
Electrophysiological Effects of the Sodium-Glucose Co-Transporter-2 (SGLT2) Inhibitor Dapagliflozin on Human Cardiac Potassium Channels

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
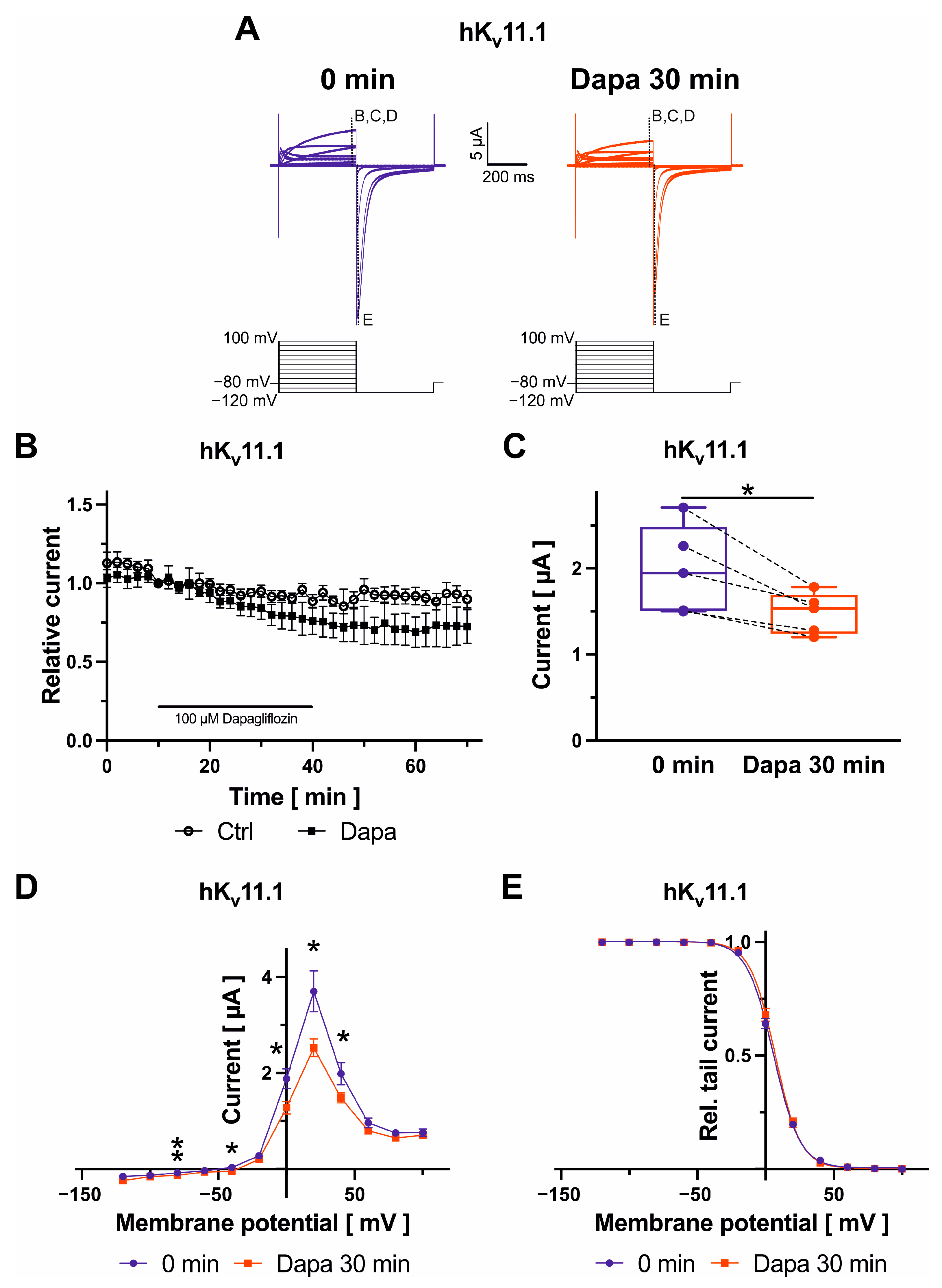
2.1. Effects of Dapagliflozin on Kv11.1 Channels
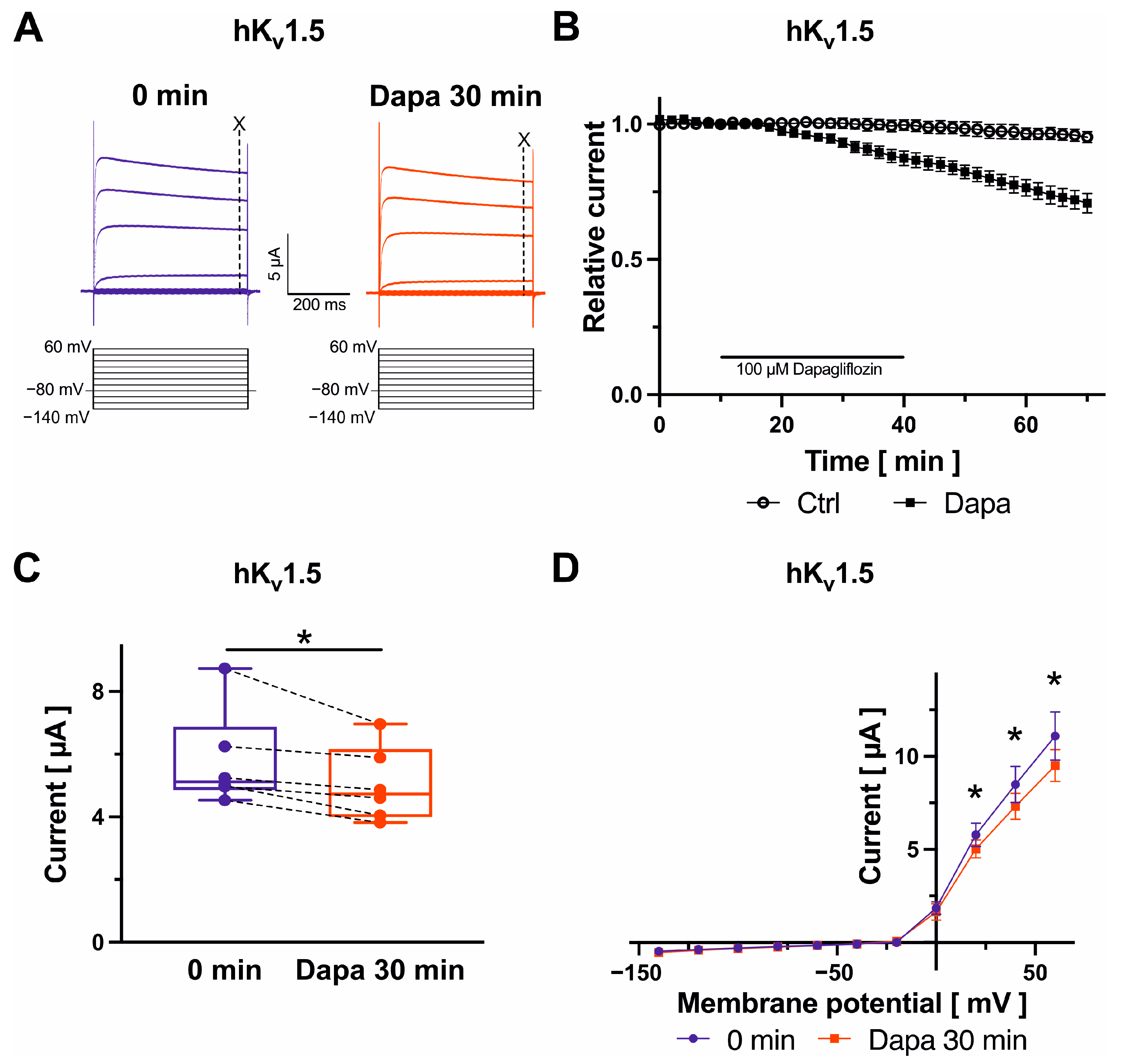
2.2. Effects on Kv1.5 Channels
2.3. Effects on Kv4.3 Channels
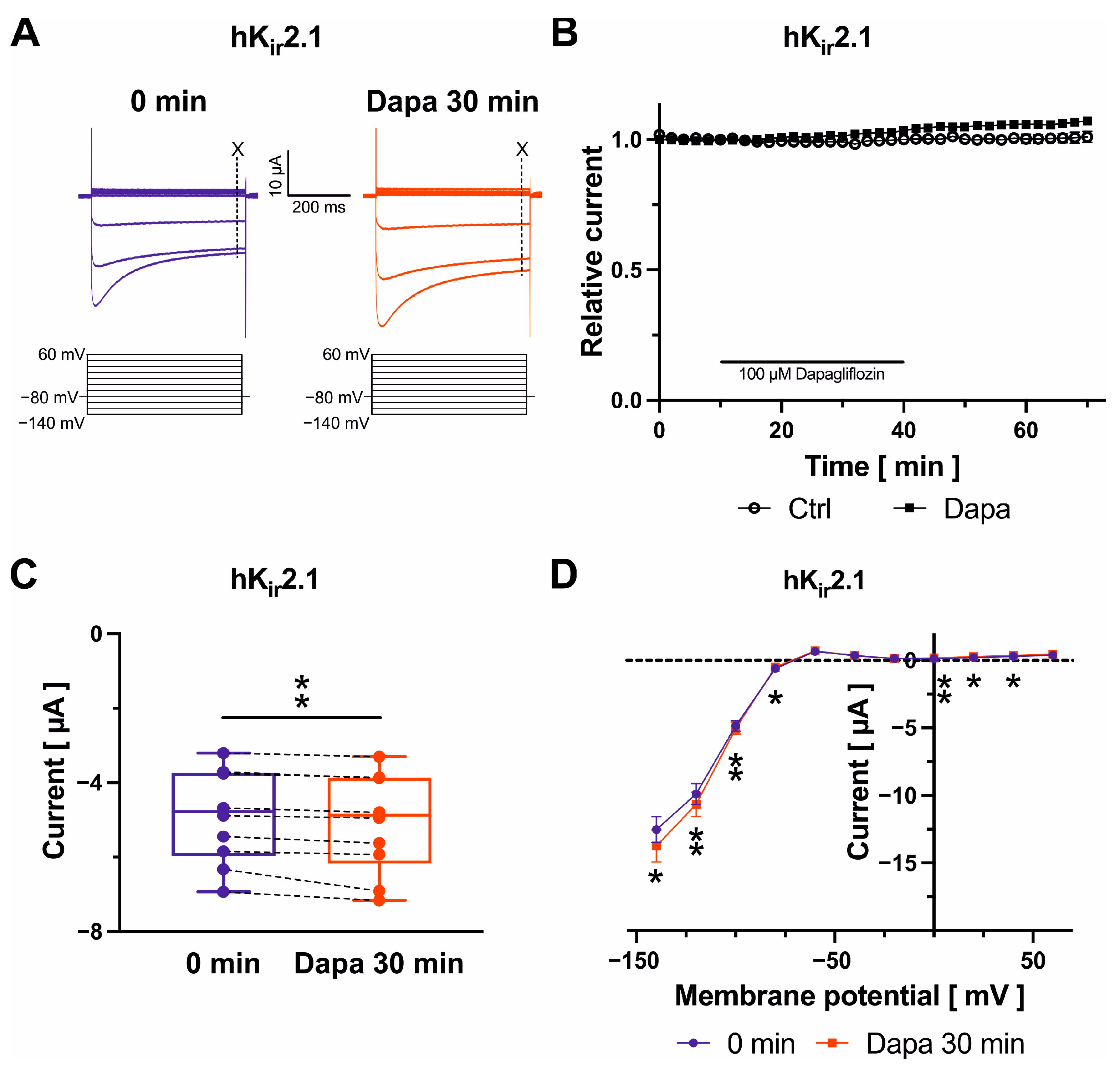
2.4. Effects on Kir2.1 Channels
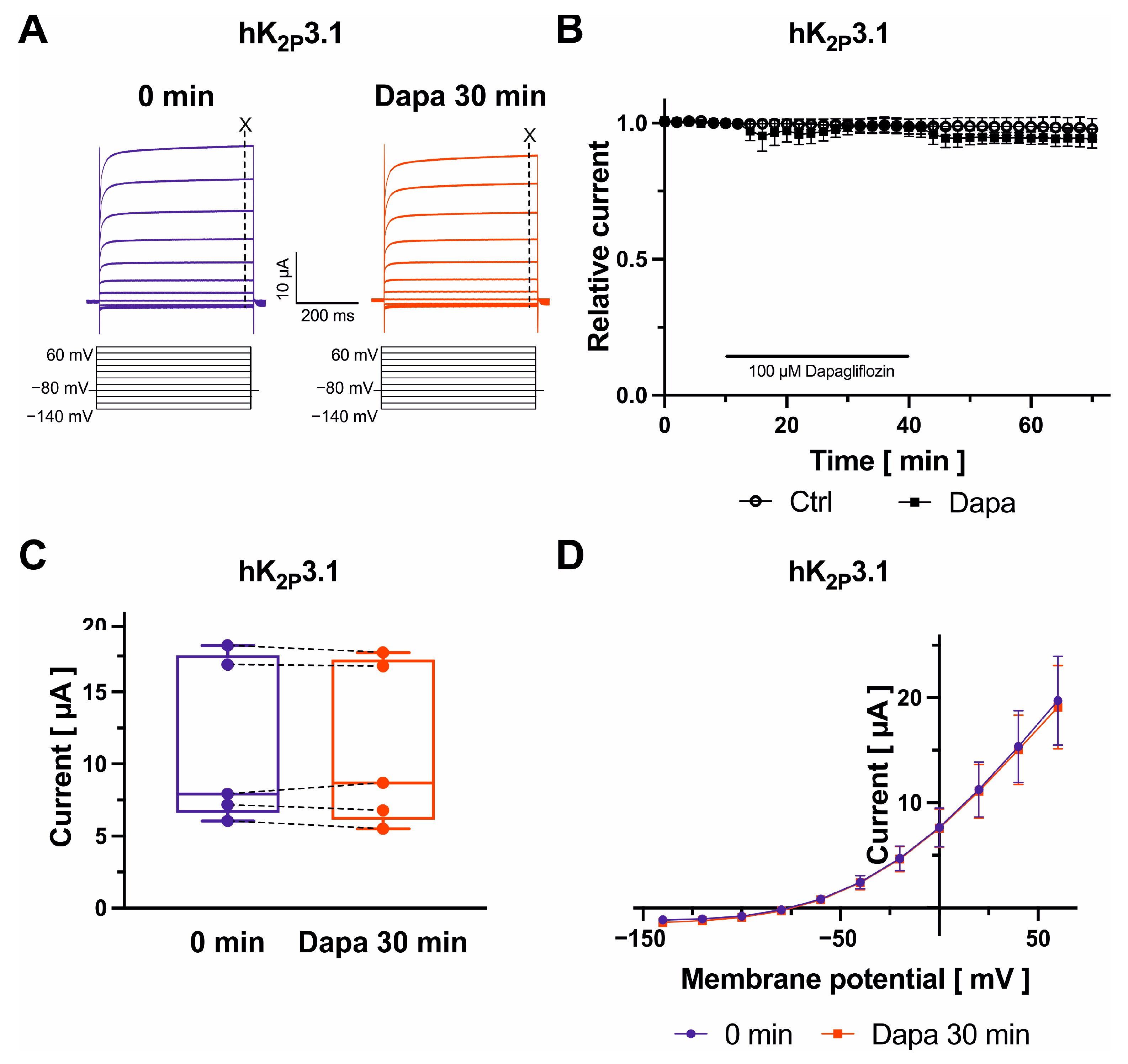
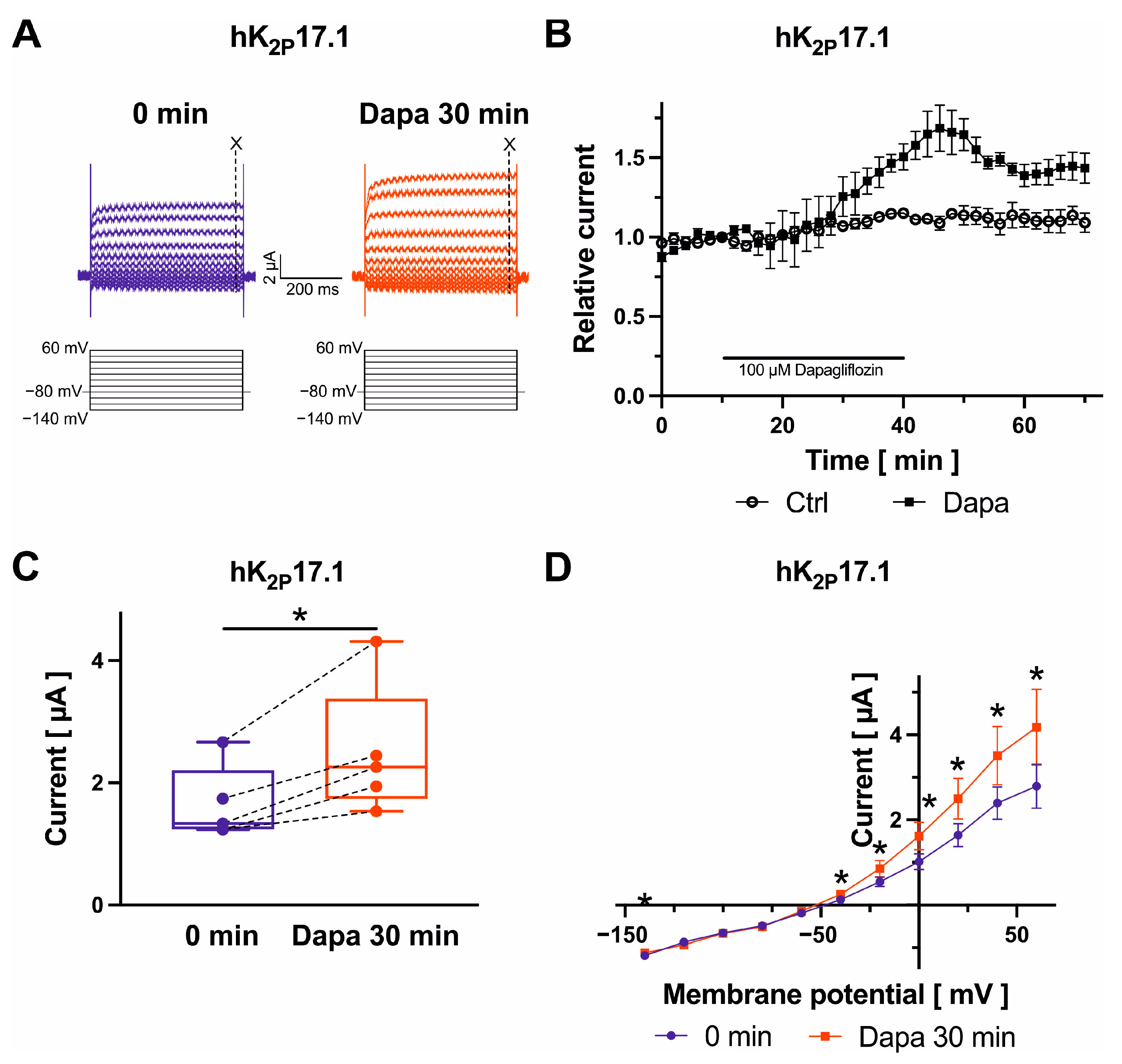
2.5. Dapagliflozin Effects on K2P2.1, K2P3.1, and K2P17.1 Channels
3. Discussion
Limitations
4. Materials and Methods
4.1. Molecular Biology
4.2. Xenopus Laevis Oocyte Preparation
4.3. Electrophysiology
4.4. Drugs
4.5. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chugh, S.S.; Havmoeller, R.; Narayanan, K.; Singh, D.; Rienstra, M.; Benjamin, E.J.; Gillum, R.F.; Kim, Y.H.; McAnulty JHJr Zheng, Z.J.; Forouzanfar, M.H.; et al. Worldwide epidemiology of atrial fibrillation: A Global Burden of Disease 2010 Study. Circulation 2014, 129, 837–847. [Google Scholar] [CrossRef]
- Kornej, J.; Börschel, C.S.; Benjamin, E.J.; Schnabel, R.B. Epidemiology of Atrial Fibrillation in the 21st Century: Novel Methods and New Insights. Circ. Res. 2020, 127, 4–20. [Google Scholar] [CrossRef]
- Wodchis, W.P.; Bhatia, R.S.; Leblanc, K.; Meshkat, N.; Morra, D. A review of the cost of atrial fibrillation. Value Health 2012, 15, 240–248. [Google Scholar] [CrossRef]
- Heijman, J.; Hohnloser, S.H.; Camm, A.J. Antiarrhythmic drugs for atrial fibrillation: Lessons from the past and opportunities for the future. Europace 2021, 23 (Suppl. 2), ii14–ii22. [Google Scholar] [CrossRef]
- Hindricks, G.; Potpara, T.; Dagres, N.; Arbelo, E.; Bax, J.J.; Blomström-Lundqvist, C.; Boriani, G.; Castella, M.; Dan, G.-A.; Dilaveris, P.E.; et al. 2020 ESC Guidelines for the diagnosis and management of atrial fibrillation developed in collaboration with the European Association for Cardio-Thoracic Surgery (EACTS): The Task Force for the diagnosis and management of atrial fibrillation of the European Society of Cardiology (ESC) Developed with the special contribution of the European Heart Rhythm Association (EHRA) of the ESC. Eur. Heart J. 2021, 42, 373–498. [Google Scholar] [CrossRef]
- Zelniker, T.A.; Bonaca, M.P.; Furtado, R.H.M.; Mosenzon, O.; Kuder, J.F.; Murphy, S.A.; Bhatt, D.L.; Leiter, L.A.; McGuire, D.K.; Wilding, J.P.H.; et al. Effect of dapagliflozin on atrial fibrillation in patients with type 2 diabetes mellitus: Insights from the DECLARE-TIMI 58 trial. Circulation 2020, 141, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, Y.; Hidru, T.H.; Yang, X.; Wang, Y.; Chen, C.; Li, K.H.C.; Tang, Y.; Wei, Y.; Tse, G.; et al. Protective effects of sodium-glucose transporter 2 inhibitors on atrial fibrillation and atrial flutter: A systematic review and meta- analysis of randomized placebo-controlled trials. Front. Endocrinol. 2021, 12, 619586. [Google Scholar] [CrossRef] [PubMed]
- Durak, A.; Olgar, Y.; Degirmenci, S.; Akkus, E.; Tuncay, E.; Turan, B. A SGLT2 inhibitor dapagliflozin suppresses prolonged ventricular-repolarization through augmentation of mitochondrial function in insulin-resistant metabolic syndrome rats. Cardiovasc. Diabetol. 2018, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shin, S.E.; Seo, M.S.; An, J.R.; Choi, I.W.; Jung, W.K.; Firth, A.L.; Lee, D.S.; Yim, M.J.; Choi, G.; et al. The anti-diabetic drug dapagliflozin induces vasodilation via activation of PKG and Kv channels. Life Sci. 2018, 197, 46–55. [Google Scholar] [CrossRef]
- Dago, M.; Crespo-García, T.; Cámara-Checa, A.; Rapún, J.; Rubio-Alarcón, M.; Marín, M.; Tamargo, J.; Caballero, R.; Delpón, E. Empagliflozin and dapagliflozin increase Na+ and inward rectifier K+ current densities in human cardiomyocytes derived from induced puripotent stem cells (hiPSC-CMs). Cells 2022, 11, 3707. [Google Scholar] [CrossRef]
- Paasche, A.; Wiedmann, F.; Kraft, M.; Seibertz, F.; Herlt, V.; Blochberger, P.L.; Jávorszky, N.; Beck, M.; Weirauch, L.; Seeger, T.; et al. Acute antiarrhythmic effects of SGLT2 inhibitors-dapagliflozin lowers the excitability of atrial cardiomyocytes. Basic Res. Cardiol. 2024, 119, 93–112. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Böhm, M.; Burri, H.; Butler, J.; Čelutkienė, J.; Chioncel, O.; et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: Developed by the Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC). With the special contribution of the Heart Failure Association (HFA) of the ESC. Eur. Heart J. 2022, 24, 4–131. [Google Scholar] [CrossRef] [PubMed]
- Mc Donagh, T.A.; Metra, M.; Adamo, M.; Gardner, R.S.; Baumbach, A.; Böhm, M.; Burri, H.; Butler, J.; Čelutkienė, J.; Chioncel, O.; et al. 2023 Focused Update of the 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2023, 44, 3627–3639. [Google Scholar] [CrossRef] [PubMed]
- Heijman, J.; Guichard, J.B.; Dobrev, D.; Nattel, S. Translational challenges in atrial fibrillation. Circ. Res. 2018, 122, 752–773. [Google Scholar] [CrossRef] [PubMed]
- Decher, N.; Rinné, S.; Bedoya, M.; Gonzalez, W.; Kiper, A.K. Molecular pharmacology of K2P potassium channels. Cell. Physiol. Biochem. 2021, 55, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Kisselbach, J.; Seyler, C.; Schweizer, P.A.; Gerstberger, R.; Becker, R.; Katus, H.A.; Thomas, D. Modulation of K2P2.1 and K2P10.1 K+ channel sensitivity to carvedilol by alternative mRNA translation initiation. Br. J. Pharmacol. 2014, 171, 5182–5194. [Google Scholar] [CrossRef]
- Lugenbiel, P.; Wenz, F.; Syren, P.; Geschwill, P.; Govorov, K.; Seyler, C.; Frank, D.; Schweizer, P.A.; Franke, J.; Weis, T.; et al. TREK-1 (K2P2.1) K+ channels are suppressed in patients with atrial fibrillation and heart failure and provide therapeutic targets for rhythm control. Basic Res. Cardiol. 2017, 112, 8. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Wiedmann, F.; Voigt, N.; Zhou, X.B.; Heijman, J.; Lang, S.; Albert, V.; Kallenberger, S.; Ruhparwar, A.; Szabó, G.; et al. Upregulation of K2P3.1 K+ current causes action potential shortening in patients with chronic atrial fibrillation. Circulation 2015, 132, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Seyler, C.; Schweizer, P.A.; Zitron, E.; Katus, H.A.; Thomas, D. Vernakalant activates human cardiac K2P17.1 background K+ channels. Biochem. Biophys. Res. Commun. 2014, 451, 415–420. [Google Scholar] [CrossRef]
- Staudacher, I.; Illg, C.; Chai, S.; Deschenes, I.; Seehausen, S.; Gramlich, D.; Müller, M.E.; Wieder, T.; Rahm, A.K.; Mayer, C.; et al. Cardiovascular pharmacology of K2P17.1 (TASK-4, TALK-2) two-pore-domain K+ channels. Naunyn Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1119–1131. [Google Scholar] [CrossRef]
- Schmidt, C.; Wiedmann, F.; Zhou, X.B.; Heijman, J.; Voigt, N.; Ratte, A.; Lang, S.; Kallenberger, S.M.; Campana, C.; Weymann, A.; et al. Inverse remodelling of K2P3.1 K+ channel expression and action potential duration in left ventricular dysfunction and atrial fibrillation: Implications for patient-specific antiarrhythmic drug therapy. Eur. Heart J. 2017, 38, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, C.; Rinné, S.; Zumhagen, S.; Kiper, A.K.; Silbernagel, N.; Netter, M.F.; Stallmeyer, B.; Schulze-Bahr, E.; Decher, N. Gain-of-function mutation in TASK-4 channels and severe cardiac conduction disorder. EMBO Mol. Med. 2014, 6, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.; Wan, X.; Nassal, D.M.; Liu, H.; Moravec, C.S.; Ramirez-Navarro, A.; Deschênes, I. Contribution of two-pore K+ channels to cardiac ventricular action potential revealed using human iPSC-derived cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1144–H1153. [Google Scholar] [CrossRef] [PubMed]
- Schotten, U.; Verheule, S.; Kirchhof, P.; Goette, A. Pathophysiological mechanisms of atrial fibrillation: A translational appraisal. Physiol. Rev. 2011, 91, 265–325. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.C.; Tristani-Firouzi, M. hERG potassium channels and cardiac arrhythmia. Nature 2006, 440, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Carlson, G.F.; Tou, C.K.; Parikh, S.; Birmingham, B.K.; Butler, K. Evaluation of the effect of dapagliflozin on cardiac repolarization: A thorough QT/QTc study. Diabetes Ther. 2011, 2, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Nakase, M.; Yahagi, K.; Horiuchi, Y.; Asami, M.; Yuzawa, H.; Komiyama, K.; Tanaka, J.; Aoki, J.; Tanabe, K. Effect of dapagliflozin on ventricular repolarization in patients with heart failure with reduced ejection fraction. Heart Vessels 2023, 38, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Mays, D.J.; Foose, J.M.; Philipson, L.H.; Tamkun, M.M. Localization of the Kv1.5 K+ channel protein in explanted cardiac tissue. J. Clin. Investig. 1995, 96, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wible, B.; Li, G.R.; Wang, Z.; Nattel, S. Antisense oligodeoxynucleotides directed against Kv1.5 mRNA specifically inhibit ultrarapid delayed rectifier K+ current in cultured adult human atrial myocytes. Circ. Res. 1997, 80, 572–579. [Google Scholar] [CrossRef]
- Van Wagoner, D.R.; Pond, A.L.; McCarthy, P.M.; Trimmer, J.S.; Nerbonne, J.M. Outward K+ current densities and Kv1.5 expression are reduced in chronic human atrial fibrillation. Circ. Res. 1997, 80, 772–781. [Google Scholar] [CrossRef]
- Olson, T.M.; Alekseev, A.E.; Liu, X.K.; Park, S.; Zingman, L.V.; Bienengraeber, M.; Sattiraju, S.; Ballew, J.D.; Jahangir, A.; Terzic, A. Kv1.5 channelopathy due to KCNA5 loss-of-function mutation causes human atrial fibrillation. Hum. Mol. Genet. 2006, 15, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Kasichayanula, S.; Liu, X.; Lacreta, F.; Griffen, S.C.; Boulton, D.W. Clinical pharmacokinetics and pharmacodynamics of dapagliflozin, a selective inhibitor of sodium-glucose co-transporter type 2. Clin. Pharmacokinet. 2014, 53, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Gierten, J.; Ficker, E.; Bloehs, R.; Schlömer, K.; Kathöfer, S.; Scholz, E.; Zitron, E.; Kiesecker, C.; Bauer, A.; Becker, R.; et al. Regulation of two-pore-domain (K2P) potassium leak channels by the tyrosine kinase inhibitor genistein. Br. J. Pharmacol. 2008, 154, 1680–1690. [Google Scholar] [CrossRef]
- Gierten, J.; Ficker, E.; Bloehs, R.; Schweizer, P.A.; Zitron, E.; Scholz, E.; Karle, C.; Katus, H.A.; Thomas, D. The human cardiac K2P3.1 (TASK-1) potassium leak channel is a molecular target for the class III antiarrhythmic drug amiodarone. Naunyn Schmiedeberg’s Arch. Pharmacol. 2010, 381, 261–270. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, M.E.; Petersenn, F.; Hackbarth, J.; Pfeiffer, J.; Gampp, H.; Frey, N.; Lugenbiel, P.; Thomas, D.; Rahm, A.-K. Electrophysiological Effects of the Sodium-Glucose Co-Transporter-2 (SGLT2) Inhibitor Dapagliflozin on Human Cardiac Potassium Channels. Int. J. Mol. Sci. 2024, 25, 5701. https://doi.org/10.3390/ijms25115701
Müller ME, Petersenn F, Hackbarth J, Pfeiffer J, Gampp H, Frey N, Lugenbiel P, Thomas D, Rahm A-K. Electrophysiological Effects of the Sodium-Glucose Co-Transporter-2 (SGLT2) Inhibitor Dapagliflozin on Human Cardiac Potassium Channels. International Journal of Molecular Sciences. 2024; 25(11):5701. https://doi.org/10.3390/ijms25115701
Chicago/Turabian StyleMüller, Mara Elena, Finn Petersenn, Juline Hackbarth, Julia Pfeiffer, Heike Gampp, Norbert Frey, Patrick Lugenbiel, Dierk Thomas, and Ann-Kathrin Rahm. 2024. "Electrophysiological Effects of the Sodium-Glucose Co-Transporter-2 (SGLT2) Inhibitor Dapagliflozin on Human Cardiac Potassium Channels" International Journal of Molecular Sciences 25, no. 11: 5701. https://doi.org/10.3390/ijms25115701
APA StyleMüller, M. E., Petersenn, F., Hackbarth, J., Pfeiffer, J., Gampp, H., Frey, N., Lugenbiel, P., Thomas, D., & Rahm, A.-K. (2024). Electrophysiological Effects of the Sodium-Glucose Co-Transporter-2 (SGLT2) Inhibitor Dapagliflozin on Human Cardiac Potassium Channels. International Journal of Molecular Sciences, 25(11), 5701. https://doi.org/10.3390/ijms25115701