Abstract
Mycoviruses are usually transmitted horizontally via hyphal anastomosis and vertically through sporulation in natural settings. Oyster mushroom spherical virus (OMSV) is a mycovirus that infects Pleurotus ostreatus, with horizontal transmission via hyphal anastomosis. However, whether OMSV can be vertically transmitted is unclear. This study aimed to investigate the transmission characteristics of OMSV to progeny via basidiospores and horizontally to a new host. A total of 37 single-basidiospore offspring were obtained from OMSV-infected P. ostreatus and Pleurotus pulmonarius for Western blot detection of OMSV. The OMSV-carrying rate among monokaryotic isolates was 19% in P. ostreatus and 44% in P. pulmonarius. Then, OMSV-free and OMSV-infected monokaryotic isolates were selected for hybridization with harvested dikaryotic progeny strains. Western blot analyses of the offspring revealed that the OMSV transmission efficiency was 50% in P. ostreatus and 75% in P. pulmonarius, indicating vertical transmission via sexual basidiospores. Furthermore, we observed the horizontal transfer of OMSV from P. pulmonarius to Pleurotus floridanus. OMSV infection in P. floridanus resulted in significant inhibition of mycelial growth and yield loss. This study was novel in reporting the vertical transmission of OMSV through basidiospores, and its infection and pathogenicity in a new host P. floridanus.
1. Introduction
Mycoviruses are prevalent in fungi, encompassing yeasts, macrofungi, and various pathogenic fungi that infect plants, insects, and humans. The first identification of a mycovirus occurred in 1962 with the discovery of the La France isometric virus (LIV) in the economically significant mushroom species Agaricus bisporus [1,2]. Since then, approximately 60 mycoviruses infecting edible fungi have been recorded, comprising double-stranded (ds)RNA, positive-sense single-stranded (+)ssRNA, and negative-sense single-stranded (−)ssRNA viruses [3,4]. Although most mycoviruses establish latent or symptomless infections within their fungal hosts, some can cause severe symptoms such as changes in growth, colony morphology, spore production, pigmentation, and pathogenicity [1,5,6,7].
In nature, mycoviruses are primarily transmitted horizontally via hyphal fusion and vertically through sporulation [8]. Vertical transmission occurs mainly via the passage of asexual and sexual spores from parents to offspring, with transmission efficiency varying among different fungal viruses [9,10]. For example, the (+)ssRNA Cryphonectria hypovirus 1 (CHV1) exhibited a 100% transmission rate to conidiospore progeny, albeit only 50% of the offspring showed virulence in the offspring of sexual spores [11]. Similarly, the dsRNA Fusarium graminearum hypovirus 1 (FgHV1) infecting F. graminearum can be transmitted vertically through asexual spores with a 100% transmission rate [12]. Another dsRNA Rhizoctonia solani partitivirus (RsPV-BS5) can exist stably in the sexual spores of R. solani AG-3, indicating that RsPV-BS5 can carry out vertical transmission in R. solani [13]. However, few studies exist about the vertical transmission of mycoviruses within edible mushroom species. In A. bisporus, the LIV has an average carrier rate of 65–75% within basidiospores [14]. In Lentinula edodes, L. edodes negative-stranded RNA virus 1 (LeNSRV1) is transmitted through basidiospores, albeit with relatively low efficiency [15]. L. edodes spherical virus (LeSV) is a dsRNA mycovirus infecting L. edodes, exhibiting a 90% LeSV-carrying rate in basidiospores [16]. To date, no studies have investigated the vertical transmission involving (+)ssRNA viruses in edible mushroom species.
Horizontal transmission via mycelium serves as a major means of mycovirus spread, with mycoviruses usually exhibiting a restricted host range, infecting the same or closely affiliated compatible groups [1,7,17]. For example, RsPV-BS5 can be transmitted horizontally from the virus-infected strain BS-5 to the virus-free strain 06-2-15 of R. solani [13]. Through co-infection experiments, Beauveria bassiana chrysovirus 2 (BbCV2) was horizontally transmitted from the virus-infected strain BbOFZK152 to the virus-free strain BbOFDH1-5-GFP [18]. Additionally, Cryphonectria naterciae fusagravirus 1 (CnFGV1) exhibited the capacity for transmissibility to other Cryphonectria species [19]. Of the ten Cryphonectria species isolates tested, six were successfully infected by CHV1 from C. parasitica via hyphal anastomosis [20]. In A. bisporus, mushroom virus X (MVX) was demonstrated to be transmitted horizontally via hyphal anastomosis from an MVX-infected strain to five other MVX-free strains [21]. Additionally, the horizontal transmission of Cordyceps chanhua partitivirus 1 (CchPV1) was observed in C. chanhua, spreading from strain RCEF5997 to RCEF5833 [22].
OMSV was first reported infecting P. ostreatus mushrooms in Korea in 2003, which was closely related to the mushroom die-back disease. It is a spherical virion with a diameter of 27 nm, harboring a (+)ssRNA genome. The complete genome of OMSV contains 5784 nucleotides encoding 7 open reading frames, with ORF1 encoding the RNA-dependent RNA polymerase and ORF2 encoding the coat protein (CP). However, the functions of the proteins predicted to be encoded by ORF3-7 currently remain unidentified [23]. OMSV occurrence has been reported in several regions in China, including Jilin province, Shandong province, and Beijing city [24,25,26]. Previous studies found that OMSV could be transmitted horizontally from an infected strain to a virus-cured strain by hyphae in P. ostreatus strain 8129, resulting in slowed mycelial growth, malformations of the fruiting bodies, and yield loss [26]. Interestingly, the OMSV could also cross the species barrier and horizontally transmit from P. ostreatus to P. pulmonarius [27]. However, no studies have investigated the vertical transmission of OMSV.
This study aimed to investigate whether OMSV could be transmitted vertically through sexual basidiospores and whether it posed a threat to other Pleurotus species through horizontal transmission. This study demonstrated that OMSV could be transmitted to offspring via basidiospores, making this study novel in reporting the vertical transmission of OMSV. In addition, the horizontal transmission of OMSV from P. pulmonarius to P. floridanus was observed. The OMSV infection not only reduced the mycelial growth rate but also led to a decrease in the yield of fruiting bodies in P. floridanus. This study provided a solid foundation for developing virus-free and antiviral strains, thereby contributing to the sustainable growth of the P. ostreatus industry.
2. Results
2.1. Transmission of OMSV by Single Spores
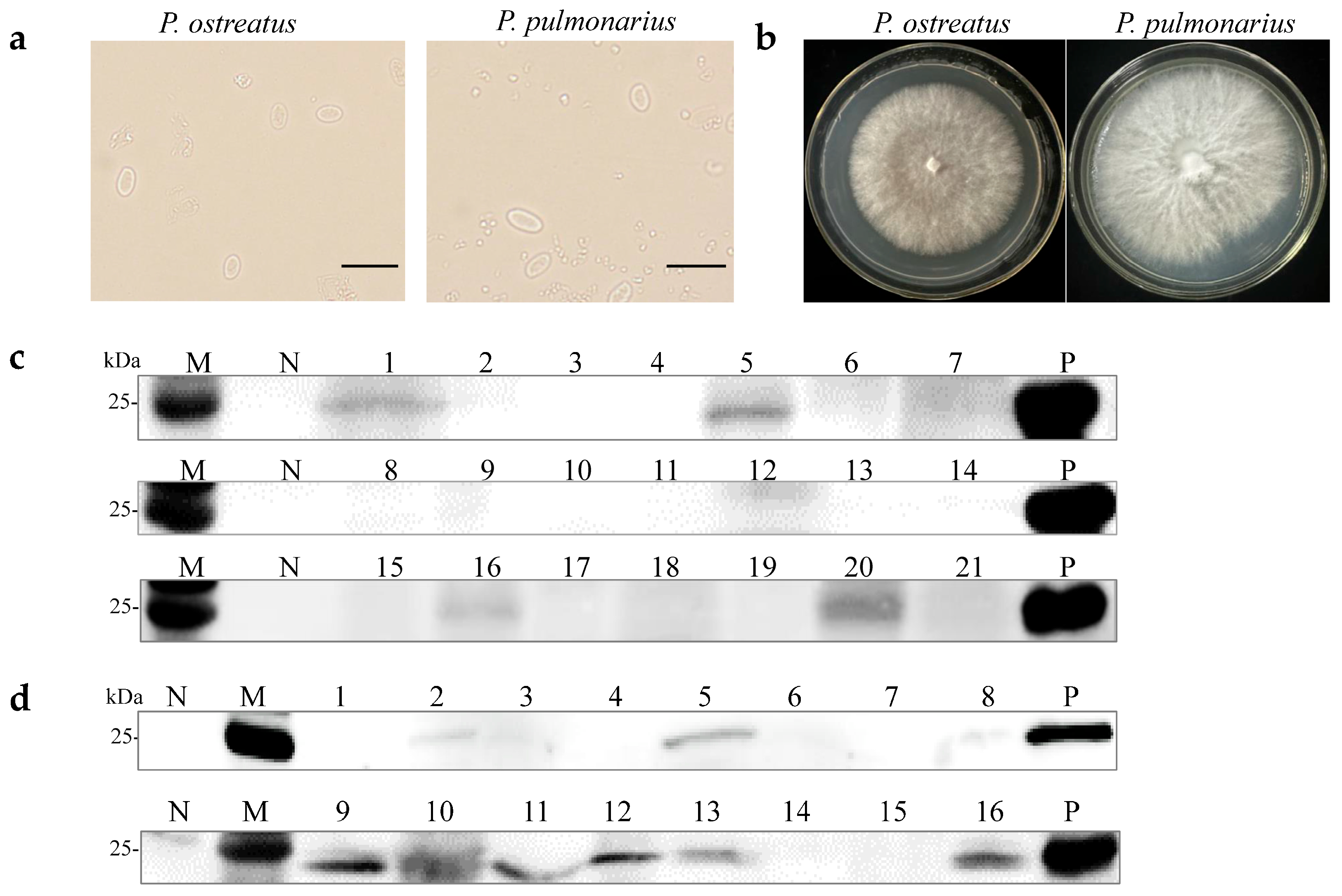
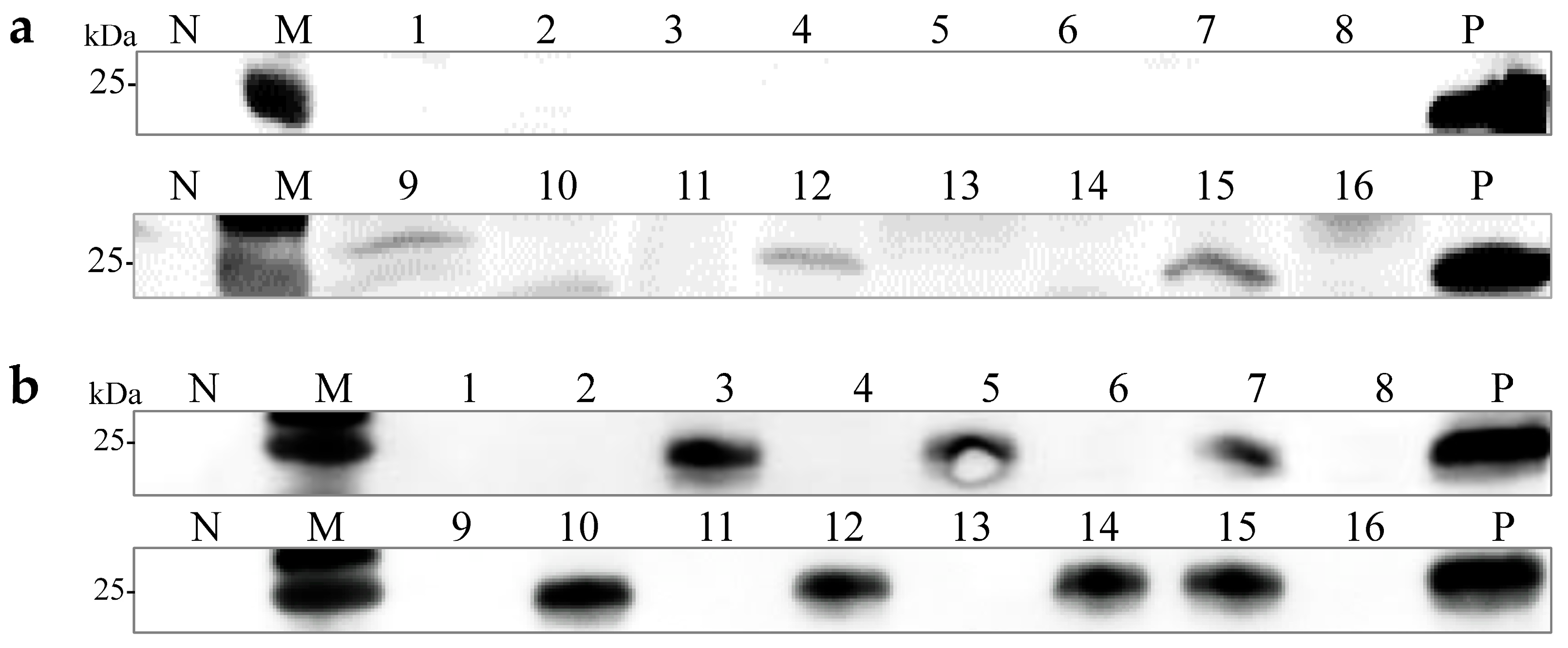
A previous study demonstrated that OMSV could be horizontally transmitted via mycelia [27]. Two OMSV-infected strains, P. ostreatus strain 8129 and P. pulmonarius strain XH2208, were cultivated to investigate the possibility of vertical transmission through sexual basidiospores. Basidiospores from both P. ostreatus and P. pulmonarius were isolated from the basidiocarp and cultured on potato dextrose agar (PDA) plates (Figure 1a). Further, 21 basidiospores of P. ostreatus and 16 basidiospores of P. pulmonarius were collected and germinated, resulting in the development of monokaryotic colonies within 10 days (Figure 1b). Optical microscopy was employed to confirm the monokaryotic strains based on the absence of clamp connections, followed by the further detection of OMSV. Western blot analyses revealed that 19% of P. ostreatus monokaryotic strains (4/21) carried OMSV (Figure 1c, Lanes 1, 5, 16, and 20), whereas 44% of P. pulmonarius monokaryotic strains (7/16) carried OMSV (Figure 1d, Lanes 2, 5, 9, 11, 12, 13, and 16). These findings indicated that OMSV could persistently exist within the sexual basidiospores of both P. ostreatus strain 8129 and P. pulmonarius strain XH2208.
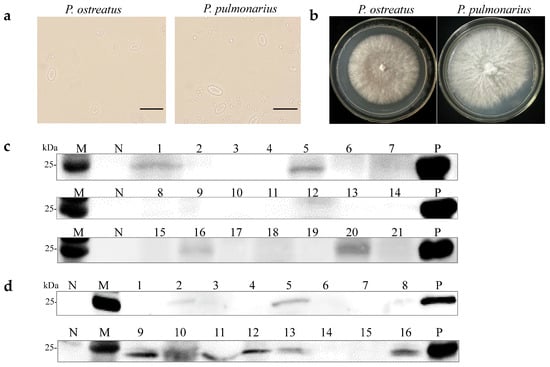
Figure 1.
Detection of OMSV for monokaryotic strains of P. ostreatus and P. pulmonarius. (a) Microscopic morphology of basidiospores of P. ostreatus or P. pulmonarius. Scale bars = 10 µm. (b) Subculturing of selected monokaryotic progeny strains from the colonies of P. ostreatus or P. pulmonarius. Western blot detection of OMSV in 21 P. ostreatus monokaryotic strains (c) and 16 P. pulmonarius monokaryotic strains (d). OMSV-free and OMSV-infected strains served as negative (N) and positive (P) controls, respectively. Lane M, molecular weights of protein marker.
2.2. Transmission of OMSV from Single Basidiospores to Dikaryotic Strain after Single-Spore Hybridization
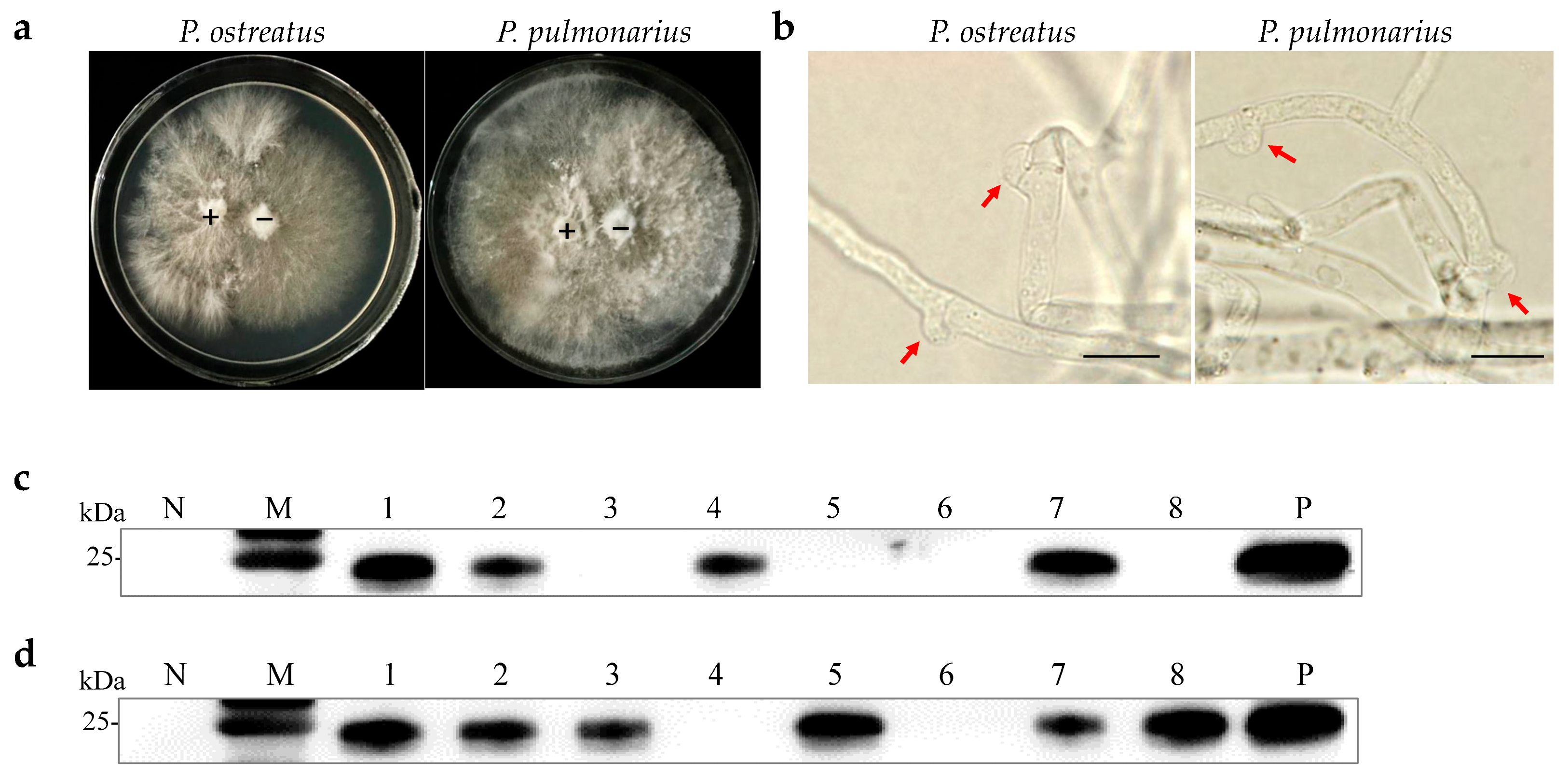
Monosporous hybridization is a vital approach in breeding Pleurotus species cultivars. Six OMSV-free and four OMSV-infected P. ostreatus monokaryotic isolates were selected and cultured equidistantly for co-cultivation to investigate whether OMSV could be transmitted to offspring through monosporous hybridization. Similarly, four OMSV-free and seven OMSV-infected P. pulmonarius monokaryotic isolates were selected and cultured equidistantly for co-cultivation. Each OMSV-infected monokaryotic isolate was crossed with each OMSV-free monokaryotic isolate in all possible combinations, resulting in 24 P. ostreatus crosses and 28 P. pulmonarius crosses. After 14 days of incubation, dikaryotic progeny strains were formed at both ends of the anastomosis where the hyphae of the two monokaryotic isolates met. The progeny strains underwent subculture on PDA (Figure 2a), followed by the microscopic examination of clamp connections to confirm the presence of dikaryotic hyphae (Figure 2b). The presence of clamp connection indicated the successful mating of these cross combinations. A total of eight successful P. ostreatus crosses and eight P. pulmonarius crosses were identified, and then the sixteen dikaryotic progeny strains were subjected to Western blot detection of OMSV. The results revealed that 50% of P. ostreatus dikaryotic progeny strains (4/8) carried OMSV (Figure 2c) whereas 75% of P. pulmonarius dikaryotic progeny strains (6/8) carried OMSV (Figure 2d).
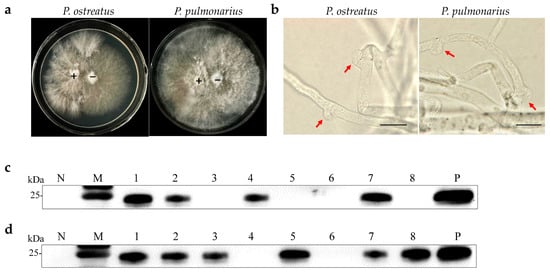
Figure 2.
OMSV detection in dikaryotic progeny strains of P. ostreatus and P. pulmonarius. (a) Dual culture of OMSV-infected (+) and OMSV-free (–) monokaryotic isolates. (b) Microscopic morphology of clamp connection. The red arrow indicates the clamp connection. Scale bars = 10 µm. Western blot analysis of OMSV in dikaryotic progeny strains of P. ostreatus (c) and P. pulmonarius (d); OMSV-free and OMSV-infected strains served as negative (N) and positive (P) controls, respectively. Lane M, molecular weights of protein marker.
2.3. Infection of the New Host P. floridanus by OMSV across the Species Barrier
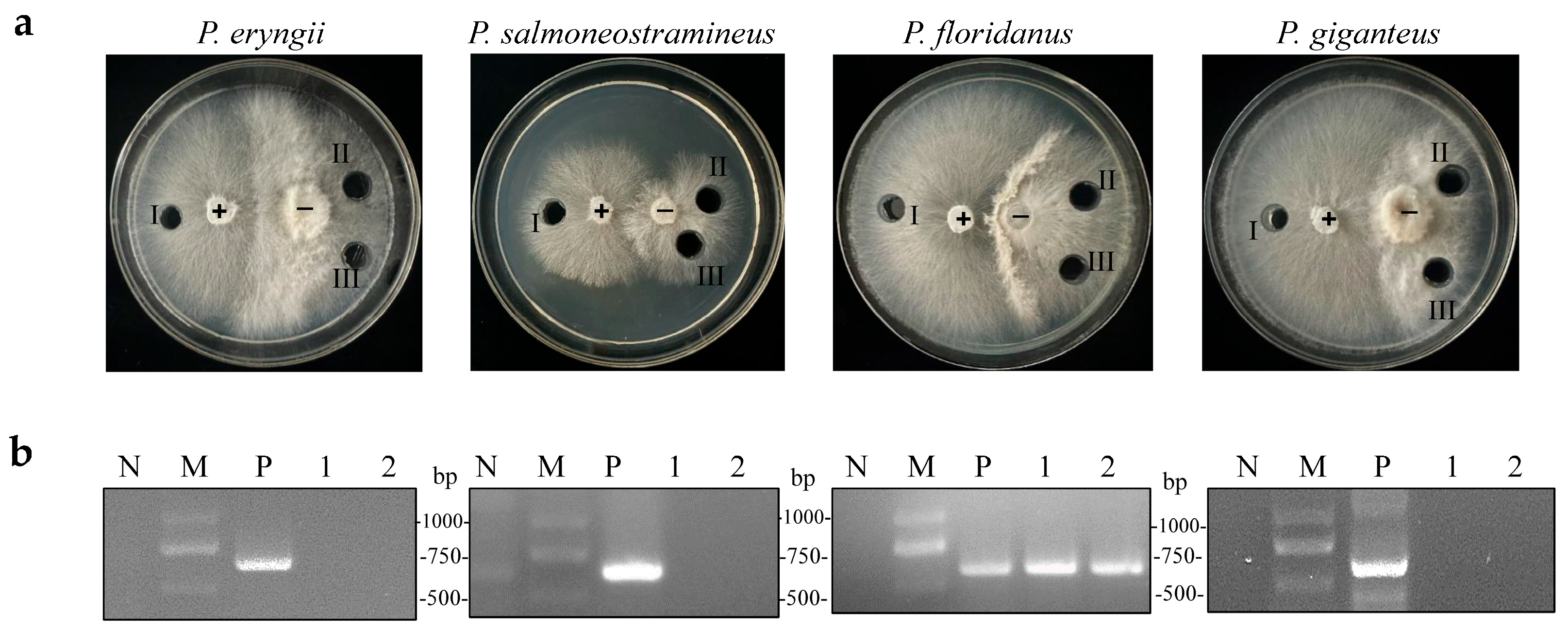
A previous study discovered that OMSV could cross the species barrier and infect P. pulmonarius [23]. An OMSV-infected P. pulmonarius strain was employed as a donor, which was co-cultivated with OMSV-free strains of P. eryngii, P. salmoneostramineus, P. floridanus, and P. giganteus, to investigate whether OMSV could infect other Pleurotus species. After several days of co-cultivation, an antagonistic relationship was observed between the donor and recipient strains (Figure 3a). Subsequently, an inoculum derived from the donor culture and two inoculums from the recipient culture were sub-cultured for 7 days. Reverse-transcription (RT) polymerase chain reaction was then employed to confirm the presence or absence of OMSV within all the tested strains. P. floridanus tested positive for OMSV, whereas the other three Pleurotus species remained negative under identical conditions (Figure 3b). Therefore, this result suggested a successful transfer of OMSV from P. pulmonarius to P. floridanus.
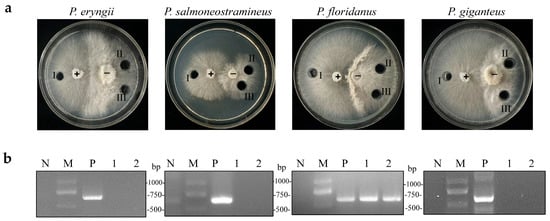
Figure 3.
Co-cultivation of OMSV-positive (+) P. pulmonarius and OMSV-free (−) Pleurotus spp. (a) Co-cultivation of the donor strain with P. eryngii, P. salmoneostramineus, P. floridanus, and P. giganteus. One inoculum from the donor side (I) and two inocula from the recipient (II and III) were sub-cultivated for further detection of OMSV. (b) RT-PCR detection of OMSV in different inoculants. OMSV-free strains of P. eryngii C1021, P. salmoneostramineus TH20901, P. floridanus LDPF2305 or P. giganteus ZD2308 served as negative controls (N). M, DNA Marker2000.
2.4. Slowing down of the Mycelial Growth of P. floridanus by OMSV Infection
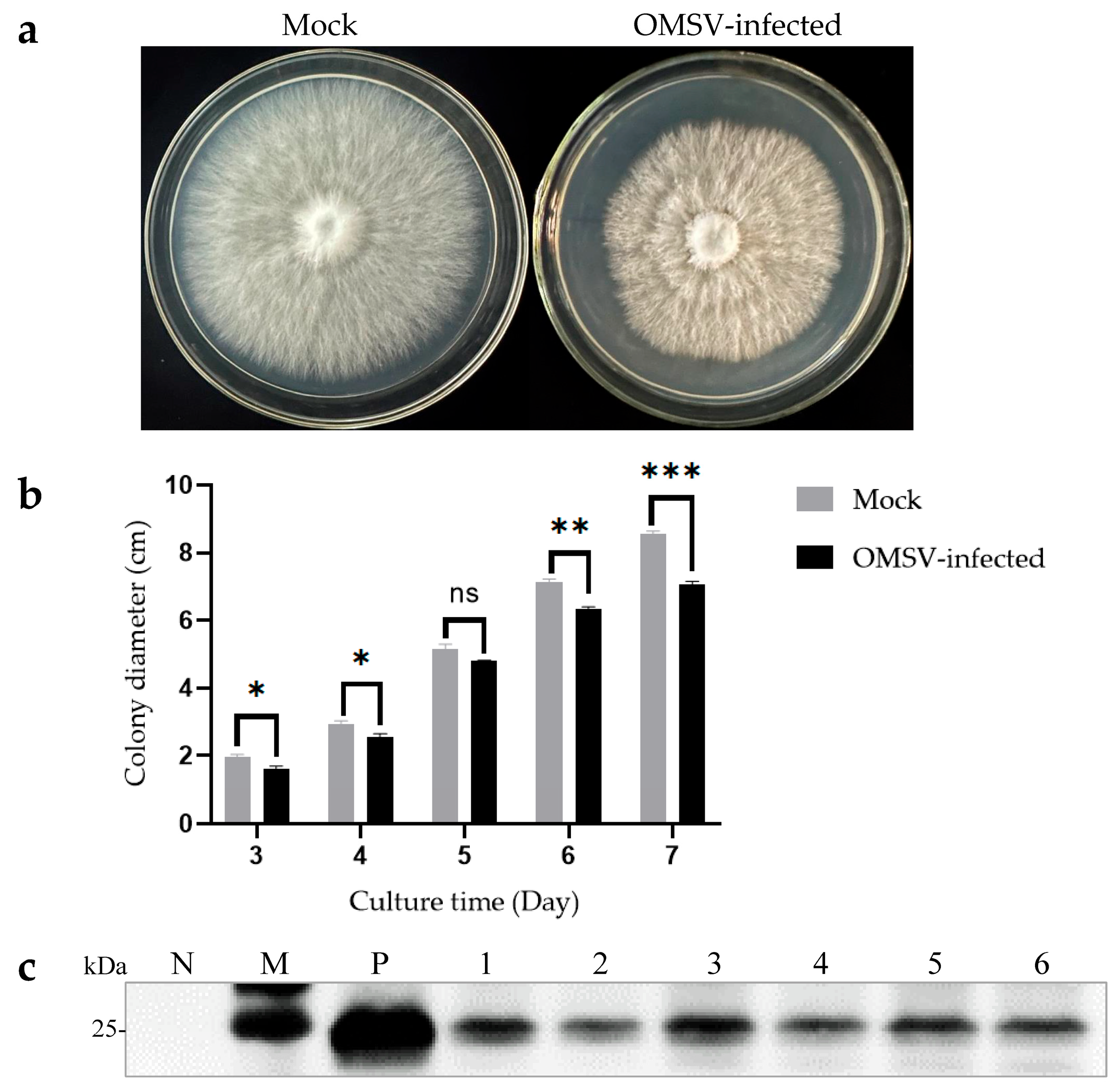
The isogenic strains that were OMSV-free and OMSV-infected were cultured on PDA plates to investigate the effect of OMSV infection on the growth of P. floridanus mycelium (Figure 4a). After 7 days of incubation, the OMSV-infected strain showed a significantly slower mycelial growth compared with the OMSV-free strain (Figure 4b). To confirm this observation, the mycelial growth rate was monitored by measuring colony diameters every day for 1 week. On day 7 after inoculation, the OMSV-infected strain exhibited a growth rate approximately 0.83-fold that of the OMSV-free strain.
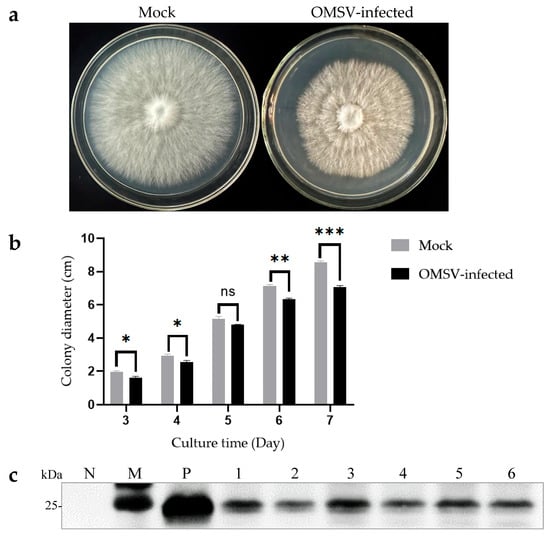
Figure 4.
Morphological characteristics of OMSV-free and OMSV-infected strains of Pleurotus floridanus following 7 days of incubation. Colony morphology (a) and growth rate (b) of the hyphae of OMSV-infected and OMSV-free (mock) strains of P. floridanus. (c) Western blot analysis of OMSV in P. floridanus fruiting bodies. Samples 1–6 correspond to six biological replicates. The OMSV-free P. floridanus sample served as negative control (N) and OMSV-infected P. pulmonarius served as positive control (P); Lane M, molecular weights of protein marker. The independent sample t-test was performed between the OMSV-free and OMSV-infected isogenic strains (ns, not significant; * p < 0.05; ** p < 0.01; *** p < 0.001).
2.5. A Reduction in Fruiting Body Yield of P. floridanus by OMSV Infection
Cultivation experiments were conducted using both OMSV-free and OMSV-infected isogenic strains to explore the potential impact of OMSV infection on the fruiting bodies of P. floridanus. The average time it took for the first flush of mushrooms to be harvested from inoculation was calculated. Interestingly, the OMSV-infected strain required an additional 4 days for harvest relative to the OMSV-free strain, indicating a deceleration in mycelial growth during culture (Table 1). To evaluate yield, both the first and second flushes of fruiting bodies were harvested and their average yield was calculated (Table 1). Significant differences were observed between the yields of the OMSV-free and OMSV-infected strains in the first flush. The average yield per bag for cultivated fruiting bodies in the OMSV-free strain was 194.08 g/bag, representing a 1.36-fold increase compared with that of its infected counterpart (141.94 g/bag). Upon reaching the second flush, the average yield per bag for the OMSV-infected strain decreased to 126.98 g/bag, representing a reduction by 0.72-fold compared with that of its uninfected counterpart (176.97 g/bag) (Table 1). The findings strongly indicated that OMSV infection led to a decrease in fruiting body yield in P. floridanus. Deformed fruiting bodies were collected and Western blot analysis was performed to confirm whether OMSV existed in these fruiting bodies. The results revealed the presence of OMSV-CP in the fruiting bodies of P. floridanus (Figure 4c).

Table 1.
Impact of OMSV infection on Pleurotus floridanus measured in cultivation tests.
2.6. Vertical Spread of OMSV in the New Host
Single spores of P. floridanus were collected and the monokaryotic strains formed after single-spore culture were examined to assess the vertical transmission potential of OMSV via basidiospores in a new host. A total of 16 spores were randomly selected for Western blot analysis, revealing that 19% of P. floridanus monokaryotic strains (3/16) carried OMSV (Figure 5a). Thirteen OMSV-free and three OMSV-infected P. floridanus monokaryotic isolates were selected and cultured at equidistance for co-cultivation. Each OMSV-infected monokaryotic isolate was crossed with each OMSV-free monokaryotic isolate in all possible combinations, resulting in 39 distinct P. floridanus crosses. After 14 days of incubation, dikaryotic progeny strains were observed at both ends of the anastomosis where the hyphae of the two monokaryotic isolates met, and these dikaryotic progeny strains were sub-cultured. Then, 16 successful hybridization groups were identified, and the dikaryotic strains were examined using Western blot analysis, revealing that 44% of P. floridanus dikaryotic progeny strains (7/16) carried OMSV (Figure 5b).
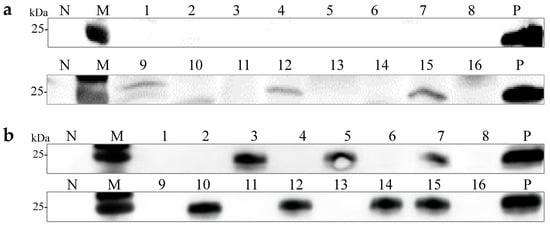
Figure 5.
Vertical transmission of OMSV in P. floridanus. (a) Western blot analysis confirmed OMSV infection in P. floridanus monokaryotic strains. (b) Western blot analysis of OMSV detection in P. floridanus dikaryotic progeny strains. The healthy P. floridanus sample served as negative control (N). Lane M, molecular weights of protein marker.
3. Discussion
Mycoviruses typically spread vertically through sporulation [8] but the efficiency varies among different fungal viruses [10]. For instance, CHV1 can be transmitted to conidiospore offspring with a 100% transmission rate but maintains only 50% virulence in sexual spores offspring [11]. Similarly, RsPV-BS5 was found in R. solani basidiospores, confirming its vertical transmission [13]. In L. edodes, the prevalence rate of LeSV in basidiospores was 90% [16]. However, investigations into the vertical transmission of mycoviruses in P. ostreatus are limited. This study demonstrated, for the first time, the vertical transmission of OMSV via basidiospores in P. ostreatus, P. pulmonarius, and P. floridanus. Monokaryotic strains of these Pleurotus species were found to carry OMSV, with carrying rates of 19%, 44%, and 19%, respectively. This suggested the vertical transmission of OMSV through sexual basidiospores. Based on these results, the transmission of OMSV to offspring through single-spore hybridization was explored. In the present study, dikaryotic progeny strains of P. ostreatus, P. pulmonarius, and P. floridanus were found to carry OMSV, with carrying rates of 50%, 75%, and 44%, respectively. Overall, this indicated the potential of OMSV to transfer vertically through successive generations within its host organism. This study was novel in employing single-spore hybridization technology to explore the vertical transmission of OMSV. Studies have reported that the vertical transmission efficiency of fungal viruses is related to the age of mycelia [28,29,30,31]. Based on it, we will explore the effect of mycelium age on the vertical transmission of OMSV. Considering the importance of single-spore hybridization in Pleurotus breeding, an OMSV-free parental strain is the first choice in cultivating virus-free cultivars. As for the parent strains that carry OMSV, we can screen the OMSV-free single spores for hybridization to obtain OMSV-free strains.
The horizontal transmission of mycoviruses occurs through mycelia. The Leptosphaeria biglobosa botybirnavirus 1 can cross-species transmit from Leptosphaeria biglobosa to Botrytis cinerea [32]. The Cryphonectria nitschkei chrysovirus 1 can be horizontally transmitted from its original host C. nitschkei to C. radicalis and C. naterciae strains [33]. A previous study conducted by the authors confirmed the horizontal transmission capability of the OMSV Chinese strain (OMSV-Ch) from P. ostreatus 8129 to P. pulmonarius strain XH2208 via mycelial contact [27]. In this study, OMSV-infected P. pulmonarius was co-cultured with other strains of Pleurotus species, revealing that OMSV could indeed spread horizontally from P. pulmonarius to P. floridanus. However, attempts to transmit OMSV to P. eryngii, P. salmoneostramineus, and P. giganteus were unsuccessful, likely due to a robust transmission barrier between the test strains. In addition, phylogenetic analyses suggested a closer evolutionary relationship between P. pulmonarius and P. floridanus but indicated a distant evolutionary relationship with P. eryngii, P. giganteus, and P. salmoneostramineus [34]. This implied the existence of a more formidable interspecies transmission barrier between these two non-transmissible species. In order to understand the interaction between OMSV and fungi, it is necessary for us to explore the molecular mechanism of OMSV movement in Pleurotus species. The OMSV-infected P. floridanus can be used as a donor strain to study whether the OMSV can infect the other OMSV-free Pleurotus species, which will help us understand the mechanisms underlying OMSV’s horizontal transmission.
Mycovirus infections often manifest with subtle symptoms or asymptomatically, but some can significantly affect fungal hosts [6]. The LIV has been implicated in causing mycelial degeneration and substantial yield reductions in A. biporus [35]. The MVX can destroy the primordium, leading to premature opening, discoloration, and malformed fruiting bodies of A. biporus [21,36]. In L. edodes, LePV1 has been associated with manifestations of both mycelial degradation and abnormal fruiting body formation [37]. The Pleurotus ostreatus ASI2792 mycovirus (PoV-ASI2792) has been reported to slow down mycelial growth and cause yield loss in the fruiting bodies of P. ostreatus [38]. More recently, it was shown that OMSV-Ch inhibited mycelial growth, induced dysmorphic symptoms in fruiting bodies, and reduced mushroom yield in both P. ostreatus and P. pulmonarius [27]. In the present study, the OMSV infection in P. floridanus resulted in a slowdown of mycelial growth and a decrease in mushroom yields. Thus, across three different Pleurotus species, OMSV emerged as a likely primary causative agent of yield losses in the host. Although the exact mechanisms underlying OMSV pathogenicity remain unclear, Previously, it was reported that the infection of PoV in P. ostreatus affects mycelial growth and fruiting body formation by directly reducing gene expression and then impairing the activity of some extracellular enzymes [38]. The OMSV infection reduced the carboxymethyl cellulase and laccase activity in the P. pulmonarius strain [27]. Further studies concerning the relative expression levels of genes and the activity levels of other extracellular enzymes such as amylase, aspartic protease, cellulase, chitinase, β-glucosidase, lipase, manganese peroxidases, polygalacturonase, and xylanase are necessary to investigate. This exploration aimed to better understand the potential molecular underpinnings of the detrimental effects of OMSV infection.
4. Materials and Methods
4.1. Single-Spore Isolation of P. ostreatus and P. pulmonarius
To collect basidiospores from the OMSV-infected mature fruiting bodies of P. floridanus strain LDPF2305, P. ostreatus strain 8129, and P. pulmonarius strain XH2208, the stipes were removed. All the tested strains were derived from the Fungarium of Ludong University. The pileus was gently positioned, with the gills facing down, on a sterilized Petri dish to allow basidiospores to naturally shed from the gills and settle onto the dish surface, thereby forming a spore print. Subsequently, the basidiospores were carefully harvested and mixed with sterile water to prepare a spore suspension. The basidiospore count was precisely determined using a hemocytometer (Shanghai Qiujing Biochemical Rechemical Reagent and Instrument Co., Ltd., Shanghai, China), and spore suspension was accordingly adjusted to a concentration of 100 basidiospores/mL. A volume of 100 μL of the diluted basidiospore suspension was evenly spread onto PDA to isolate 20–30 monokaryotic strains. The inoculated plates were then incubated at a constant temperature of 24 °C for 10 days until colonies of approximately 1 cm in diameter emerged from germinating basidiospores. The microscopic examination confirmed the absence of clamp connections.
4.2. Monosporous Hybridization
Monosporous hybridization was performed using the two-point method. A dual-culture technique was employed to facilitate the anastomosis of monokaryotic isolates from two strains each of P. floridanus, P. ostreatus, and P. pulmonarius, encompassing all feasible pairings. An OMSV-free monokaryotic isolate and an OMSV-infected monokaryotic isolate were inoculated on opposite sides of a PDA, maintaining a 10 mm distance between them. The plate was then incubated at a constant temperature of 25 °C. Dikaryotic progeny strains were formed at both ends of the anastomosis where the hyphae of the two monokaryotic isolates met, typically requiring approximately 14 days. The dikaryotic progeny strains were sub-cultured on PDA. Successful hybridization was confirmed if clamp connection structures were observed under the microscope.
4.3. Protein Extraction and Western Blot
The hyphae to be detected was ground into powder with liquid nitrogen, followed by thorough mixing with 2 × SDS loading buffer [27]. This mixture was subjected to boiling at a temperature of 100 °C for 10 min, and then it was centrifuged for 10 min at 12,000 rpm. The samples were separated by electrophoresis via 12.5% SDS-PAGE gels and subsequently transferred onto a nitrocellulose membrane (GE Healthcare). For detection of OMSV, the membrane was subjected to incubation with polyclonal antibodies against OMSV CP, followed by treatment with horseradish peroxidase-conjugated goat anti-rabbit secondary antibodies [27]. The detection of antibody–antigen binding interactions was visualized by using Omni-ECL™ Pico Light Chemiluminescence Kit. The secondary antibody and chemiluminescence kit were sourced from Shanghai Epizyme Biomedical Technology (Shanghai, China).
4.4. Horizontal Transmission of OMSV
Horizontal transmission experiments were executed using OMSV-infected P. pulmonarius as a donor strain. OMSV-infected strain (P. pulmonarius strain XH2208) and OMSV-free strain (P. floridanus strain LDPF2305, P. eryngii strain C1021, P. salmoneostramineus strain TH20901, and P. giganteus strain ZD2308) were co-cultivated separately on PDA at a constant temperature of 25 °C for a duration of 7 days, and all strains were from the Fungarium of the Ludong University. Upon completion of the co-culture, three mycelial agar plugs (I, II, and III) were selected for sub-culture on PDA. The occurrence of horizontal transmission of OMSV was subsequently detected through RT-PCR amplification, utilizing OMSV-specific primer.
4.5. RNA Extraction and Reverse Transcription PCR (RT-PCR)
Approximately 0.1 g fresh mycelium was collected and homogenized in liquid nitrogen. Total RNA extraction was accomplished using RNA Easy Fast Plant Tissue Kit (Tiangen, Beijing, China). To perform the reverse transcription reaction, 10 μL RT master mixture was prepared, comprising 5 μL ddH2O, 3 μL 5 × RT-PCR buffer, 1 μL dNTP mix, 0.5 μL reverse primer (OMSV-R, GAGATGTAGACRTTGAAAGC), 0.25 μL M-MLV reverse transcriptase, and 0.25 μL RNase Inhibitor, which was then combined with 5 μL of RNA. Reverse transcription proceeded at 37 °C for a duration of 90 min. Following the RT reaction, 30 μL PCR mixture containing 15 µL 2 × Taq PCR MasterMixII (Tiangen, Beijing, China), 2 µL cDNA template, 1 µL of specific primers (OMSV-F/OMSV-R, ACCCCCCCAGGATCTCAAGCTTC/GAGATGTAGACRTTGAAAGC), and 11 µL ddH2O [26]. The resulting PCR amplicons were subjected to electrophoretic separation in a 1% agarose gel, operating at a voltage of 120 V for 45 min.
4.6. Mycelial Growth Rate Measurement
To investigate the influence of OMSV on mycelial growth in P. floridanus, OMSV-free and OMSV-infected strains were individually inoculated at the central point of PDA (9 cm in diameter), which were incubated in darkness at 25 °C. Following inoculation, the vertical line was made with the center of the mycelium block as the intersection point. It took about 7 days to record growth length of mycelium every day until the colony diameters filled the plate. Concurrently, the average mycelium growth rate (mm/day) was calculated. All samples were repeated in triplicate.
4.7. Cultivation of P. floridanus
A 7.5 mm activated mycelium was cut with a sterilized cylindrical cutter and placed in a 200 mL potato glucose broth (PDB). The culture was maintained in darkness at 25 °C with agitation at 150 rpm for a 10-day period. The basic cultivation substrate consisted of 78% cottonseed shells, 20% wheat bran, and 2% pulverized lime with 60% water saturation and was packed in polypropylene bags (weighing 1.5 kg). Before the substrate mixing, cottonseed shells were immersed in room temperature tap water for 12 h. The moisture content was verified by manually squeezing the substrate until no further water exuded. The blended substrate was sterilized at 121 °C for 3 h and then cooled to 20 °C. Each bag was inoculated with 20 mL P. floridanus mycelium which had been previously cultivated in PDB. The bags were then placed in a dark cultivation chamber maintained at 24 °C. When the mycelium filled the whole bag, the bag was transferred to the fruiting room chamber for culture. The bags were carefully opened by making a small incision with a knife. The growth chamber was aerated by periodically opening the door and maintained a relative humidity of 80–85% by humidifier, while the temperature was controlled at 22–24 °C via an air conditioner. The fully mature fruiting bodies were harvested 7 days after the formation of the primordium. During the cropping period, the mushroom flushes were harvested twice, with the cumulative yield from 60 bags of P. floridanus being documented. Statistical analysis was performed using SPSS (version 26) software. The independent sample t-test was employed to determine the statistical significance of the variations. A p-value < 0.05 was employed for defining a statistically significant difference. Data were reported as mean ± standard deviations.
5. Conclusions
In summary, this study provided strong evidence of OMSV transmission through sexual basidiospores, demonstrating its vertical transmission capability. Furthermore, OMSV was consistently transmitted to progeny via single-spore hybridization, marking the first reported instance of the vertical transmission of OMSV. Meanwhile, OMSV was found to be horizontally transmitted from P. pulmonarius to P. floridanus, resulting in significant reductions in mycelial growth and mushroom yield. These findings enhanced the understanding of the potential threats posed by OMSV to Pleurotus species.
Author Contributions
Conceptualization, X.Z., Y.W. and Z.W.; methodology, Y.Y., X.H., G.S. and Y.W.; investigation, Y.W., Z.W., Z.S., H.H., L.Y., Y.L. and J.W.; data curation, X.Z.; writing—original draft preparation, X.Z. and Y.W.; writing—review and editing X.Z. and Y.W.; funding acquisition, X.Z. and X.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the Yantai Science and Technology Development Project, Shandong Province, China (2023JCYJ087), the Youth Innovation Technology Support Planning Project for Institution of Higher Education of Shandong Province (2022KJ121), the National Natural Science Foundation of China (31900139), and the Edible Fruiting bodies Genetic Breeding Innovation Team of Shandong Agricultural Industry Technology System (SDAIT-07-03).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article.
Acknowledgments
We sincerely thank Chenggui Han (China Agricultural University, China) for his helpful comments on this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ghabrial, S.A.; Caston, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Hollings, M. Viruses Associated with A Die-Back Disease of Cultivated Mushroom. Nature 1962, 196, 962–965. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, J.; Li, Y. Diversity of mycoviruses in edible fungi. Virus Genes 2022, 58, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Son, M.; Yu, J.; Kim, K.-H. Five Questions about Mycoviruses. PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New Insights into Mycoviruses and Exploration for the Biological Control of Crop Fungal Diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef]
- Valverde, R.A.; Gutierrez, D.L. Transmission of a dsRNA in bell pepper and evidence that it consists of the genome of an endornavirus. Virus Genes 2007, 35, 399–403. [Google Scholar] [CrossRef]
- Wang, Q.; Mu, F.; Xie, J.; Cheng, J.; Fu, Y.; Jiang, D. A Single ssRNA Segment Encoding RdRp Is Sufficient for Replication, Infection, and Transmission of Ourmia-Like Virus in Fungi. Front. Microbiol. 2020, 11, 379. [Google Scholar] [CrossRef]
- Ghabrial, S.A. Origin, Adaptation and Evolutionary Pathways of Fungal Viruses. Virus Genes 1998, 16, 119–131. [Google Scholar] [CrossRef] [PubMed]
- van Heerden, S.W.; Geletka, L.M.; Preisig, O.; Nuss, D.L.; Wingfield, B.D.; Wingfield, M.J. Characterization of south African Cryphonectria cubensis isolates infected with a C. parasitica hypovirus. Phytopathology 2001, 91, 628–632. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Li, P.; Qiu, D.; Guo, L. Transcriptome-Based Discovery of Fusarium graminearum Stress Responses to FgHV1 Infection. Int. J. Mol. Sci. 2016, 17, 1922. [Google Scholar] [CrossRef] [PubMed]
- Sun, A.; Zhao, L.; Sun, Y.; Chen, Y.; Li, C.; Dong, W.; Yang, G. Horizontal and Vertical Transmission of a Mycovirus Closely Related to the Partitivirus RhsV717 That Confers Hypovirulence in Rhizoctonia solani. Viruses 2023, 15, 2088. [Google Scholar] [CrossRef]
- Romaine, C.P.; Schlagnhaufer, P.U. Transmission of La France isometric virus during basidiosporogenesis in Agaricus bisporus. Mycologia 1993, 85, 175. [Google Scholar] [CrossRef]
- Guo, M.; Shen, G.; Wang, J.; Liu, M.; Bian, Y.; Xu, Z. Mycoviral diversity and characteristics of a negative-stranded RNA virus LeNSRV1 in the edible mushroom Lentinula edodes. Virology 2021, 555, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Won, H.-K.; Park, S.-J.; Kim, D.-K.; Shin, M.J.; Kim, N.; Lee, S.-H.; Kwon, Y.-C.; Ko, H.K.; Ro, H.-S.; Lee, H.-S. Isolation and characterization of a mycovirus in Lentinula edodes. J. Microbiol. 2013, 51, 118–122. [Google Scholar] [CrossRef]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, W.; Lu, Y.; Kang, Q.; Sui, L.; Liu, H.; Zhao, Y.; Zou, X.; Li, Q. Hypovirulence-associated mycovirus epidemics cause pathogenicity degeneration of Beauveria bassiana in the field. Virol. J. 2023, 20, 255. [Google Scholar] [CrossRef]
- Cornejo, C.; Hisano, S.; Bragança, H.; Suzuki, N.; Rigling, D. A New Double-Stranded RNA Mycovirus in Cryphonectria naterciae Is Able to Cross the Species Barrier and Is Deleterious to a New Host. J. Fungi 2021, 7, 861. [Google Scholar] [CrossRef]
- Liu, Y.; Linder-Basso, D.; Hillman, B.I.; Kaneko, S.; Milgroom, M.G. Evidence for interspecies transmission of viruses in natural populations of filamentous fungi in the genus Cryphonectria. Mol. Ecol. 2003, 12, 1619–1628. [Google Scholar] [CrossRef]
- O’Connor, E.; Doyle, S.; Amini, A.; Grogan, H.; Fitzpatrick, D.A. Transmission of mushroom virus X and the impact of virus infection on the transcriptomes and proteomes of different strains of Agaricus bisporus. Fungal Biol. 2021, 125, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Shi, N.; Wang, P.; Zhang, Y.; Peng, F.; Yang, G.; Huang, B. A Novel Gammapartitivirus That Causes Changes in Fungal Development and Multi-Stress Tolerance to Important Medicinal Fungus Cordyceps chanhua. J. Fungi 2022, 8, 1309. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.J.; Lim, D.; Lee, H.-S. Characterization of a novel single-stranded RNA mycovirus in Pleurotus ostreatus. Virology 2003, 314, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Dai, J.; Zhang, Y.J.; Li, Y. Cloning of Coat Protein Gene of Oyster Mushroom Spherical Virus and Preliminary Analysis of Viral Pathogenicity. J. Northeast. Agric. Sci. 2022, 47, 43–47. [Google Scholar]
- Shi, Y.C.; Xie, A.T.; Wang, X.L.; Wang, S.J.; Liu, X.D.; Zhou, T. Detection of oyster mushroom spherical virus in Beijing and its controlling measures. China Plant Prot. 2016, 36, 9–11+21+81. [Google Scholar]
- Hu, H.-J.; Wang, J.-R.; Cheng, X.-H.; Liu, Y.; Zhang, X.-Y. Preliminary Studies on the Effects of Oyster Mushroom Spherical Virus China Strain on the Mycelial Growth and Fruiting Body Yield of the Edible Mushroom Pleurotus ostreatus. Biology 2022, 11, 574. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, H.; Zhao, Y.; Wang, Y.; Zhang, W.; You, L.; Wang, J.; Liu, Y.; Cheng, X. Oyster Mushroom Spherical Virus Crosses the Species Barrier and Is Pathogenic to a New Host Pleurotus pulmonarius. Int. J. Mol. Sci. 2023, 24, 10584. [Google Scholar] [CrossRef]
- Khan, H.A.; Sato, Y.; Kondo, H.; Jamal, A.; Bhatti, M.F.; Suzuki, N. A second capsidless hadakavirus strain with 10 positive-sense single-stranded RNA genomic segments from Fusarium nygamai. Arch. Virol. 2021, 166, 2711–2722. [Google Scholar] [CrossRef]
- Khan, H.A.; Shamsi, W.; Jamal, A.; Javaied, M.; Sadiq, M.; Fatma, T.; Ahmed, A.; Arshad, M.; Waseem, M.; Babar, S.; et al. Assessment of mycoviral diversity in Pakistani fungal isolates revealed infection by 11 novel viruses of a single strain of Fusarium mangiferae isolate SP1. J. Gen. Virol. 2021, 102, 001690. [Google Scholar] [CrossRef]
- Sato, Y.; Shamsi, W.; Jamal, A.; Bhatti, M.F.; Kondo, H.; Suzuki, N. Hadaka Virus 1: A capsidless eleven-segmented positive-sense single-stranded RNA virus from a phytopathogenic fungus, Fusarium oxysporum. mBio 2020, 11, e00450-20. [Google Scholar] [CrossRef]
- Yaegashi, H.; Sawahata, T.; Ito, T.; Kanematsu, S. A novel colony-print immunoassay reveals differential patterns of distribution and horizontal transmission of four unrelated mycoviruses in Rosellinia necatrix. Virology 2011, 409, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhou, K.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. Viral cross-class transmission results in disease of a phytopathogenic fungus. ISME J. 2022, 16, 2763–2774. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.; Chiba, S.; Kondo, H.; Suzuki, N. Cryphonectria nitschkei chrysovirus 1 with unique molecular features and a very narrow host range. Virology 2020, 554, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Yan, Z.-F.; Kook, M.; Li, C.-T.; Yi, T.-H. Genetic and Chemical Diversity of Edible Mushroom Pleurotus Species. BioMed Res. Int. 2022, 2022, 6068185. [Google Scholar] [CrossRef] [PubMed]
- Romaine, C.; Goodin, M. Unraveling the viral complex associated with La France disease of the cultivated mushroom, Agaricus bisporus. In dsRNA Genetic Elements; CRC Press: Boca Raton, FL, USA, 2001; pp. 237–257. [Google Scholar]
- Rao, J.R.; Nelson, D.W.A.; McClean, S. The Enigma of Double-stranded RNA (dsRNA) Associated with Mushroom Virus X (MVX). Curr. Issues Mol. Biol. 2007, 9, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Bian, Y.; Wang, J.; Wang, G.; Ma, X.; Xu, Z. Biological and Molecular Characteristics of a Novel Partitivirus Infecting the Edible Fungus Lentinula edodes. Plant Dis. 2017, 101, 726–733. [Google Scholar] [CrossRef]
- Song, H.-Y.; Kim, N.; Kim, D.-H.; Kim, J.-M. The PoV mycovirus affects extracellular enzyme expression and fruiting body yield in the oyster mushroom, Pleurotus ostreatus. Sci. Rep. 2020, 10, 1094. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).