pharm. 2020, 80, 103461. [Google Scholar] [CrossRef]
- Feng, F.; Li, N.; Cheng, P.; Zhang, H.; Wang, H.; Wang, Y.; Wang, W. Tanshinone IIA attenuates silica-induced pulmonary fibrosis via inhibition of TGF-beta1-Smad signaling pathway. Biomed. Pharmacother. 2020, 121, 109586. [Google Scholar] [CrossRef]
- Feng, F.; Cheng, P.; Xu, S.; Li, N.; Wang, H.; Zhang, Y.; Wang, W. Tanshinone IIA attenuates silica-induced pulmonary fibrosis via Nrf2-mediated inhibition of EMT and TGF-β1/Smad signaling. Chem. -Biol. Interact. 2020, 319, 109024. [Google Scholar] [CrossRef]
- Yang, J.; Wang, T.; Li, Y.; Yao, W.; Ji, X.; Wu, Q.; Han, L.; Han, R.; Yan, W.; Yuan, J.; et al. Earthworm extract attenuates silica-induced pulmonary fibrosis through Nrf2-dependent mechanisms. Lab. Investig. 2016, 96, 1279–1300. [Google Scholar] [CrossRef]
- Pang, X.; Shao, L.; Nie, X.; Yan, H.; Li, C.; Yeo, A.J.; Lavin, M.F.; Xia, Q.; Shao, H.; Yu, G.; et al. Emodin attenuates silica-induced lung injury by inhibition of inflammation, apoptosis and epithelial-mesenchymal transition. Int. Immunopharmacol. 2021, 91, 107277. [Google Scholar] [CrossRef]
- Zhu, Y.; Chai, Y.; Xiao, G.; Liu, Y.; Xie, X.; Xiao, W.; Zhou, P.; Ma, W.; Zhang, C.; Li, L. Astragalus and its formulas as a therapeutic option for fibrotic diseases: Pharmacology and mechanisms. Front. Pharmacol. 2022, 13, 1040350. [Google Scholar] [CrossRef]
- Hu, A.; Chen, W.; Huang, Y. Advances in the Pathogenesis, Prevention, and Treatment of Transfusion-related Acute Lung Injury. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. Acta Acad. Med. Sin. 2020, 42, 674–680. [Google Scholar] [CrossRef]
- Tang, M.; Yang, Z.; Liu, J.; Zhang, X.; Guan, L.; Liu, X.; Zeng, M. Combined intervention with N-acetylcysteine and desipramine alleviated silicosis development by regulating the Nrf2/HO-1 and ASMase/ceramide signaling pathways. Ecotoxicol. Environ. Saf. 2022, 242, 113914. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Shao, L.; Fang, J.; Zhang, J.; Chen, Y.; Yeo, A.J.; Lavin, M.F.; Yu, G.; Shao, H. Hesperetin attenuates silica-induced lung injury by reducing oxidative damage and inflammatory response. Exp. Ther. Med. 2021, 21, 297. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, D.; Li, X.; Guan, M.; Zhou, Y.; Li, L.; Jia, W.; Zhou, C.; Shu, C.; Wang, C.; et al. Fullerene nanoparticles: A promising candidate for the alleviation of silicosis-associated pulmonary inflammation. Nanoscale 2020, 12, 17470–17479. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, M.; Giardina, C.; Hubbard, A.K. Silica-induced caspase activation in mouse alveolar macrophages is dependent upon mitochondrial integrity and aspartic proteolysis. Toxicol. Sci. 2003, 76, 91–101. [Google Scholar] [CrossRef]
- Gozal, E.; Ortiz, L.A.; Zou, X.; Burow, M.E.; Lasky, J.A.; Friedman, M. Silica-induced apoptosis in murine macrophage: Involvement of tumor necrosis factor-alpha and nuclear factor-kappaB activation. Am. J. Respir. Cell Mol. Biol. 2002, 27, 91–98. [Google Scholar] [CrossRef]
- Yao, S.Q.; He, Q.C.; Yuan, J.X.; Chen, J.; Chen, G.; Lu, Y.; Bai, Y.P.; Zhang, C.M.; Yuan, Y.; Xu, Y.J. Role of Fas/FasL pathway-mediated alveolar macrophages releasing inflammatory cytokines in human silicosis. Biomed. Environ. Sci. 2013, 26, 930–933. [Google Scholar] [CrossRef]
- Wang, L.; Bowman, L.; Lu, Y.; Rojanasakul, Y.; Mercer, R.R.; Castranova, V.; Ding, M. Essential role of p53 in silica-induced apoptosis. Am. J. Physiology. Lung Cell. Mol. Physiol. 2005, 288, L488–L496. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Zhao, J.; Su, K.; He, K.; Da, Y.; Xia, H. Endoplasmic Reticulum Stress Regulates Cardiomyocyte Apoptosis in Myocardial Fibrosis Development via PERK-Mediated Autophagy. Cardiovasc. Toxicol. 2020, 20, 618–626. [Google Scholar] [CrossRef]
- Lawrence, J.; Nho, R. The Role of the Mammalian Target of Rapamycin (mTOR) in Pulmonary Fibrosis. Int. J. Mol. Sci. 2018, 19, 778. [Google Scholar] [CrossRef]
- Hu, M.; Yang, J.; Xu, Y. Isoorientin suppresses sepsis-induced acute lung injury in mice by activating an EPCR-dependent JAK2/STAT3 pathway. J. Mol. Histol. 2022, 53, 97–109. [Google Scholar] [CrossRef]
- Fan, M.; Xiao, H.; Song, D.; Zhu, L.; Zhang, J.; Zhang, X.; Wang, J.; Dai, H.; Wang, C. A Novel N-Arylpyridone Compound Alleviates the Inflammatory and Fibrotic Reaction of Silicosis by Inhibiting the ASK1-p38 Pathway and Regulating Macrophage Polarization. Front. Pharmacol. 2022, 13, 848435. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tang, K.; Hu, P.; Tan, S.; Yang, S.; Yang, C.; Chen, G.; Luo, Y.; Zou, H. Atractylenolide III alleviates the apoptosis through inhibition of autophagy by the mTOR-dependent pathway in alveolar macrophages of human silicosis. Mol. Cell. Biochem. 2021, 476, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, H.; Cao, Z.; Gu, J.; Pei, L.; Jia, M.; Su, M. Kaempferol Modulates Autophagy and Alleviates Silica-Induced Pulmonary Fibrosis. DNA Cell Biol. 2019, 38, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.J.; Liu, Y.; Zhang, Z.; Yang, P.R.; Li, Z.G.; Song, M.Y.; Qi, X.M.; Han, Z.F.; Pang, J.L.; Li, B.C.; et al. Pirfenidone ameliorates silica-induced lung inflammation and fibrosis in mice by inhibiting the secretion of interleukin-17A. Acta Pharmacol. Sin. 2022, 43, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, R.R.; Elkashef, W.F.; Said, E. Tadalafil reduces airway hyperactivity and protects against lung and respiratory airways dysfunction in a rat model of silicosis. Int. Immunopharmacol. 2016, 40, 530–541. [Google Scholar] [CrossRef]
- Han, J.; Jia, Y.; Wang, S.; Gan, X. The Improvement Effect of Sodium Ferulate on the Formation of Pulmonary Fibrosis in Silicosis Mice Through the Neutrophil Alkaline Phosphatase 3 (NALP3)/Transforming Growth Factor-β1 (TGF-β1)/α-Smooth Muscle Actin (α-SMA) Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2021, 27, e927978. [Google Scholar] [CrossRef]
- Yoldas, O.; Karaca, T.; Bilgin, B.C.; Yilmaz, O.H.; Simsek, G.G.; Alici, I.O.; Uzdogan, A.; Karaca, N.; Akin, T.; Yoldas, S.; et al. Tamoxifen citrate: A glimmer of hope for silicosis. J. Surg. Res. 2015, 193, 429–434. [Google Scholar] [CrossRef]
- Ge, A.; Ma, Y.; Liu, Y.N.; Li, Y.S.; Gu, H.; Zhang, J.X.; Wang, Q.X.; Zeng, X.N.; Huang, M. Diosmetin prevents TGF-β1-induced epithelial-mesenchymal transition via ROS/MAPK signaling pathways. Life Sci. 2016, 153, 1–8. [Google Scholar] [CrossRef]
- Li, N.; Feng, F.; Wu, K.; Zhang, H.; Zhang, W.; Wang, W. Inhibitory effects of astragaloside IV on silica-induced pulmonary fibrosis via inactivating TGF-β1/Smad3 signaling. Biomed. Pharmacother. 2019, 119, 109387. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Y.; Zhu, Z.; Yang, G.; An, G.; Li, X.; Niu, P.; Chen, L.; Tian, L. BMP-7 attenuated silica-induced pulmonary fibrosis through modulation of the balance between TGF-β/Smad and BMP-7/Smad signaling pathway. Chem. Biol. Interact. 2016, 243, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.F.; Wei, Y.L.; Wang, X.; Wang, L.N.; Li, X. Buyang Huanwu Tang inhibits cellular epithelial-to-mesenchymal transition by inhibiting TGF-β1 activation of PI3K/Akt signaling pathway in pulmonary fibrosis model in vitro. BMC Complement. Med. Ther. 2020, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.J.; He, X.Y.; Wang, W.X.; Liang, J.; Zhang, Y.D.; Liang, J.T.; Chen, D.Y. Dahuang Zhechong Pills Suppress Silicosis Fibrosis Progression via p38 MAPK/TGF-β1/Smad Pathway In Vitro. Evid. -Based Complement. Altern. Med. 2021, 2021, 6662261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, X.; Hecker, L.; Kurundkar, D.; Kurundkar, A.; Liu, H.; Jin, T.H.; Desai, L.; Bernard, K.; Thannickal, V.J. Inhibition of mechanosensitive signaling in myofibroblasts ameliorates experimental pulmonary fibrosis. J. Clin. Investig. 2013, 123, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dai, W.; Wang, Y.; Gu, Q.; Yang, D.; Zhang, M. Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice. Int. J. Environ. Res. Public Health 2015, 12, 10739–10754. [Google Scholar] [CrossRef]
- Fan, L.H.; Liu, T.F.; Guo, M.; Liu, M.L.; Wang, Z.P.; Si, S.J. Effect of schisandrin B on lung mRNA expression of transforming growth factor-beta1 signal transduction molecule in rat lungs exposed to silica. Chin. J. Ind. Hyg. Occup. Dis. 2011, 29, 255–259. [Google Scholar]
- Li, Y.; Meng, W.; Zhao, X. Mechanism of Gallus domesticus extract reducing pulmonary interstitial fibrosis in pneumoconiosis rats by regulating the autophagy level of alveolar macrophages. J. Clin. Exp. Med. 2021, 20, 792–796. [Google Scholar]
- Cruz, F.F.; Horta, L.F.; Maia Lde, A.; Lopes-Pacheco, M.; da Silva, A.B.; Morales, M.M.; Gonçalves-de-Albuquerque, C.F.; Takiya, C.M.; de Castro-Faria-Neto, H.C.; Rocco, P.R. Dasatinib Reduces Lung Inflammation and Fibrosis in Acute Experimental Silicosis. PLoS ONE 2016, 11, e0147005. [Google Scholar] [CrossRef]
- Cheng, D.; Xu, Q.; Wang, Y.; Li, G.; Sun, W.; Ma, D.; Zhou, S.; Liu, Y.; Han, L.; Ni, C. Metformin attenuates silica-induced pulmonary fibrosis via AMPK signaling. J. Transl. Med. 2021, 19, 349. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Hao, C.; Li, Y.; Guo, Y.; Si, H.; He, J.; Deng, M.; Niu, Z.; Wang, C.; Xu, X.; et al. Lysophosphatidylcholine acyltransferase 1 alleviates silica-induced pulmonary fibrosis by modulating lipid metabolism. Biomed. Pharmacother. 2022, 155, 113638. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.M.; Fang, Z.H.; Yao, Y. Therapeutic efficacy of tetrandrine tablets combined with matrine injection in treatment of silicosis. Chin. J. Ind. Hyg. Occup. Dis. 2012, 30, 778–780. [Google Scholar]
- Guo, X.; Qi, J.; Li, H.; Xing, Z. Clinical efficacy of acetylcysteine combined with tetrandrine tablets on patients with silicosis and its effect on exercise tolerance and pulmonary function. Exp. Ther. Med. 2020, 20, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Singh, R. Protective effects of intranasal curcumin on silica-induced lung damage. Cytokine 2022, 157, 155949. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Xiao, T.; Yang, J.H.; Qin, Y.; Gao, J.J.; Liu, H.J.; Zhou, H.G. Parthenolide attenuated bleomycin-induced pulmonary fibrosis via the NF-κB/Snail signaling pathway. Respir. Res. 2018, 19, 111. [Google Scholar] [CrossRef]
- Zhou, Z.; Kandhare, A.D.; Kandhare, A.A.; Bodhankar, S.L. Hesperidin ameliorates bleomycin-induced experimental pulmonary fibrosis via inhibition of TGF-beta1/Smad3/AMPK and IkappaBalpha/NF-kappaB pathways. EXCLI J. 2019, 18, 723–745. [Google Scholar] [CrossRef]
- Xiong, Y.; Cui, X.; Zhou, Y.; Chai, G.; Jiang, X.; Ge, G.; Wang, Y.; Sun, H.; Che, H.; Nie, Y.; et al. Dehydrocostus lactone inhibits BLM-induced pulmonary fibrosis and inflammation in mice via the JNK and p38 MAPK-mediated NF-kappaB signaling pathways. Int. Immunopharmacol. 2021, 98, 107780. [Google Scholar] [CrossRef]
- Liang, W.; Greven, J.; Fragoulis, A.; Horst, K.; Bläsius, F.; Wruck, C.; Pufe, T.; Kobbe, P.; Hildebrand, F.; Lichte, P. Sulforaphane-Dependent Up-Regulation of NRF2 Activity Alleviates Both Systemic Inflammatory Response and Lung Injury After Hemorrhagic Shock/Resuscitation in Mice. Shock 2022, 57, 221–229. [Google Scholar] [CrossRef]
- Lan, Y.J.; Cheng, M.H.; Ji, H.M.; Bi, Y.Q.; Han, Y.Y.; Yang, C.Y.; Gu, X.; Gao, J.; Dong, H.L. Melatonin ameliorates bleomycin-induced pulmonary fibrosis via activating NRF2 and inhibiting galectin-3 expression. Acta Pharmacol. Sin. 2022, 44, 1029–1037. [Google Scholar] [CrossRef]
- Pan, L.; Lu, Y.; Li, Z.; Tan, Y.; Yang, H.; Ruan, P.; Li, R. Ginkgo biloba Extract EGb761 Attenuates Bleomycin-Induced Experimental Pulmonary Fibrosis in Mice by Regulating the Balance of M1/M2 Macrophages and Nuclear Factor Kappa B (NF-κB)-Mediated Cellular Apoptosis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e922634. [Google Scholar] [CrossRef] [PubMed]
- Kyung, S.Y.; Kim, D.Y.; Yoon, J.Y.; Son, E.S.; Kim, Y.J.; Park, J.W.; Jeong, S.H. Sulforaphane attenuates pulmonary fibrosis by inhibiting the epithelial-mesenchymal transition. BMC Pharmacol. Toxicol. 2018, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.X.; Qiu, J.; Zhou, H.H.; Yu, Y.; Zhou, D.L.; Xu, Y.; Zhu, M.Z.; Ge, X.P.; Li, J.M.; Lv, C.J.; et al. Dihydroartemisinin alleviates oxidative stress in bleomycin-induced pulmonary fibrosis. Life Sci. 2018, 205, 176–183. [Google Scholar] [CrossRef]
- Qian, W.; Cai, X.; Qian, Q.; Zhang, W.; Wang, D. Astragaloside IV modulates TGF-β1-dependent epithelial-mesenchymal transition in bleomycin-induced pulmonary fibrosis. J. Cell. Mol. Med. 2018, 22, 4354–4365. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, D.; Xu, L.; Zhao, P.; Deng, Z.; Mo, X.; Li, P.; Qi, L.; Li, J.; Gao, J. Total extract of Yupingfeng attenuates bleomycin-induced pulmonary fibrosis in rats. Phytomedicine: Int. J. Phytother. Phytopharm. 2015, 22, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Zhao, F.; Sang, X.; Qiao, Y.; Shao, R.; Wang, Y.; Gao, S.; Fan, G.; Zhu, Y.; Yang, J. Combination therapy of tanshinone IIA and puerarin for pulmonary fibrosis via targeting IL6-JAK2-STAT3/STAT1 signaling pathways. Phytother. Res. 2021, 35, 5883–5898. [Google Scholar] [CrossRef]
- Dong, S.H.; Liu, Y.W.; Wei, F.; Tan, H.Z.; Han, Z.D. Asiatic acid ameliorates pulmonary fibrosis induced by bleomycin (BLM) via suppressing pro-fibrotic and inflammatory signaling pathways. Biomed. Pharmacother. 2017, 89, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhao, P.; Xie, Z.; Wang, L.; Chen, S. Oridonin Inhibits Myofibroblast Differentiation and Bleomycin-induced Pulmonary Fibrosis by Regulating Transforming Growth Factor β (TGFβ)/Smad Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 7548–7555. [Google Scholar] [CrossRef]
- Sun, S.C.; Han, R.; Hou, S.S.; Yi, H.Q.; Chi, S.J.; Zhang, A.H. Juglanin alleviates bleomycin-induced lung injury by suppressing inflammation and fibrosis via targeting sting signaling. Biomed. Pharmacother. 2020, 127, 110119. [Google Scholar] [CrossRef]
- Zhang, K.; Si, X.P.; Huang, J.; Han, J.; Liang, X.; Xu, X.B.; Wang, Y.T.; Li, G.Y.; Wang, H.Y.; Wang, J.H. Preventive Effects of Rhodiola rosea L. on Bleomycin-Induced Pulmonary Fibrosis in Rats. Int. J. Mol. Sci. 2016, 17, 879. [Google Scholar] [CrossRef]
- Rajasekar, N.; Sivanantham, A.; Kar, A.; Mahapatra, S.K.; Ahirwar, R.; Thimmulappa, R.K.; Paramasivam, S.G.; Subbiah, R. Tannic acid alleviates experimental pulmonary fibrosis in mice by inhibiting inflammatory response and fibrotic process. Inflammopharmacology 2020, 28, 1301–1314. [Google Scholar] [CrossRef] [PubMed]
- Prashanth Goud, M.; Bale, S.; Pulivendala, G.; Godugu, C. Therapeutic effects of Nimbolide, an autophagy regulator, in ameliorating pulmonary fibrosis through attenuation of TGF-β1 driven epithelial-to-mesenchymal transition. Int. Immunopharmacol. 2019, 75, 105755. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhou, Y.; Yu, J.; Du, Y.; Tan, Y.; Ke, Y.; Wang, J.; Han, B.; Ge, F. Ophiocordyceps lanpingensis polysaccharides attenuate pulmonary fibrosis in mice. Biomed. Pharmacother. 2020, 126, 110058. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wen, H.T.; Gu, K.J.; Hu, X.D.; Yang, T.; Yan, X.F.; Ye, T.J.; Huo, J.L.; Hu, J. Total extract of Xin Jia Xuan Bai Cheng Qi decoction inhibits pulmonary fibrosis via the TGF-β/Smad signaling pathways in vivo and in vitro. Drug Des. Dev. Ther. 2019, 13, 2873–2886. [Google Scholar] [CrossRef]
- Chen, F.; Wang, P.L.; Fan, X.S.; Yu, J.H.; Zhu, Y.; Zhu, Z.H. Effect of Renshen Pingfei Decoction, a traditional Chinese prescription, on IPF induced by Bleomycin in rats and regulation of TGF-β1/Smad3. J. Ethnopharmacol. 2016, 186, 289–297. [Google Scholar] [CrossRef]
- Meng, L.; Zhang, X.; Wang, H.; Dong, H.; Gu, X.; Yu, X.; Liu, Y. Yangyin Yiqi Mixture Ameliorates Bleomycin-Induced Pulmonary Fibrosis in Rats through Inhibiting TGF-β1/Smad Pathway and Epithelial to Mesenchymal Transition. Evid. -Based Complement. Altern. Med. 2019, 2019, 2710509. [Google Scholar] [CrossRef]
- Liu, B.; Lü, W.; Ge, H.; Tang, H.; Li, R.; Zhang, C. Protective Effect of the Traditional Chinese Patent Medicine Qing-Xuan Granule against Bleomycin-Induced Pulmonary Fibrosis in Mice. Chem. Biodivers. 2019, 16, e1900467. [Google Scholar] [CrossRef]
- Li, H.; Wang, Z.; Zhang, J.; Wang, Y.; Yu, C.; Zhang, J.; Song, X.; Lv, C. Feifukang ameliorates pulmonary fibrosis by inhibiting JAK-STAT signaling pathway. BMC Complement. Altern. Med. 2018, 18, 234. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, L.F.; Zhao, Y.; Wei, L.M.; Guo, P.; Yu, M.; Cao, B.; Li, T.; Chen, H.; Zou, Z.M. The Chinese Herbal Medicine Formula mKG Suppresses Pulmonary Fibrosis of Mice Induced by Bleomycin. Int. J. Mol. Sci. 2016, 17, 238. [Google Scholar] [CrossRef]
- Yu, J.Z.; Ying, Y.; Liu, Y.; Sun, C.B.; Dai, C.; Zhao, S.; Tian, S.Z.; Peng, J.; Han, N.P.; Yuan, J.L.; et al. Antifibrotic action of Yifei Sanjie formula enhanced autophagy via PI3K-AKT-mTOR signaling pathway in mouse model of pulmonary fibrosis. Biomed. Pharmacother. 2019, 118, 109293. [Google Scholar] [CrossRef]
- Bai, Y.; Li, J.; Zhao, P.; Li, Y.; Li, M.; Feng, S.; Qin, Y.; Tian, Y.; Zhou, T. A Chinese Herbal Formula Ameliorates Pulmonary Fibrosis by Inhibiting Oxidative Stress via Upregulating Nrf2. Front. Pharmacol. 2018, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, X.; Wang, S.; Wang, W.; Guo, F.; Chen, Y.; Pan, B.; Zhang, M.; Fan, X. Combination of Salvia miltiorrhiza and ligustrazine attenuates bleomycin-induced pulmonary fibrosis in rats via modulating TNF-α and TGF-β. Chin. Med. 2018, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Q.; Wei, L.; Wang, Z.; Ma, W.; Liu, F.; Shen, Y.; Zhang, S.; Zhang, X.; Li, H.; et al. Dexamethasone combined with berberine is an effective therapy for bleomycin-induced pulmonary fibrosis in rats. Exp. Ther. Med. 2019, 18, 2385–2392. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Wang, J. Dioscin attenuates Bleomycin-Induced acute lung injury via inhibiting the inflammatory response in mice. Exp. Lung Res. 2019, 45, 236–244. [Google Scholar] [CrossRef]
- Veith, C.; Drent, M.; Bast, A.; van Schooten, F.J.; Boots, A.W. The disturbed redox-balance in pulmonary fibrosis is modulated by the plant flavonoid quercetin. Toxicol. Appl. Pharmacol. 2017, 336, 40–48. [Google Scholar] [CrossRef]
- Tian, S.L.; Yang, Y.; Liu, X.L.; Xu, Q.B. Emodin Attenuates Bleomycin-Induced Pulmonary Fibrosis via Anti-Inflammatory and Anti-Oxidative Activities in Rats. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 1. [Google Scholar] [CrossRef]
- Johnson, S.; Shaikh, S.B.; Muneesa, F.; Rashmi, B.; Bhandary, Y.P. Radiation induced apoptosis and pulmonary fibrosis: Curcumin an effective intervention? Int. J. Radiat. Biol. 2020, 96, 709–717. [Google Scholar] [CrossRef]
- Gao, J.; Li, C.; Wang, X.; Sun, X.; Zhang, R.; Chen, C.; Yu, M.; Liu, Y.; Zhu, Y.; Chen, J. Oridonin attenuates lung inflammation and fibrosis in silicosis via covalent targeting iNOS. Biomed. Pharmacother. 2022, 153, 113532. [Google Scholar] [CrossRef]
- Jia, L.; Sun, P.; Gao, H.; Shen, J.; Gao, Y.; Meng, C.; Fu, S.; Yao, H.; Zhang, G. Mangiferin attenuates bleomycin-induced pulmonary fibrosis in mice through inhibiting TLR4/p65 and TGF-beta1/Smad2/3 pathway. J. Pharm. Pharmacol. 2019, 71, 1017–1028. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Chen, S.; Cui, G.; Peng, C.; Lavin, M.F.; Sun, X.; Zhang, E.; Yang, Y.; Guan, Y.; Du, Z.; Shao, H. Transplantation of adipose-derived mesenchymal stem cells attenuates pulmonary fibrosis of silicosis via anti-inflammatory and anti-apoptosis effects in rats. Stem Cell Res. Ther. 2018, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, E.; Oliveira, H.; Silva, J.D.; Menna-Barreto, R.F.S.; Takyia, C.M.; Suk, J.S.; Witwer, K.W.; Paulaitis, M.E.; Hanes, J.; Rocco, P.R.M.; et al. Therapeutic effects of adipose-tissue-derived mesenchymal stromal cells and their extracellular vesicles in experimental silicosis. Respir. Res. 2018, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xu, D.; Li, Q.; Yang, Y.; Xu, H.; Wei, Z.; Wang, R.; Zhang, W.; Liu, Y.; Geng, Y.; et al. N-acetyl-seryl-aspartyl-lysyl-proline (Ac-SDKP) attenuates silicotic fibrosis by suppressing apoptosis of alveolar type II epithelial cells via mediation of endoplasmic reticulum stress. Toxicol. Appl. Pharmacol. 2018, 350, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Averyanov, A.; Koroleva, I.; Konoplyannikov, M.; Revkova, V.; Lesnyak, V.; Kalsin, V.; Danilevskaya, O.; Nikitin, A.; Sotnikova, A.; Kotova, S.; et al. First-in-human high-cumulative-dose stem cell therapy in idiopathic pulmonary fibrosis with rapid lung function decline. Stem Cells Transl. Med. 2020, 9, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Madl, C.M.; Heilshorn, S.C.; Blau, H.M. Bioengineering strategies to accelerate stem cell therapeutics. Nature 2018, 557, 335–342. [Google Scholar] [CrossRef]
- Chu, M.; Wu, S.; Wang, W.; Yu, Y.; Zhang, M.; Sang, L.; Tian, T.; Lu, Y.; Yuan, W.; Huang, Q.; et al. Functional variant of the carboxypeptidase M (CPM) gene may affect silica-related pneumoconiosis susceptibility by its expression: A multistage case-control study. Occup. Environ. Med. 2019, 76, 169–174. [Google Scholar] [CrossRef]
- Lv, J.; Xiao, J.; Jia, Q.; Meng, X.; Yang, Z.; Pu, S.; Li, M.; Yu, T.; Zhang, Y.; Wang, H.; et al. Identification of key pathways and genes in the progression of silicosis based on WGCNA. Inhal. Toxicol. 2022, 34, 304–318. [Google Scholar] [CrossRef]
- Wang, X.; Xu, K.; Yang, X.Y.; Liu, J.; Zeng, Q.; Wang, F.S. Upregulated miR-29c suppresses silica-induced lung fibrosis through the Wnt/beta-catenin pathway in mice. Hum. Exp. Toxicol. 2018, 37, 944–952. [Google Scholar] [CrossRef]
- Xu, T.; Yan, W.; Wu, Q.; Xu, Q.; Yuan, J.; Li, Y.; Li, P.; Pan, H.; Ni, C. MiR-326 Inhibits Inflammation and Promotes Autophagy in Silica-Induced Pulmonary Fibrosis through Targeting TNFSF14 and PTBP1. Chem. Res. Toxicol. 2019, 32, 2192–2203. [Google Scholar] [CrossRef]
- Chu, M.; Wu, S.; Wang, W.; Mao, L.; Yu, Y.; Jiang, L.; Yuan, W.; Zhang, M.; Sang, L.; Huang, Q.; et al. miRNA sequencing reveals miRNA-4508 from peripheral blood lymphocytes as potential diagnostic biomarker for silica-related pulmonary fibrosis: A multistage study. Respirology 2020, 25, 511–517. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, J.; Duan, Y.; Xu, H.; Cheng, Q.; Gao, X.; Li, S.; Yang, F.; Liu, H.; Yuan, J. Protein Expression Profile in Rat Silicosis Model Reveals Upregulation of PTPN2 and Its Inhibitory Effect on Epithelial-Mesenchymal Transition by Dephosphorylation of STAT3. Int. J. Mol. Sci. 2020, 21, 1189. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, Z.; Liu, J.; Song, M.; Zhang, T.; Chen, Y.; Hu, H.; Yang, P.; Li, B.; Song, X.; et al. Gefitinib and fostamatinib target EGFR and SYK to attenuate silicosis: A multi-omics study with drug exploration. Signal Transduct. Target. Ther. 2022, 7, 157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, P.; Xie, Y.; Fan, L.; You, X.; Yang, S.; Yao, Y.; Chen, W.; Ma, J. Insights into the mechanism underlying crystalline silica-induced pulmonary fibrosis via transcriptome-wide m(6)A methylation profile. Ecotoxicol. Environ. Saf. 2022, 247, 114215. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Pacheco, M.; Bandeira, E.; Morales, M.M. Cell-Based Therapy for Silicosis. Stem Cells Int. 2016, 2016, 5091838. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Wang, J.X.; Sun, Y.L.; Han, Z.F.; Zhou, Y.T.; Liu, Y.; Fan, T.H.; Li, Z.G.; Qi, X.M.; Luo, Y.; et al. Tetrandrine alleviates silicosis by inhibiting canonical and non-canonical NLRP3 inflammasome activation in lung macrophages. Acta Pharmacol. Sin. 2022, 43, 1274–1284. [Google Scholar] [CrossRef]
- Barratt, S.L.; Creamer, A.; Hayton, C.; Chaudhuri, N. Idiopathic Pulmonary Fibrosis (IPF): An Overview. J. Clin. Med. 2018, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Xing, C.; Li, M.; Jia, Q.; Bo, C.; Zhang, Z. Pirfenidone ameliorates pulmonary inflammation and fibrosis in a rat silicosis model by inhibiting macrophage polarization and JAK2/STAT3 signaling pathways. Ecotoxicol. Environ. Saf. 2022, 244, 114066. [Google Scholar] [CrossRef]
- Makino, S. Progressive fibrosing interstitial lung diseases: A new concept and indication of nintedanib. Mod. Rheumatol. 2021, 31, 13–19. [Google Scholar] [CrossRef]
- Sekihara, K.; Aokage, K.; Miyoshi, T.; Tane, K.; Ishii, G.; Tsuboi, M. Perioperative pirfenidone treatment as prophylaxis against acute exacerbation of idiopathic pulmonary fibrosis: A single-center analysis. Surg. Today 2020, 50, 905–911. [Google Scholar] [CrossRef]
- Andrade da Silva, L.H.; Vieira, J.B.; Cabral, M.R.; Antunes, M.A.; Lee, D.; Cruz, F.F.; Hanes, J.; Rocco, P.R.M.; Morales, M.M.; Suk, J.S. Development of nintedanib nanosuspension for inhaled treatment of experimental silicosis. Bioeng. Transl. Med. 2023, 8, e10401. [Google Scholar] [CrossRef]



































Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).