
Pin1 Downregulation Is Involved in Excess Retinoic Acid-Induced Failure of Neural Tube Closure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Atra Causes Brain Abnormalities in Mouse and Zebrafish Embryos
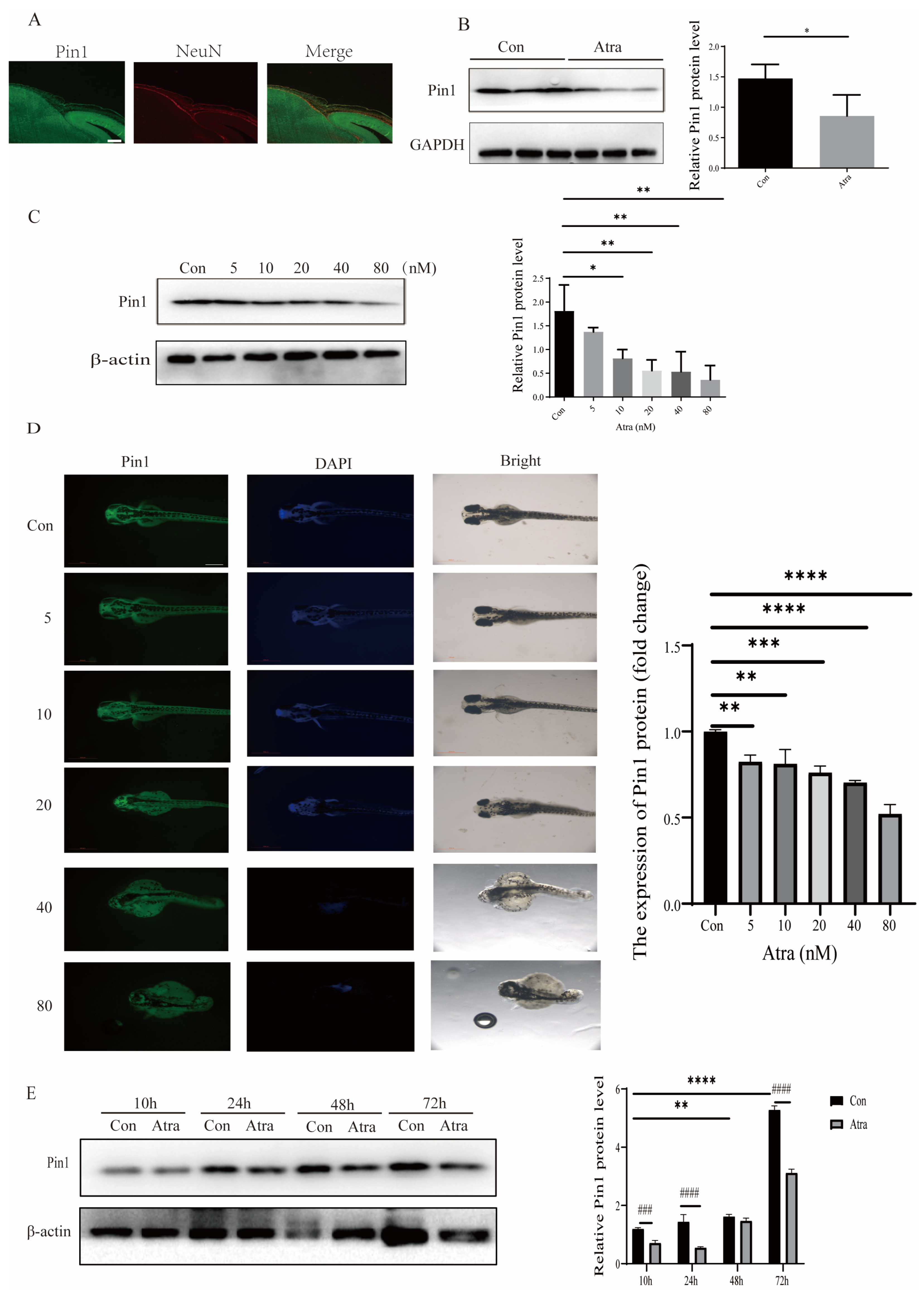
2.2. Pin1 Expression Was Reduced in Embryos and Neurons, Induced by RA and NTDs in Patients
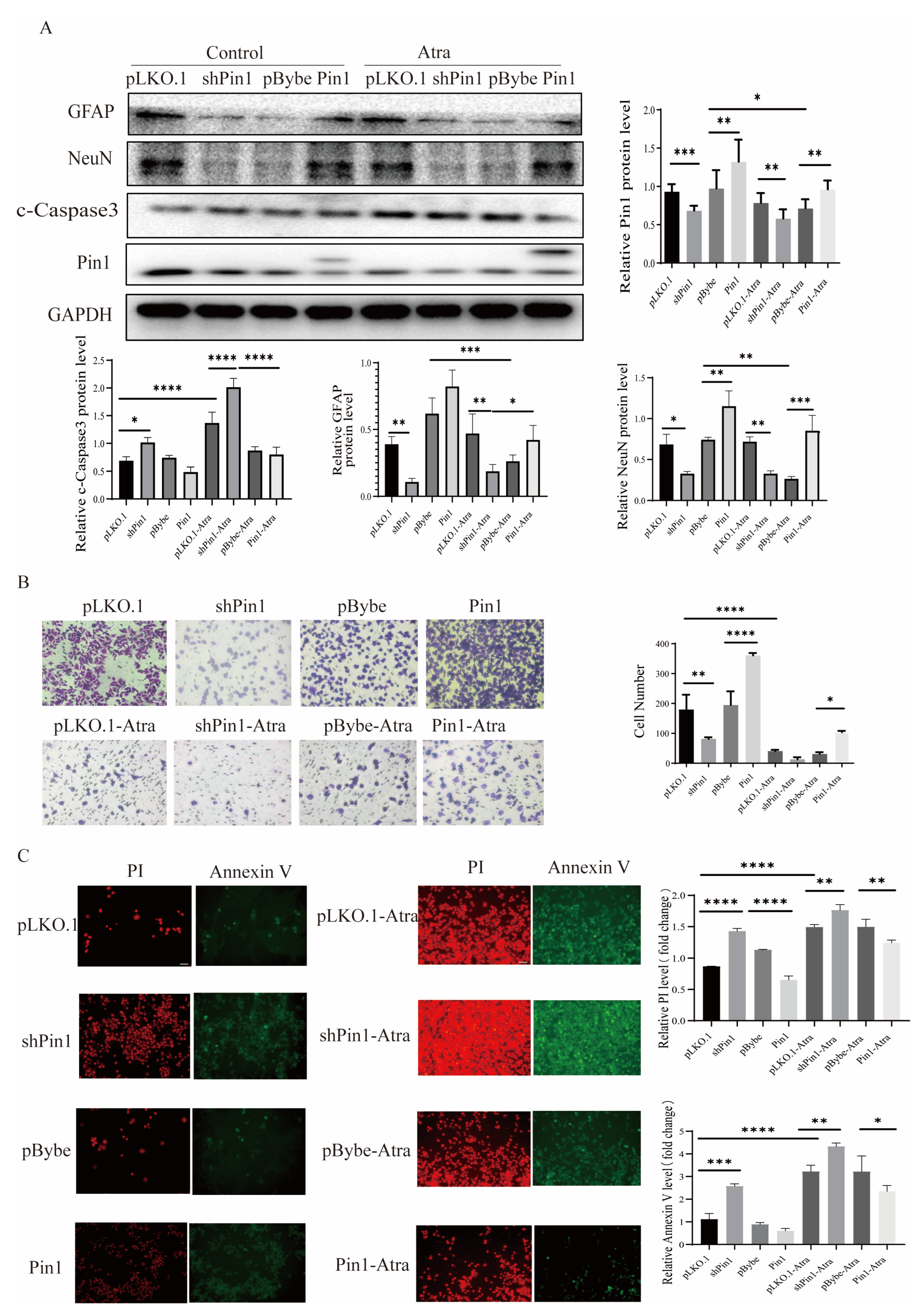
2.3. Pin1 Influenced the Apoptosis, Proliferation, Differentiation, and Migration of Neurons In Vitro
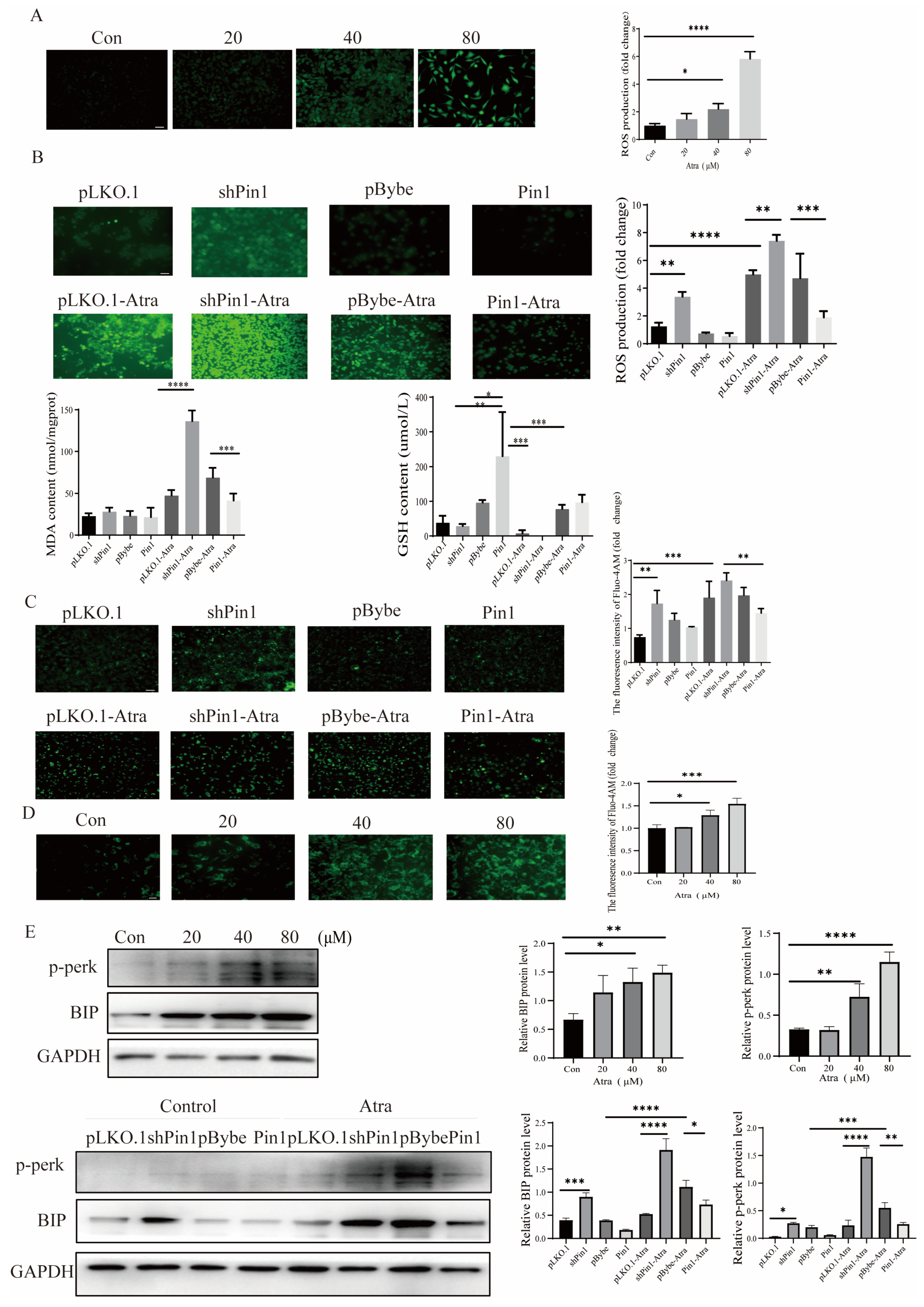
2.4. Pin1 Regulated ATRA-Induced OS and the ER in SHSY5Y Cells
2.5. The Knockdown of Pin1 Impacted the Proliferation of NSCs, as well as Oxidative Stress (OS) and Endoplasmic Reticulum Stress (ERs)
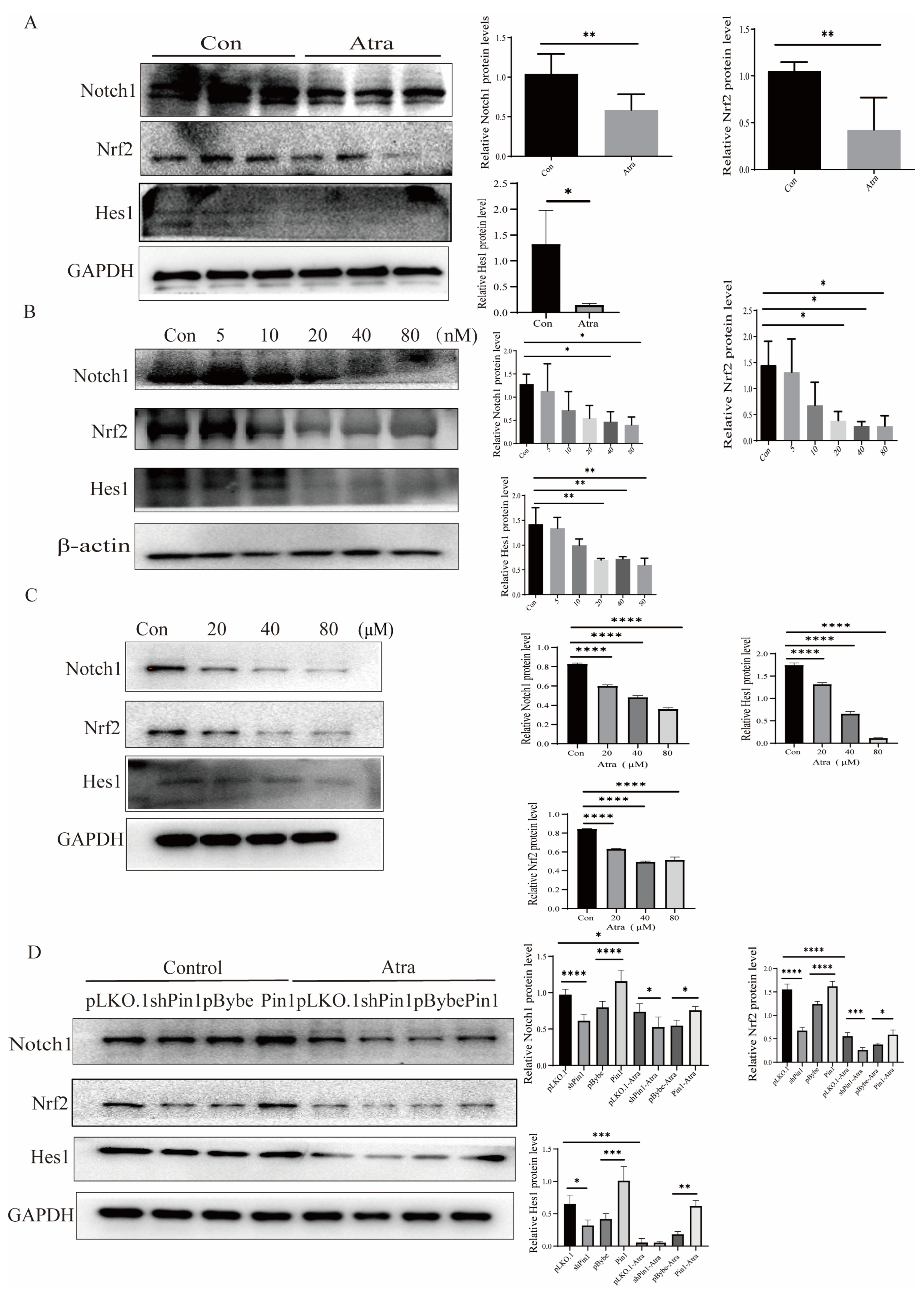
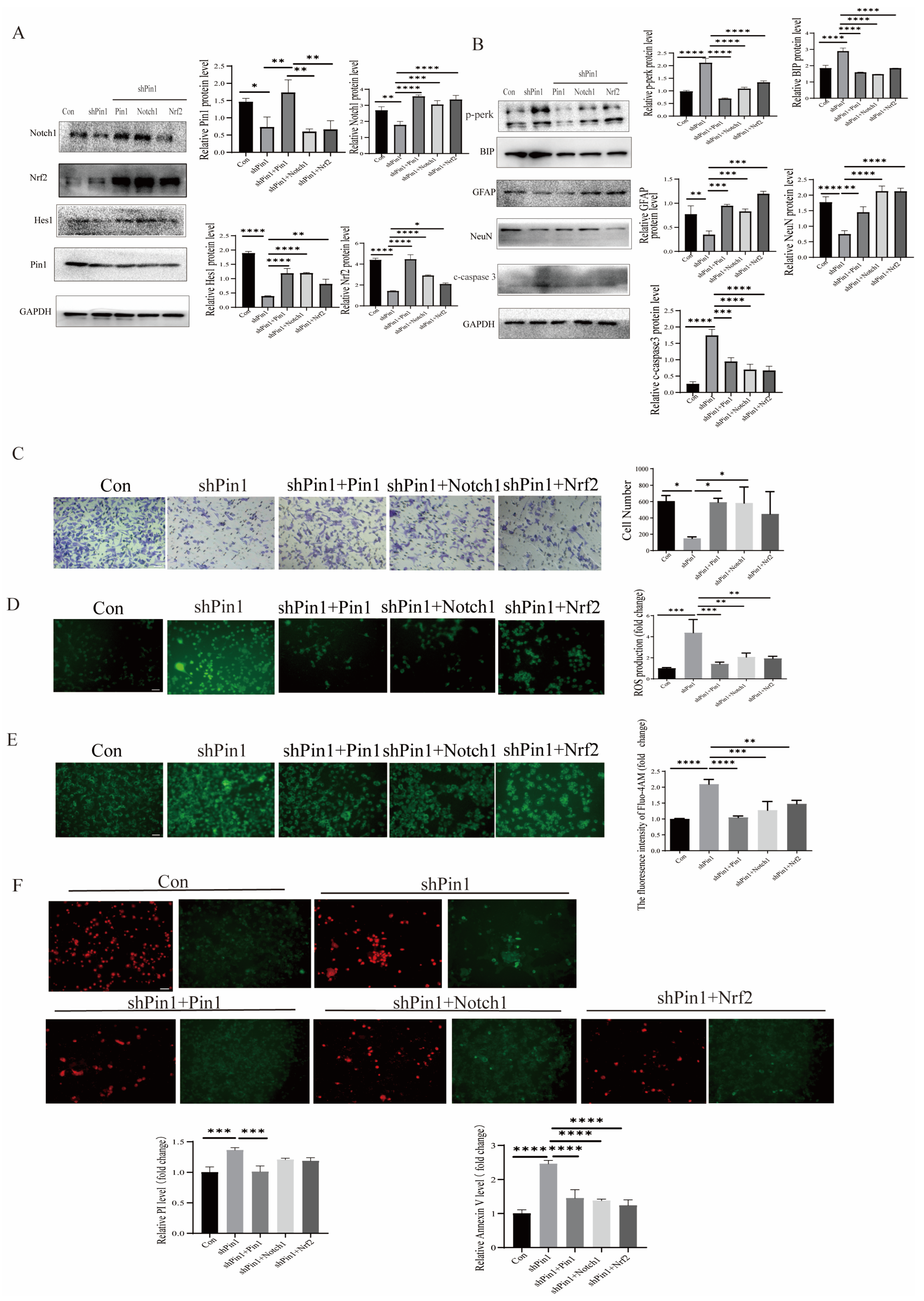
2.6. Pin1 Regulated the Notch1-Nrf2 Pathway in the Development of NTDs
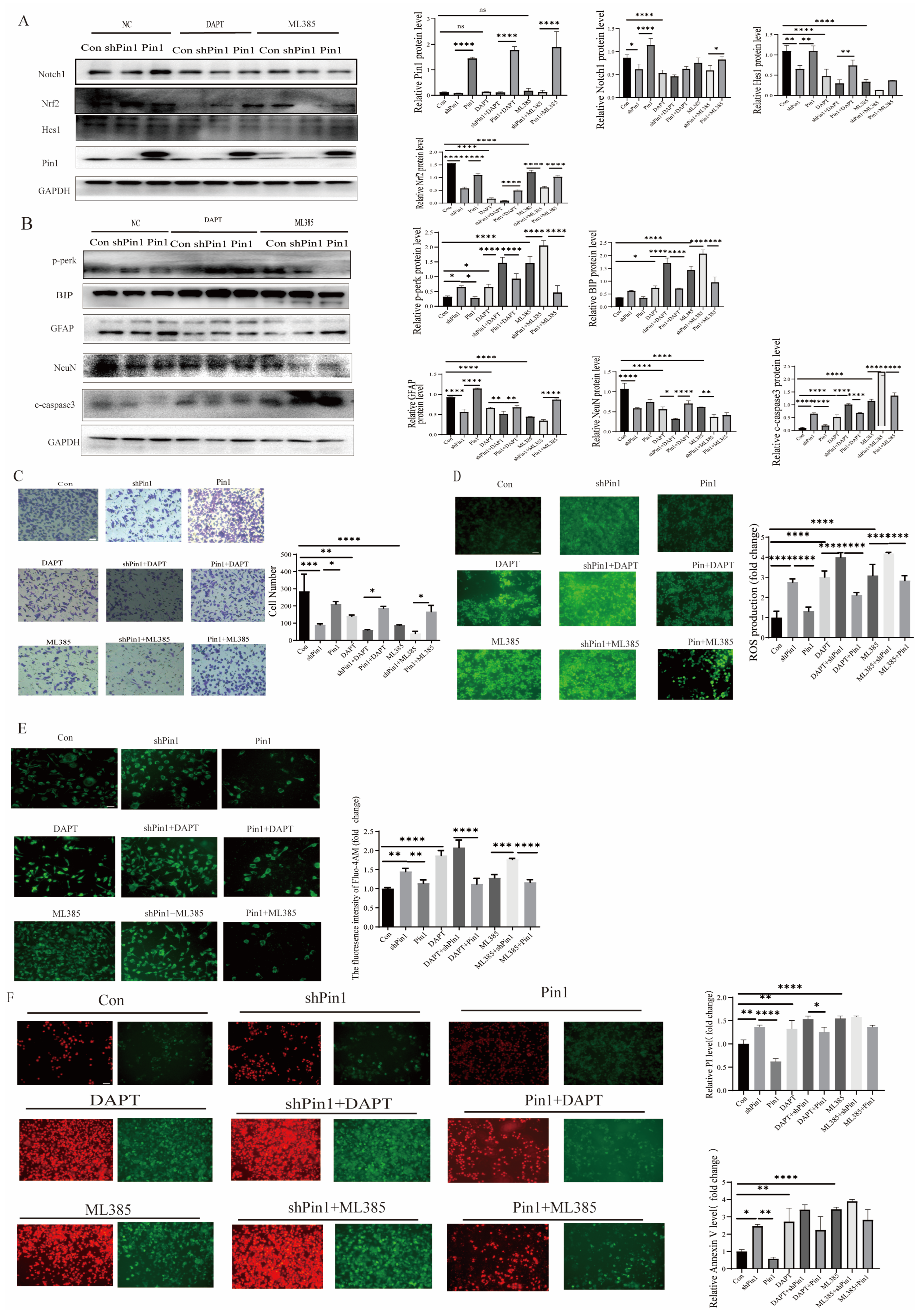
2.7. Notch1-Nrf2 Inhibitors Exacerbated Atra-Induced Damage in SHSY5Y Cells
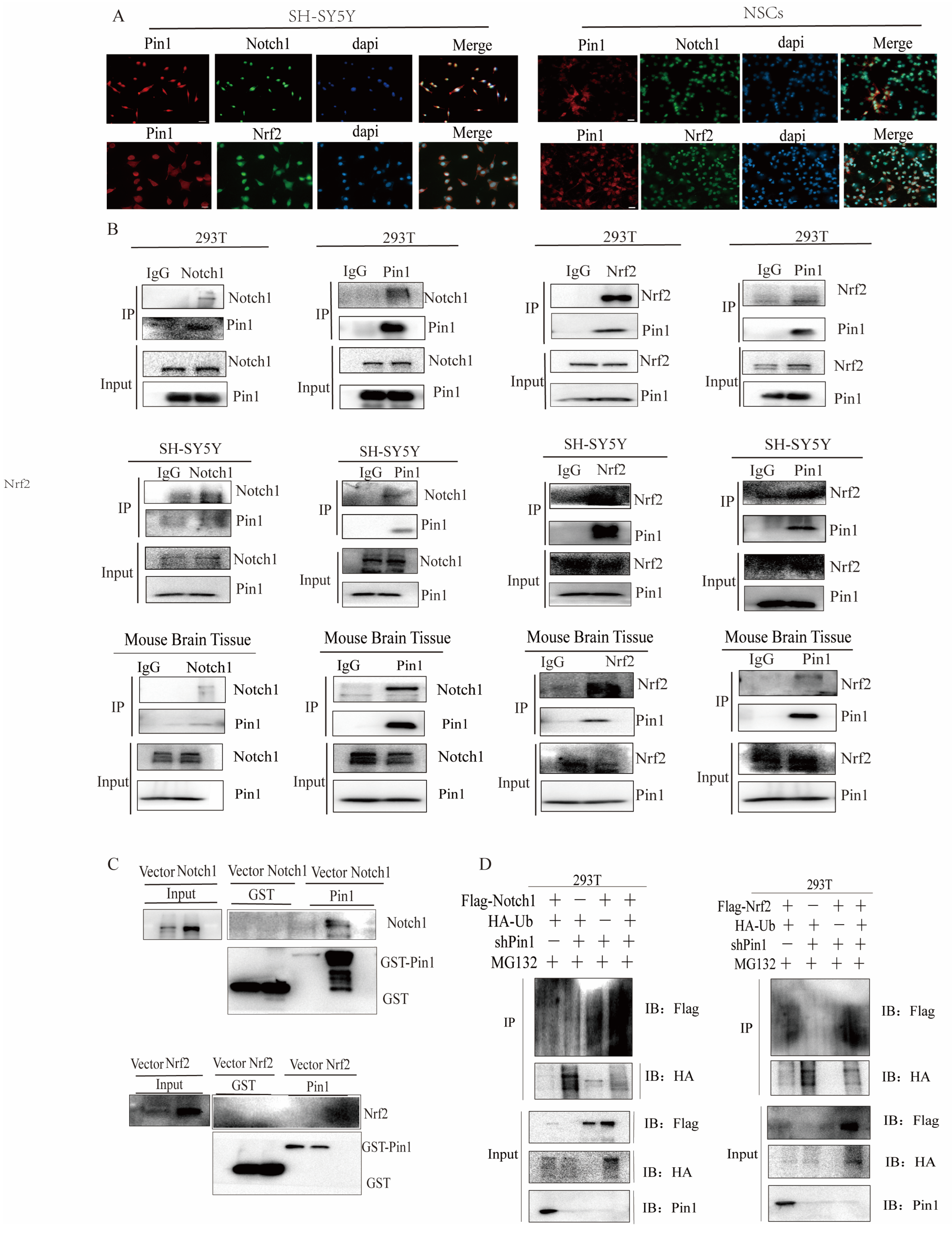
2.8. Pin1 Directly Interacts with Notch1-Nrf2
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Patients and Specimens
4.4. Cell Lines and Cell Culture
4.5. Cell Model of NTDs
4.6. Cell Transfection
4.7. Drug Treatment
4.8. Cloning Assay
4.9. Measurement of Oxidative Stress
4.10. Measurement of Malondialdehyde (MDA) and Glutathione (GSH) Levels
4.11. Assay of Intracellular Ca2+ with Fluo-4 AM
4.12. Western Blotting (WB)
4.13. Cell Proliferation Assays
4.14. Transwell Assays
4.15. PI/Annexin V Cell Apoptosis Assay
4.16. Ubiquitination Assay
4.17. Histological Analysis
4.18. Immunoprecipitation and GST Precipitation Assays
4.19. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, N.; Xu, J.; Zhang, X.; Li, S.; Zhu, W.; Cui, H.; Sun, Y.; Han, B.; Ma, A. Effect of Notch1 on neural tube defects and neural stem cell differentiation induced by all-trans retinoic acid. Mol. Med. Rep. 2021, 23, 220. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ji, G.; Zhou, J.; Du, J.; Li, X.; Shi, W.; Hu, Y.; Zhou, W.; Hao, A. Pcgf1 Regulates Early Neural Tube Development through Histone Methylation in Zebrafish. Front. Cell Dev. Biol. 2020, 8, 581636. [Google Scholar] [CrossRef] [PubMed]
- Botto, L.D.; Moore, C.A.; Khoury, M.J.; Erickson, J.D. Neural-tube defects. N. Engl. J. Med. 1999, 341, 1509–1519. [Google Scholar] [CrossRef]
- Yu, J.; Guo, Q.; Mu, J.B.; Zhang, T.; Li, R.K.; Xie, J. Nr2e1 Downregulation Is Involved in Excess Retinoic Acid-induced Developmental Abnormality in the Mouse Brain. Biomed. Environ. Sci. BES 2017, 30, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sen, J. Retinoic acid signaling regulates development of the dorsal forebrain midline and the choroid plexus in the chick. Development 2015, 142, 1293–1298. [Google Scholar] [CrossRef]
- Duester, G. Retinoic acid regulation of the somitogenesis clock. Birth Defects Res. Part C Embryo Today Rev. 2007, 81, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.X.; Au, K.S.; Morrison, A.C.; Fletcher, J.M.; Ostermaier, K.K.; Tyerman, G.H.; Northrup, H. Association of retinoic acid receptor genes with meningomyelocele. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. Retinoid signalling in the development of the central nervous system. Nat. Rev. Neurosci. 2002, 3, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Kozono, S.; Kats, L.; Nechama, M.; Li, W.; Guarnerio, J.; Luo, M.; You, M.H.; Yao, Y.; Kondo, A.; et al. Active Pin1 is a key target of all-trans retinoic acid in acute promyelocytic leukemia and breast cancer. Nat. Med. 2015, 21, 457–466. [Google Scholar] [CrossRef]
- Lu, K.P.; Hanes, S.D.; Hunter, T. A human peptidyl-prolyl isomerase essential for regulation of mitosis. Nature 1996, 380, 544–547. [Google Scholar] [CrossRef]
- Ranganathan, R.; Lu, K.P.; Hunter, T.; Noel, J.P. Structural and functional analysis of the mitotic rotamase Pin1 suggests substrate recognition is phosphorylation dependent. Cell 1997, 89, 875–886. [Google Scholar] [CrossRef]
- Lu, K.P.; Zhou, X.Z. The prolyl isomerase PIN1: A pivotal new twist in phosphorylation signalling and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 904–916. [Google Scholar] [CrossRef]
- Nakamura, K.; Kosugi, I.; Lee, D.Y.; Hafner, A.; Sinclair, D.A.; Ryo, A.; Lu, K.P. Prolyl isomerase Pin1 regulates neuronal differentiation via β-catenin. Mol. Cell. Biol. 2012, 32, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Kalpana, R.; Ansah, T.; Charlton, C.; Hale, A.; Channon, K.M.; Srinivasan, S.; Gangula, P.R. Impairment of Nrf2- and Nitrergic-Mediated Gastrointestinal Motility in an MPTP Mouse Model of Parkinson’s Disease. Dig. Dis. Sci. 2019, 64, 3502–3517. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Murakami, S.; Yoneda, T.; Nakamura, M.; Zhang, L.; Uezumi, A.; Fukuda, S.; Kokubo, H.; Tsujikawa, K.; Fukada, S. Evidence of Notch-Hesr-Nrf2 Axis in Muscle Stem Cells, but Absence of Nrf2 Has No Effect on Their Quiescent and Undifferentiated State. PLoS ONE 2015, 10, e0138517. [Google Scholar] [CrossRef]
- Rustighi, A.; Tiberi, L.; Soldano, A.; Napoli, M.; Nuciforo, P.; Rosato, A.; Kaplan, F.; Capobianco, A.; Pece, S.; Di Fiore, P.P.; et al. The prolyl-isomerase Pin1 is a Notch1 target that enhances Notch1 activation in cancer. Nat. Cell Biol. 2009, 11, 133–142. [Google Scholar] [CrossRef]
- Saeidi, S.; Kim, S.J.; Guillen-Quispe, Y.N.; Jagadeesh, A.S.V.; Han, H.J.; Kim, S.H.; Zhong, X.; Piao, J.Y.; Kim, S.J.; Jeong, J.; et al. Peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 directly binds and stabilizes Nrf2 in breast cancer. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2022, 36, e22068. [Google Scholar] [CrossRef]
- Yang, P.; Reece, E.A.; Wang, F.; Gabbay-Benziv, R. Decoding the oxidative stress hypothesis in diabetic embryopathy through proapoptotic kinase signaling. Am. J. Obstet. Gynecol. 2015, 212, 569–579. [Google Scholar] [CrossRef]
- Wang, M.; Kaufman, R.J. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature 2016, 529, 326–335. [Google Scholar] [CrossRef]
- Chen, R.; Huo, L.; Shi, X.; Bai, R.; Zhang, Z.; Zhao, Y.; Chang, Y.; Chen, C. Endoplasmic reticulum stress induced by zinc oxide nanoparticles is an earlier biomarker for nanotoxicological evaluation. ACS Nano 2014, 8, 2562–2574. [Google Scholar] [CrossRef]
- Logue, S.E.; Cleary, P.; Saveljeva, S.; Samali, A. New directions in ER stress-induced cell death. Apoptosis Int. J. Program. Cell Death 2013, 18, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pang, J.; Ye, L.; Zhang, Z.; Lin, S.; Lin, N.; Lee, T.H.; Liu, H. Disorders of the central nervous system: Insights from Notch and Nrf2 signaling. Biomed. Pharmacother. 2023, 166, 115383. [Google Scholar] [CrossRef]
- Cecconi, F.; Piacentini, M.; Fimia, G.M. The involvement of cell death and survival in neural tube defects: A distinct role for apoptosis and autophagy? Cell Death Differ. 2008, 15, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Cearns, M.D.; Escuin, S.; Alexandre, P.; Greene, N.D.; Copp, A.J. Microtubules, polarity and vertebrate neural tube morphogenesis. J. Anat. 2016, 229, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, W.; Li, X.; Gao, M.; Ji, S.; Tian, W.; Ji, G.; Du, J.; Hao, A. SOX19b regulates the premature neuronal differentiation of neural stem cells through EZH2-mediated histone methylation in neural tube development of zebrafish. Stem Cell Res. Ther. 2019, 10, 389. [Google Scholar] [CrossRef] [PubMed]
- Rogner, U.C.; Spyropoulos, D.D.; Le Novère, N.; Changeux, J.P.; Avner, P. Control of neurulation by the nucleosome assembly protein-1-like 2. Nat. Genet. 2000, 25, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.D.; Copp, A.J. Neural tube defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Zhao, H.; Niu, B.; Li, W.I.; Zhou, R.; Zhang, T.; Xie, J. Correlation of polymorphism of MTHFRs and RFC-1 genes with neural tube defects in China. Birth Defects Res. Part A Clin. Mol. Teratol. 2008, 82, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Poletta, F.A.; Rittler, M.; Saleme, C.; Campaña, H.; Gili, J.A.; Pawluk, M.S.; Gimenez, L.G.; Cosentino, V.R.; Castilla, E.E.; López-Camelo, J.S. Neural tube defects: Sex ratio changes after fortification with folic acid. PLoS ONE 2018, 13, e0193127. [Google Scholar] [CrossRef] [PubMed]
- Czeizel, A.E. Periconceptional folic acid-containing multivitamin supplementation for the prevention of neural tube defects and cardiovascular malformations. Ann. Nutr. Metab. 2011, 59, 38–40. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, G.; Luo, X.; Zhang, P.; Peng, S.; Cheng, X.; Wang, M.; Yang, X. Endoplasmic reticulum stress-related calcium imbalance plays an important role on Zinc oxide nanoparticles-induced failure of neural tube closure during embryogenesis. Environ. Int. 2021, 152, 106495. [Google Scholar] [CrossRef] [PubMed]
- Seervi, M.; Xue, D. Mitochondrial Cell Death Pathways in Caenorhabiditis elegans. Curr. Top. Dev. Biol. 2015, 114, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Driver, J.A.; Zhou, X.Z.; Lu, K.P. Pin1 dysregulation helps to explain the inverse association between cancer and Alzheimer’s disease. Biochim. Biophys. Acta 2015, 1850, 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Liou, Y.C.; Sun, A.; Ryo, A.; Zhou, X.Z.; Yu, Z.X.; Huang, H.K.; Uchida, T.; Bronson, R.; Bing, G.; Li, X.; et al. Role of the prolyl isomerase Pin1 in protecting against age-dependent neurodegeneration. Nature 2003, 424, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, R.; Pizzarelli, R.; Pedroni, A.; Fritschy, J.M.; Del Sal, G.; Cherubini, E.; Zacchi, P. Pin1-dependent signalling negatively affects GABAergic transmission by modulating neuroligin2/gephyrin interaction. Nat. Commun. 2014, 5, 5066. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, R.; De Filippo, R.; Middei, S.; Stancheva, S.; Pastore, B.; Ammassari-Teule, M.; Barberis, A.; Cherubini, E.; Zacchi, P. Pin1 Modulates the Synaptic Content of NMDA Receptors via Prolyl-Isomerization of PSD-95. J. Neurosci. 2016, 36, 5437–5447. [Google Scholar] [CrossRef]
- Fu, C.; Wu, Y.; Liu, S.; Luo, C.; Lu, Y.; Liu, M.; Wang, L.; Zhang, Y.; Liu, X. Rehmannioside A improves cognitive impairment and alleviates ferroptosis via activating PI3K/AKT/Nrf2 and SLC7A11/GPX4 signaling pathway after ischemia. J. Ethnopharmacol. 2022, 289, 115021. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Sui, M.; Lü, X.; Jin, D.; Zhuang, Z.; Yan, T. Electroacupuncture promotes neural stem cell proliferation and neurogenesis in the dentate gyrus of rats following stroke via upregulation of Notch1 expression. Mol. Med. Rep. 2015, 12, 6911–6917. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, N.; Chartoumpekis, D.V.; Kensler, T.W. Crosstalk between Nrf2 and Notch signaling. Free Radic. Biol. Med. 2015, 88, 158–167. [Google Scholar] [CrossRef]
- Nie, J.H.; Shen, Y.; Roshdy, M.; Cheng, X.; Wang, G.; Yang, X. Polystyrene nanoplastics exposure caused defective neural tube morphogenesis through caveolae-mediated endocytosis and faulty apoptosis. Nanotoxicology 2021, 15, 885–904. [Google Scholar] [CrossRef]
- Lian, X.; Lin, Y.M.; Kozono, S.; Herbert, M.K.; Li, X.; Yuan, X.; Guo, J.; Guo, Y.; Tang, M.; Lin, J.; et al. Pin1 inhibition exerts potent activity against acute myeloid leukemia through blocking multiple cancer-driving pathways. J. Hematol. Oncol. 2018, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhu, Z.; Xu, S.; Yang, L.N.; Liao, X.H.; Zheng, M.; Yang, D.; Wang, J.; Chen, D.; Wang, L.; et al. MicroRNA-140-5p inhibits hepatocellular carcinoma by directly targeting the unique isomerase Pin1 to block multiple cancer-driving pathways. Sci. Rep. 2017, 7, 45915. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Pang, J.; Ye, L.; Zhang, Z.; Kang, J.; Qiu, Z.; Lin, N.; Liu, H. Pin1 Downregulation Is Involved in Excess Retinoic Acid-Induced Failure of Neural Tube Closure. Int. J. Mol. Sci. 2024, 25, 5588. https://doi.org/10.3390/ijms25115588
Chen Y, Pang J, Ye L, Zhang Z, Kang J, Qiu Z, Lin N, Liu H. Pin1 Downregulation Is Involved in Excess Retinoic Acid-Induced Failure of Neural Tube Closure. International Journal of Molecular Sciences. 2024; 25(11):5588. https://doi.org/10.3390/ijms25115588
Chicago/Turabian StyleChen, Yuwen, Jiao Pang, Lu Ye, Zhentao Zhang, Junfeng Kang, Zhuotao Qiu, Na Lin, and Hekun Liu. 2024. "Pin1 Downregulation Is Involved in Excess Retinoic Acid-Induced Failure of Neural Tube Closure" International Journal of Molecular Sciences 25, no. 11: 5588. https://doi.org/10.3390/ijms25115588
APA StyleChen, Y., Pang, J., Ye, L., Zhang, Z., Kang, J., Qiu, Z., Lin, N., & Liu, H. (2024). Pin1 Downregulation Is Involved in Excess Retinoic Acid-Induced Failure of Neural Tube Closure. International Journal of Molecular Sciences, 25(11), 5588. https://doi.org/10.3390/ijms25115588