

Substances Secreted by Lactobacillus spp. from the Urinary Tract Microbiota Play a Protective Role against Proteus mirabilis Infections and Their Complications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
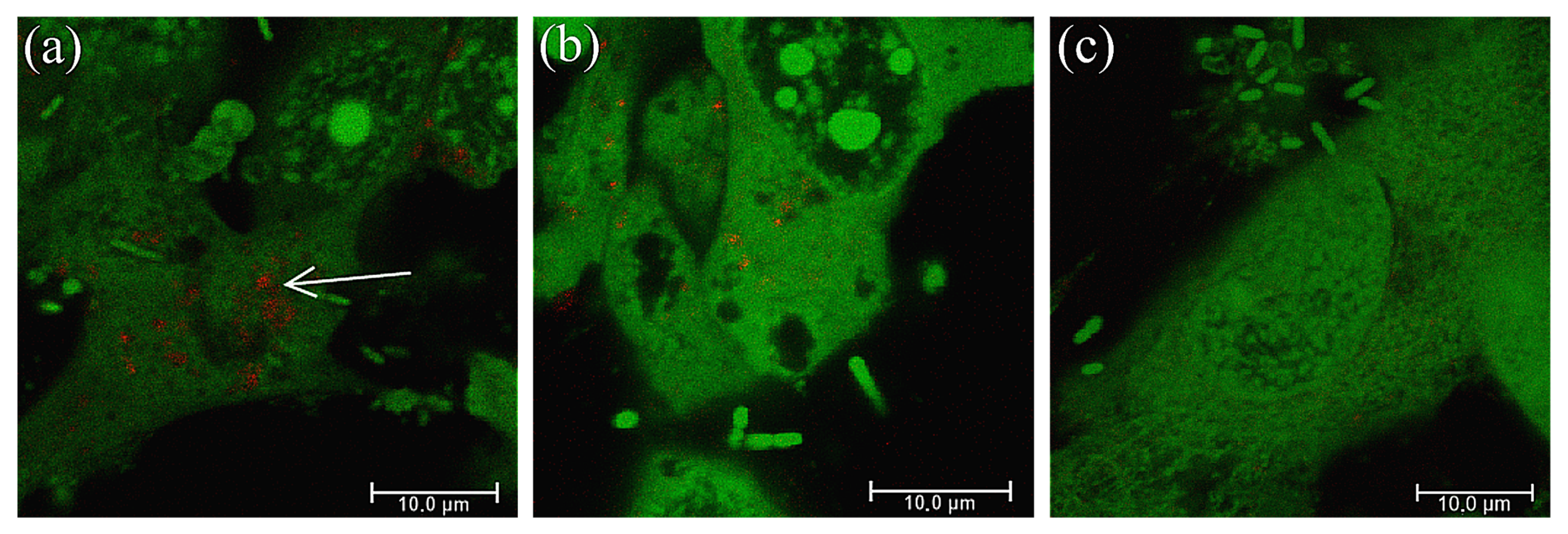
2.1. Extracellular Substances Secreted by Lactobacillus May Reduce the Adhesion of P. mirabilis Strains to the Bladder Epithelium In Vitro
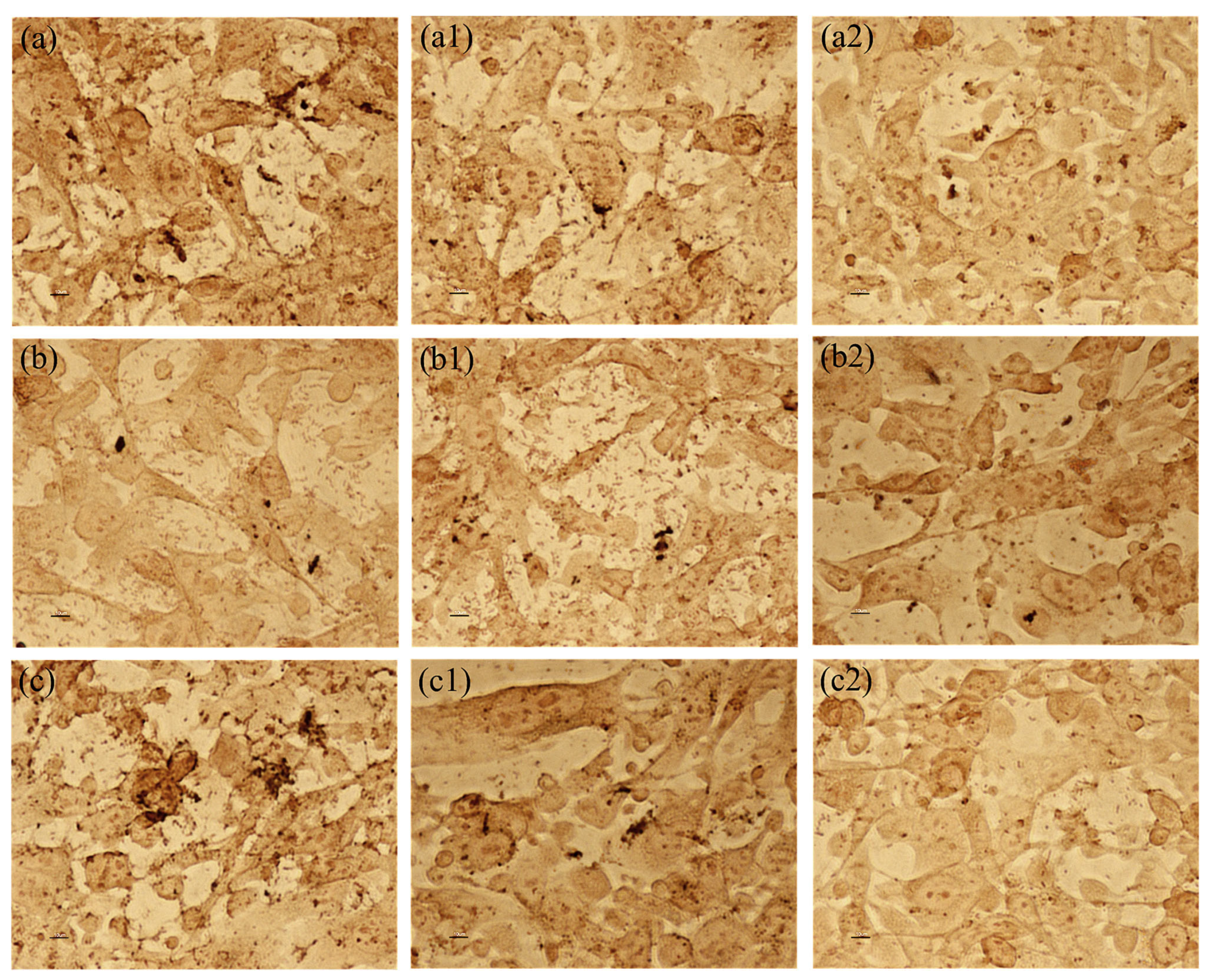
2.2. The Impact of the Substances Secreted by Lactobacillus on Cell Cytotoxicity Induced by P. mirabilis
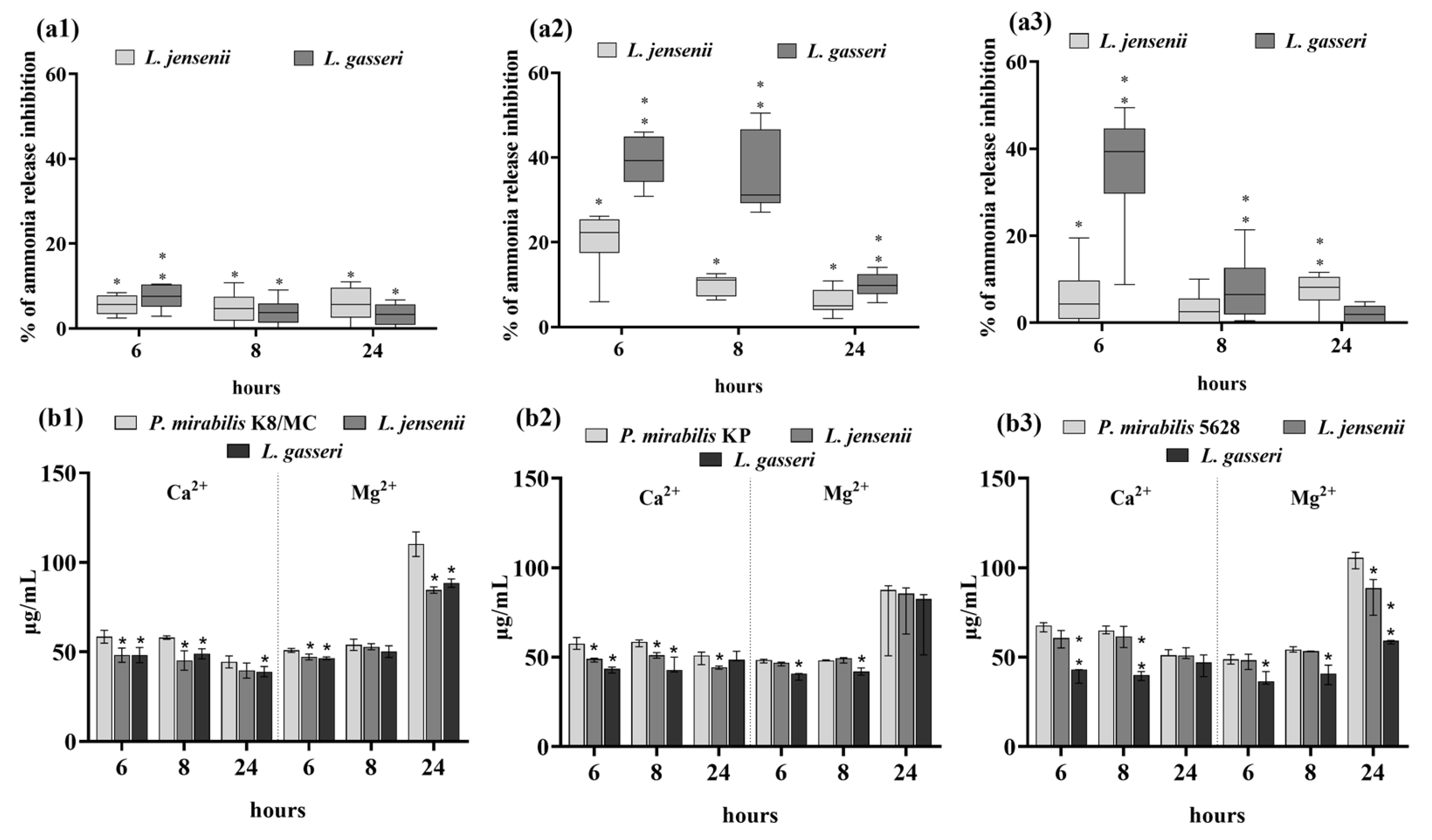
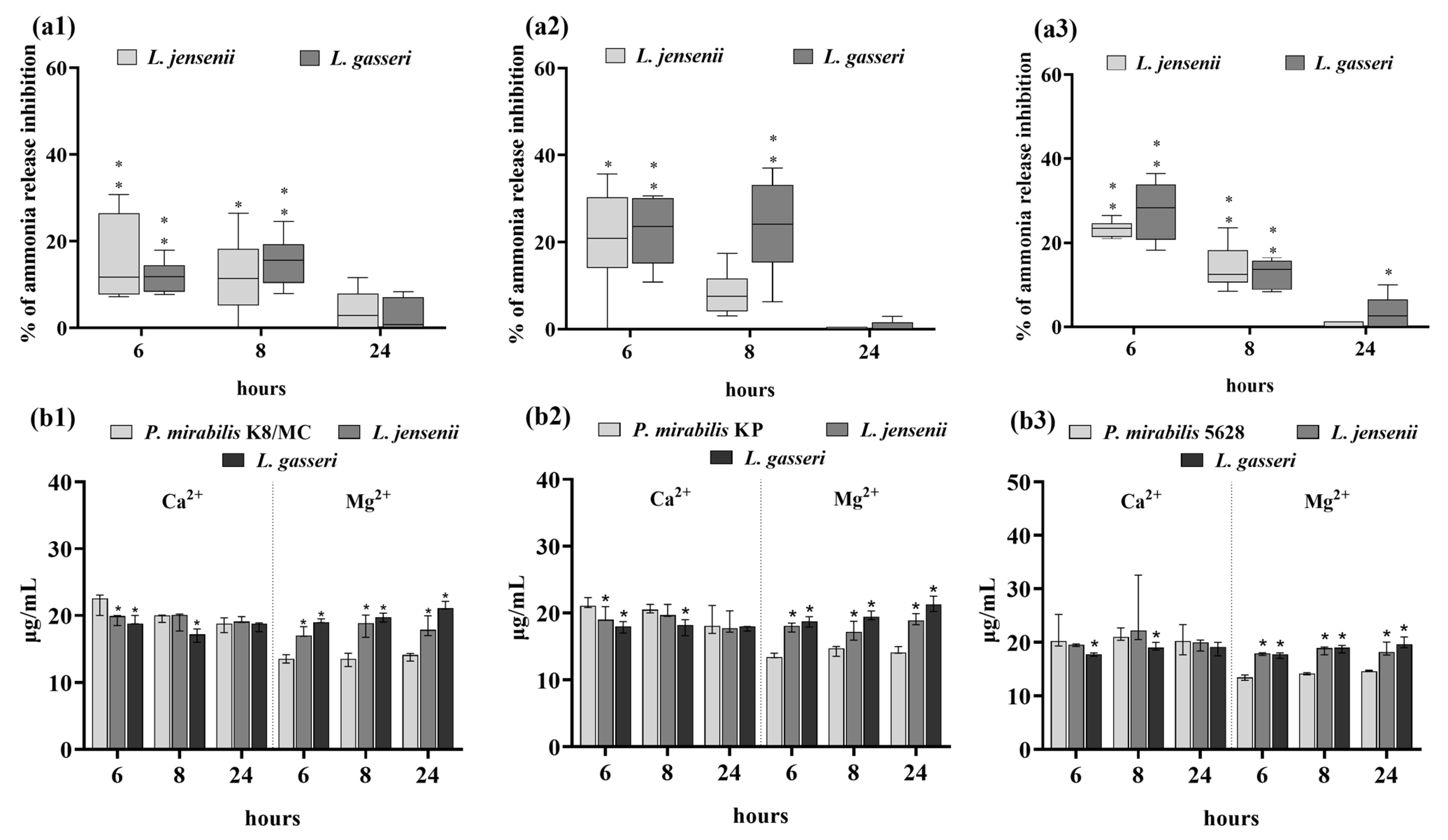
2.3. Extracellular Substances of Lactobacillus Inhibit the Crystallization of Urine Components Caused by P. mirabilis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Adhesion Assay
4.3. In Vitro Cytotoxicity Studies
4.4. Crystallization Assay
4.5. Imaging of Cell-Associated Crystallization
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armbruster, C.E.; Mobley, H.L.T. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.L.; Kamm, M.A.; Ng, S.C.; Morrison, M. Proteus spp. as Putative Gastrointestinal Pathogens. Clin. Microbiol. Rev. 2018, 31, e00085-17. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Huang, Z.; Yang, T.; Wang, G.; Li, P.; Yang, B.; Li, J. Pathogenesis of Proteus mirabilis in Catheter-Associated Urinary Tract Infections. Urol. Int. 2021, 105, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Nielubowicz, G.R.; Mobley, H.L.T. Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 2010, 7, 430–441. [Google Scholar] [CrossRef]
- Karki, N.; Leslie, S.W. Struvite and Triple Phosphate Renal Calculi. Available online: https://www.ncbi.nlm.nih.gov/books/NBK568783/ (accessed on 30 May 2023).
- Wang, Z.; Zhang, Y.; Zhang, J.; Deng, Q.; Liang, H. Recent advances on the mechanisms of kidney stone formation (Review). Int. J. Mol. Med. 2021, 48, 149. [Google Scholar] [CrossRef]
- Torzewska, A.; Budzyńska, A.; Białczak-Kokot, M.; Rózalski, A. In vitro studies of epithelium-associated crystallization caused by uropathogens during urinary calculi development. Microb. Pathog. 2014, 71–72, 25–31. [Google Scholar] [CrossRef]
- Marien, T.; Miller, N.L. Treatment of the Infected Stone. Urol. Clin. N. Am. 2015, 42, 459–472. [Google Scholar] [CrossRef]
- Türk, C.; Petřík, A.; Sarica, K.; Seitz, C.; Skolarikos, A.; Straub, M.; Knoll, T. EAU Guidelines on Interventional Treatment for Urolithiasis. Eur. Urol. 2016, 69, 475–482. [Google Scholar] [CrossRef]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—A role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef]
- Putonti, C.; Shapiro, J.W.; Ene, A.; Tsibere, O.; Wolfe, A.J. Comparative Genomic Study of Lactobacillus jensenii and the Newly Defined Lactobacillus mulieris Species Identifies Species-Specific Functionality. mSphere 2020, 5, e00560-20. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, A.; Pedro, N.A.; Salazar, S.B.; Mira, N.P. Effect of Acetic Acid and Lactic Acid at Low pH in Growth and Azole Resistance of Candida albicans and Candida glabrata. Front. Microbiol. 2019, 9, 3265. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli inactivate Chlamydia trachomatis through lactic acid but not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed]
- Leccese Terraf, M.C.; Juarez Tomás, M.S.; Rault, L.; Le Loir, Y.; Even, S.; Nader-Macías, M.E.F. In vitro effect of vaginal lactobacilli on the growth and adhesion abilities of uropathogenic Escherichia coli. Arch. Microbiol. 2017, 199, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Szczerbiec, D.; Piechocka, J.; Głowacki, R.; Torzewska, A. Organic Acids Secreted by Lactobacillus spp. Isolated from Urine and Their Antimicrobial Activity against Uropathogenic Proteus mirabilis. Molecules 2022, 27, 5557. [Google Scholar] [CrossRef]
- Szczerbiec, D.; Bednarska-Szczepaniak, K.; Torzewska, A. Department of Biology of Bacteria, Faculty of Biology and Environmental Protection, University of Lodz, Banacha 12/16, 90-237, Lodz, Poland. 2023; manuscript in revision. [Google Scholar]
- Espinosa-Ortiz, E.J.; Eisner, B.H.; Lange, D.; Gerlach, R. Current insights into the mechanisms and management of infection stones. Nat. Rev. Urol. 2019, 16, 35–53. [Google Scholar] [CrossRef]
- Flannigan, R.; Choy, W.H.; Chew, B.; Lange, D. Renal struvite stones—Pathogenesis, microbiology, and management strategies. Nat. Rev. Urol. 2014, 11, 333–341. [Google Scholar] [CrossRef]
- Wong, H.Y.; Riedl, C.R.; Griffith, D.P. Kindey Stones: Medical and Surgical Management; Co, F., Favus, M., Pak, C., Parks, J., Preminger, G., Eds.; Jaypee Brothers Medical Publishers: New Delhi, India, 1996; pp. 941–950. [Google Scholar]
- Gettman, M.T.; Segura, J.W. Struvite stones: Diagnosis and current treatment concepts. J. Endourol. 1999, 13, 653–658. [Google Scholar] [CrossRef]
- Bichler, K.H.; Eipper, E.; Naber, K.; Braun, V.; Zimmermann, R.; Lahme, S. Urinary infection stones. Int. J. Antimicrob. Agents 2002, 19, 488–498. [Google Scholar] [CrossRef]
- Kramer, G.; Klingler, H.C.; Steiner, G.E. Role of bacteria in the development of kidney stones. Curr. Opin. Urol. 2000, 10, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.D.; Hultgren, S.J. Urinary tract infections: Microbial pathogenesis, host-pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 2020, 18, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, Q.Z.; Wang, Y.Y.; Li, Y.L.; Zhou, G.T. Biomimetic synthesis of struvite with biogenic morphology and implication for pathological biomineralization. Sci. Rep. 2015, 5, 7718. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Niu, X.; Wang, B.; Na, R.; Xiao, B.; Yang, H. Evaluation of the Inhibitory Effects of Lactobacillus gasseri and Lactobacillus crispatus on the Adhesion of Seven Common Lower Genital Tract Infection-Causing Pathogens to Vaginal Epithelial Cells. Front. Med. 2020, 7, 284. [Google Scholar] [CrossRef]
- Johnson, J.A.; Delaney, L.F.; Ojha, V.; Rudraraju, M.; Hintze, K.R.; Siddiqui, N.Y.; Sysoeva, T.A. Commensal Urinary Lactobacilli Inhibit Major Uropathogens In Vitro with Heterogeneity at Species and Strain Level. Front. Cell Infect. Microbiol. 2022, 12, 870603. [Google Scholar] [CrossRef] [PubMed]
- Jayashree, S.; Karthikeyan, R.; Nithyalakshmi, S.; Ranjani, J.; Gunasekaran, P.; Rajendhran, J. Anti-adhesion property of the potential probiotic strain Lactobacillus fermentum 8711 against methicillin-resistant Staphylococcus aureus (MRSA). Front. Microbiol. 2018, 9, 411. [Google Scholar] [CrossRef]
- Maudsdotter, L.; Jonsson, H.; Roos, S.; Jonsson, A.B. Lactobacilli Reduce Cell Cytotoxicity Caused by Streptococcus pyogenes by Producing Lactic Acid That Degrades the Toxic Component Lipoteichoic Acid. Antimicrob. Agents Chemother. 2011, 55, 1622–1628. [Google Scholar] [CrossRef]
- Manzoor, M.A.P.; Mujeeburahiman, M.; Duwal, S.R.; Rekha, P.D. Investigation on growth and morphology of in vitro generated struvite crystals. Biocatal. Agric. Biotechnol. 2019, 17, 566–570. [Google Scholar] [CrossRef]
- Romani, A.M.P. Magnesium in health and disease. Met. Ions Life Sci. 2013, 13, 49–79. [Google Scholar] [CrossRef]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ sensing: Role in calcium homeostasis and signaling. Mol. Cell. 2017, 66, 780–788. [Google Scholar] [CrossRef]
- Mathoera, R.B.; Kok, D.J.; Verduin, C.M.; Nijman, R.J.M. Pathological and Therapeutic Significance of Cellular Invasion by Proteus mirabilis in an Enterocystoplasty Infection Stone Model. Infect. Immun. 2002, 70, 7022–7032. [Google Scholar] [CrossRef]
- Lin, S.-L.; Chung, C.-Y.; Chen, Z.-T.; Huang, C.-C.; Li, Y.-Z.; Huang, E.Y.-H.; Chiang, H.K. Observation and identification of autofluorescent urine crystals may be linked to a sign of urolithiasis. Urolithiasis 2022, 50, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Hütt, P.; Lapp, E.; Štšepetova, J.; Smidt, I.; Taelma, H.; Borovkova, N.; Oopkaup, H.; Ahelik, A.; Rööp, T.; Hoidmets, D.; et al. Characterisation of probiotic properties in human vaginal lactobacilli strains. Microb. Ecol. Health Dis. 2016, 27, 30484. [Google Scholar] [CrossRef]
- Punia Bangar, S.; Suri, S.; Trif, M.; Ozogul, F. Organic acids production from lactic acid bacteria: A preservation approach. Food Biosci. 2022, 46, 101615. [Google Scholar] [CrossRef]
- Pohanka, M. D-Lactic Acid as a Metabolite: Toxicology, Diagnosis, and Detection. Biomed Res. Int. 2020, 2020, 3419034. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Mendes-Soares, H.; Linhares, I.M.; Jayaram, A.; Ledger, W.J.; Forney, L.J. Influence of vaginal bacteria and D- and L-lactic acid isomers on vaginal extracellular matrix metalloproteinase inducer: Implications for protection against upper genital tract infections. mBio 2013, 4, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Griffith, D.P.; Musher, D.M.; Itin, C. Urease. The primary cause of infection-induced urinary stones. Investig. Urol. 1976, 13, 346–350. [Google Scholar]
- Toniolo, G.; Efthimiadou, E.K.; Kordas, G.; Chatgilialoglu, C. Development of multi-layered and multi-sensitive polymeric nanocontainers for cancer therapy: In vitro evaluation. Sci. Rep. 2018, 8, 14704. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-Hypochlorite Reaction for Determination of Ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Akiyoshi, T.; Ota, H.; Iijima, K.; Son, B.-K.; Kahyo, T.; Setou, M.; Ogawa, S.; Ouchi, Y.; Akishita, M. A novel organ culture model of aorta for vascular calcification. Atherosclerosis 2016, 244, 51–58. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczerbiec, D.; Słaba, M.; Torzewska, A. Substances Secreted by Lactobacillus spp. from the Urinary Tract Microbiota Play a Protective Role against Proteus mirabilis Infections and Their Complications. Int. J. Mol. Sci. 2024, 25, 103. https://doi.org/10.3390/ijms25010103
Szczerbiec D, Słaba M, Torzewska A. Substances Secreted by Lactobacillus spp. from the Urinary Tract Microbiota Play a Protective Role against Proteus mirabilis Infections and Their Complications. International Journal of Molecular Sciences. 2024; 25(1):103. https://doi.org/10.3390/ijms25010103
Chicago/Turabian StyleSzczerbiec, Dominika, Mirosława Słaba, and Agnieszka Torzewska. 2024. "Substances Secreted by Lactobacillus spp. from the Urinary Tract Microbiota Play a Protective Role against Proteus mirabilis Infections and Their Complications" International Journal of Molecular Sciences 25, no. 1: 103. https://doi.org/10.3390/ijms25010103