

Extracellular Interactors of the IGF System: Impact on Cancer Hallmarks and Therapeutic Approaches




Abstract
1. Introduction
2. General Overview of the IGF System
3. The IGF System in Cancer: A Crucial Hub at the Crossroads between the Intracellular and Extracellular Compartments
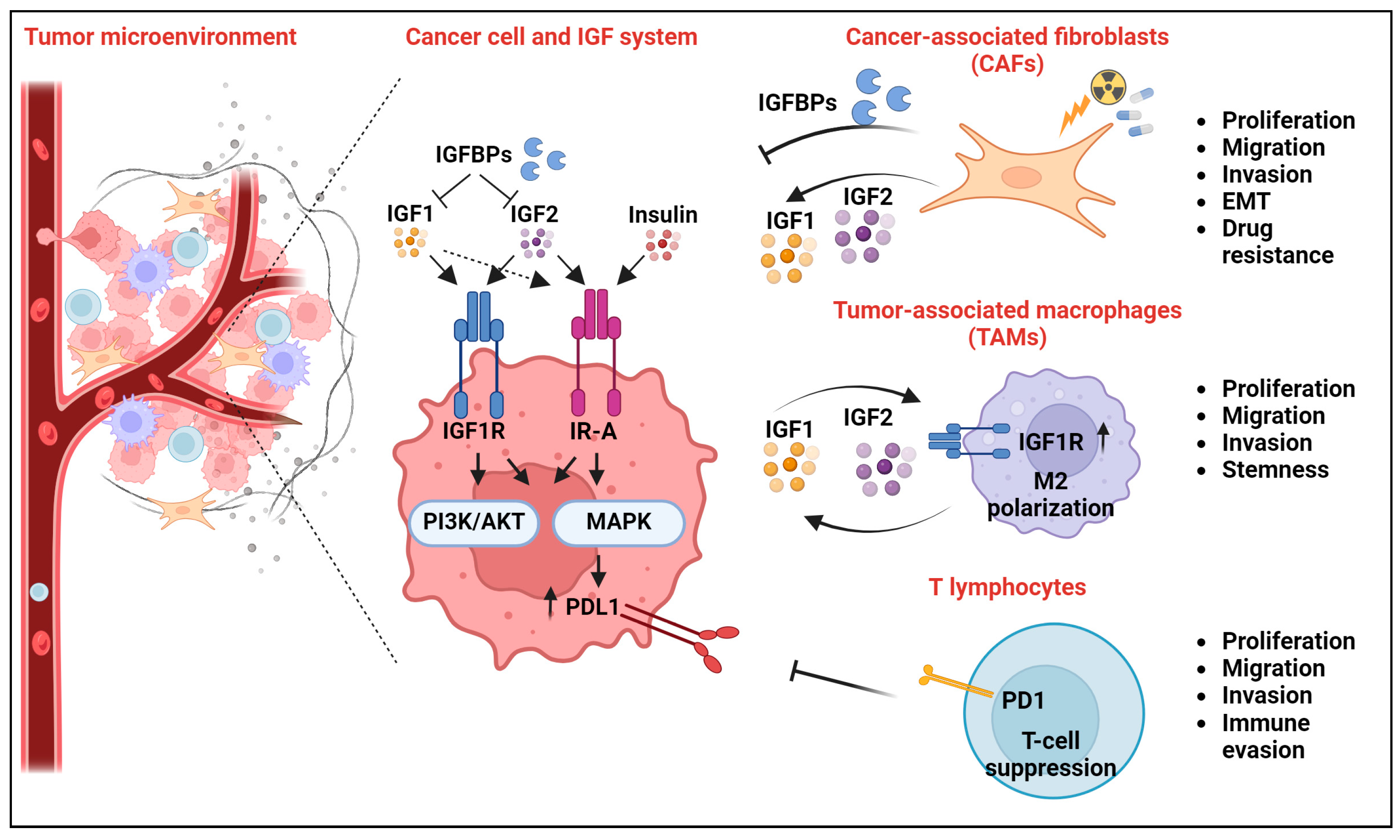
4. Novel Extracellular Interactors of the IGF System and Their Impact on Cancer Hallmarks
4.1. The IGF System and Cancer-Associated Fibroblasts
4.2. The IGF System and Tumor-Associated Macrophages
4.3. The IGF System and T Lymphocytes
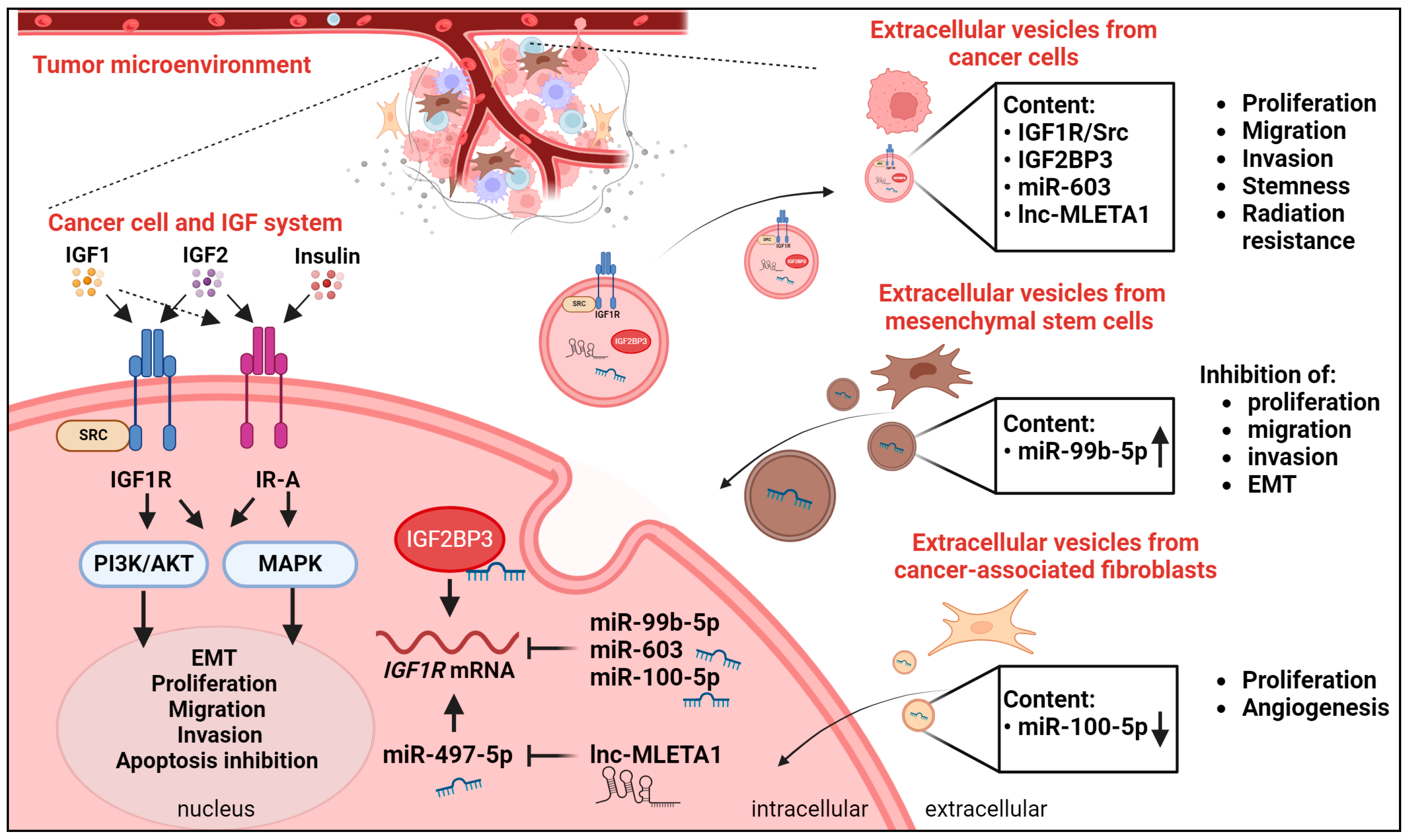
4.4. The IGF System and Extracellular Vesicles
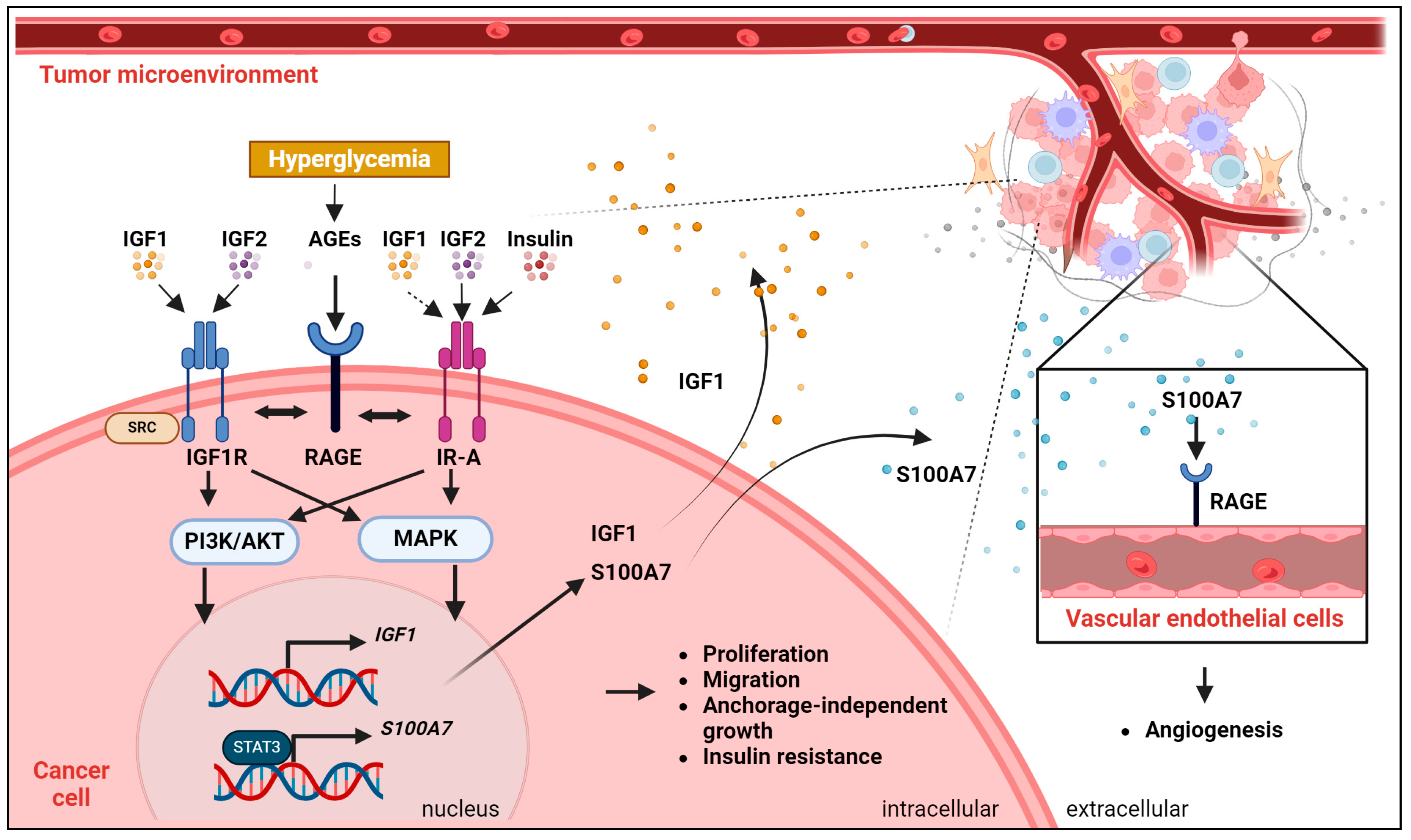
4.5. The IGF System and Glycation
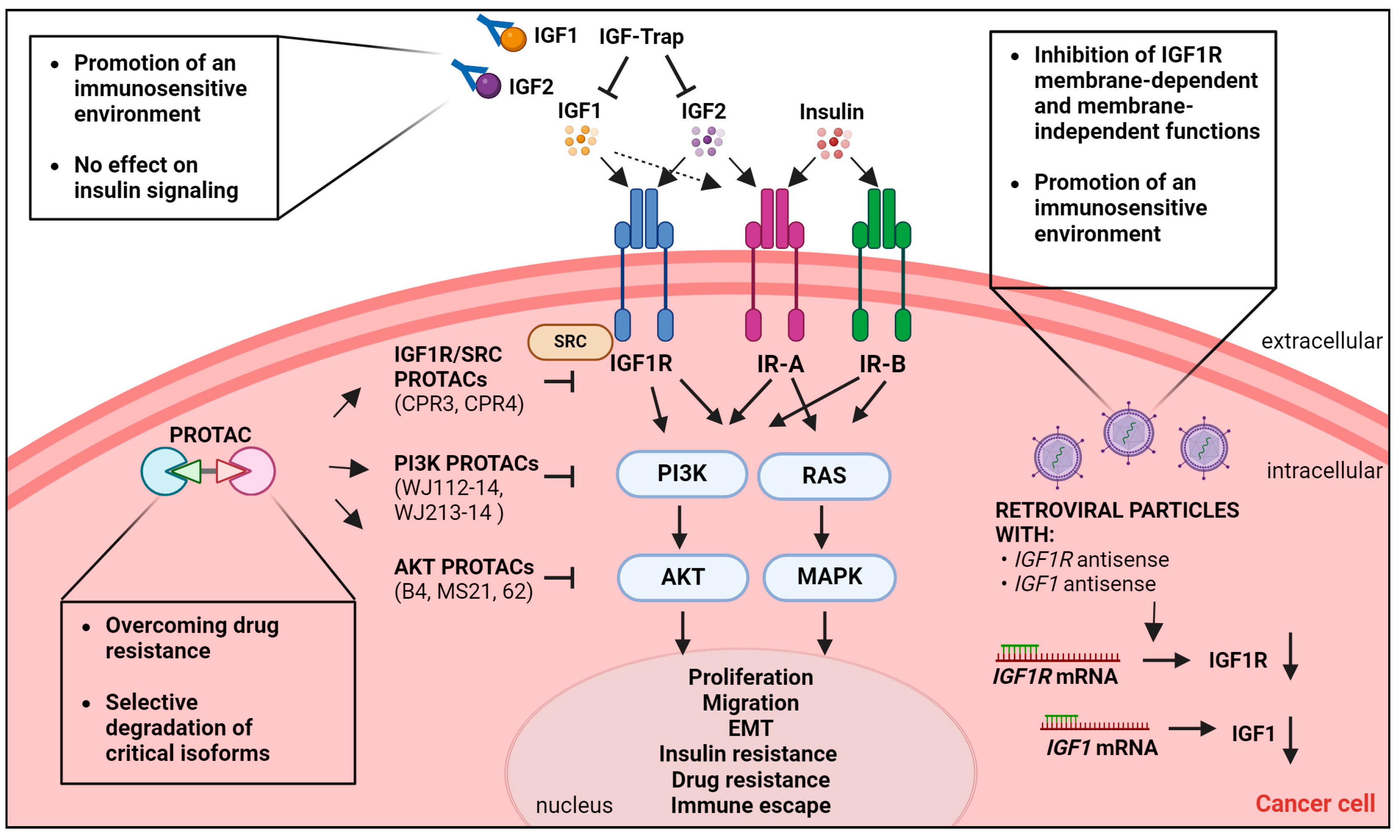
5. Emerging Therapeutic Strategies to Target the IGF System in Cancer
5.1. IGF-Trap
5.2. Gene Therapy
5.3. Targeted Protein Degradation-Based Approaches
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- LeRoith, D.; Holly, J.M.P.; Forbes, B.E. Insulin-like growth factors: Ligands, binding proteins, and receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef] [PubMed]
- Deyev, I.E.; Mitrofanova, A.V.; Zhevlenev, E.S.; Radionov, N.; Berchatova, A.A.; Popova, N.V.; Serova, O.V.; Petrenko, A.G. Structural determinants of the insulin receptor-related receptor activation by alkali. J. Biol. Chem. 2013, 288, 33884–33893. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Hyperinsulinaemia in cancer. Nat. Rev. Cancer 2020, 20, 629–644. [Google Scholar] [CrossRef]
- Blum, W.F.; Alherbish, A.; Alsagheir, A.; El Awwa, A.; Kaplan, W.; Koledova, E.; Savage, M.O. The growth hormone-insulin-like growth factor-I axis in the diagnosis and treatment of growth disorders. Endocr. Connect. 2018, 7, R212–R222. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Osher, E.; Macaulay, V.M. Therapeutic Targeting of the IGF Axis. Cells 2019, 8, 895. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Zhang, Q.; Duan, Q.; Tan, Y.; Sun, T.; Qi, C.; Qin, Y.; Liu, H. Identification of IGF1R mutation as a novel predictor of efficacious immunotherapy in melanoma. J. Transl. Med. 2022, 20, 172. [Google Scholar] [CrossRef]
- Piarulli, G.; Puls, F.; Wangberg, B.; Fagman, H.; Hansson, M.; Nilsson, J.; Arbajian, E.; Mertens, F. Gene fusion involving the insulin-like growth factor 1 receptor in an ALK-negative inflammatory myofibroblastic tumour. Histopathology 2019, 74, 1098–1102. [Google Scholar] [CrossRef]
- Mancarella, C.; Morrione, A.; Scotlandi, K. Novel Regulators of the IGF System in Cancer. Biomolecules 2021, 11, 273. [Google Scholar] [CrossRef]
- Mancarella, C.; Morrione, A.; Scotlandi, K. Unraveling the IGF System Interactome in Sarcomas Exploits Novel Therapeutic Options. Cells 2021, 10, 2075. [Google Scholar] [CrossRef]
- Aiello, A.; Pandini, G.; Sarfstein, R.; Werner, H.; Manfioletti, G.; Vigneri, R.; Belfiore, A. HMGA1 protein is a positive regulator of the insulin-like growth factor-I receptor gene. Eur. J. Cancer 2010, 46, 1919–1926. [Google Scholar] [CrossRef]
- Neuhausen, S.L.; Brummel, S.; Ding, Y.C.; Singer, C.F.; Pfeiler, G.; Lynch, H.T.; Nathanson, K.L.; Rebbeck, T.R.; Garber, J.E.; Couch, F.; et al. Genetic variation in insulin-like growth factor signaling genes and breast cancer risk among BRCA1 and BRCA2 carriers. Breast Cancer Res. 2009, 11, R76. [Google Scholar] [CrossRef]
- Yuen, J.S.; Cockman, M.E.; Sullivan, M.; Protheroe, A.; Turner, G.D.; Roberts, I.S.; Pugh, C.W.; Werner, H.; Macaulay, V.M. The VHL tumor suppressor inhibits expression of the IGF1R and its loss induces IGF1R upregulation in human clear cell renal carcinoma. Oncogene 2007, 26, 6499–6508. [Google Scholar] [CrossRef] [PubMed]
- Idelman, G.; Glaser, T.; Roberts, C.T., Jr.; Werner, H. WT1-p53 interactions in insulin-like growth factor-I receptor gene regulation. J. Biol. Chem. 2003, 278, 3474–3482. [Google Scholar] [CrossRef] [PubMed]
- Meisel Sharon, S.; Pozniak, Y.; Geiger, T.; Werner, H. TMPRSS2-ERG fusion protein regulates insulin-like growth factor-1 receptor (IGF1R) gene expression in prostate cancer: Involvement of transcription factor Sp1. Oncotarget 2016, 7, 51375–51392. [Google Scholar] [CrossRef]
- Werner, H.; Meisel-Sharon, S.; Bruchim, I. Oncogenic fusion proteins adopt the insulin-like growth factor signaling pathway. Mol. Cancer 2018, 17, 28. [Google Scholar] [CrossRef]
- Loganathan, S.N.; Tang, N.; Fleming, J.T.; Ma, Y.; Guo, Y.; Borinstein, S.C.; Chiang, C.; Wang, J. BET bromodomain inhibitors suppress EWS-FLI1-dependent transcription and the IGF1 autocrine mechanism in Ewing sarcoma. Oncotarget 2016, 7, 43504–43517. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Yu, Y.; Bilke, S.; Walker, R.L.; Mayeenuddin, L.H.; Azorsa, D.O.; Yang, F.; Pineda, M.; Helman, L.J.; Meltzer, P.S. Genome-wide identification of PAX3-FKHR binding sites in rhabdomyosarcoma reveals candidate target genes important for development and cancer. Cancer Res. 2010, 70, 6497–6508. [Google Scholar] [CrossRef]
- Peng, R.; Cao, J.; Zhang, C.; Zhou, J.; Su, B.B.; Tu, D.Y.; Jiang, G.Q.; Jin, S.J.; Xu, Y.P.; Bai, D.S. In vivo CRISPR screen identifies LTN1 as a novel tumor suppressor ubiquitinating insulin-like growth factor 2 mRNA-binding protein 1 in hepatocellular carcinoma. Hepatol. Commun. 2023, 7, e0256. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Yin, C.; Lin, K.; Li, Y.; Yang, Q.; Wu, Z.; Du, H.; Ren, D.; Dai, Y.; Peng, X. m(6) A modification of lncRNA PCAT6 promotes bone metastasis in prostate cancer through IGF2BP2-mediated IGF1R mRNA stabilization. Clin. Transl. Med. 2021, 11, e426. [Google Scholar] [CrossRef]
- Mancarella, C.; Pasello, M.; Manara, M.C.; Toracchio, L.; Sciandra, E.F.; Picci, P.; Scotlandi, K. Insulin-Like Growth Factor 2 mRNA-Binding Protein 3 Influences Sensitivity to Anti-IGF System Agents Through the Translational Regulation of IGF1R. Front. Endocrinol. (Lausanne.) 2018, 9, 178. [Google Scholar] [CrossRef]
- Martin, A.; Fernandez, M.C.; Cattaneo, E.R.; Schuster, C.D.; Venara, M.; Clement, F.; Berenstein, A.; Lombardi, M.G.; Bergada, I.; Gutierrez, M.; et al. Type 1 Insulin-Like Growth Factor Receptor Nuclear Localization in High-Grade Glioma Cells Enhances Motility, Metabolism, and In Vivo Tumorigenesis. Front. Endocrinol. (Lausanne.) 2022, 13, 849279. [Google Scholar] [CrossRef]
- Yoshihiro, T.; Ariyama, H.; Yamaguchi, K.; Imajima, T.; Yamaga, S.; Tsuchihashi, K.; Isobe, T.; Kusaba, H.; Akashi, K.; Baba, E. Inhibition of insulin-like growth factor-1 receptor enhances eribulin-induced DNA damage in colorectal cancer. Cancer Sci. 2022, 113, 4207–4218. [Google Scholar] [CrossRef]
- Mills, J.V.; Osher, E.; Rieunier, G.; Mills, I.G.; Macaulay, V.M. IGF-1R nuclear import and recruitment to chromatin involves both alpha and beta subunits. Discov. Oncol. 2021, 12, 13. [Google Scholar] [CrossRef]
- Li, Y.; Li, K.; Pan, T.; Xie, Q.; Cheng, Y.; Wu, X.; Xu, R.; Liu, X.; Liu, L.; Gao, J.; et al. Translocation of IGF-1R in endoplasmic reticulum enhances SERCA2 activity to trigger Ca(2+)(ER) perturbation in hepatocellular carcinoma. Acta Pharm. Sin. B 2023, 13, 3744–3755. [Google Scholar] [CrossRef] [PubMed]
- Rieger, L.; O’Shea, S.; Godsmark, G.; Stanicka, J.; Kelly, G.; O’Connor, R. IGF-1 receptor activity in the Golgi of migratory cancer cells depends on adhesion-dependent phosphorylation of Tyr(1250) and Tyr(1251). Sci. Signal. 2020, 13, eaba3176. [Google Scholar] [CrossRef]
- Manara, M.C.; Terracciano, M.; Mancarella, C.; Sciandra, M.; Guerzoni, C.; Pasello, M.; Grilli, A.; Zini, N.; Picci, P.; Colombo, M.P.; et al. CD99 triggering induces methuosis of Ewing sarcoma cells through IGF-1R/RAS/Rac1 signaling. Oncotarget 2016, 7, 79925–79942. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Ke, J.R.; Yu, Z.; Shi, R.; Gao, S.S.; Li, J.F.; Gao, Z.X.; Ke, C.S.; Han, H.X.; et al. Pro-prion, as a membrane adaptor protein for E3 ligase c-Cbl, facilitates the ubiquitination of IGF-1R, promoting melanoma metastasis. Cell Rep. 2022, 41, 111834. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yu, Y.; Wang, W.; Jiang, Y.; Li, Y.; Jiang, X.; Qiao, Y.; Chen, L.; Zhao, X.; Liu, J.; et al. Targeting the E3 ligase NEDD4 as a novel therapeutic strategy for IGF1 signal pathway-driven gastric cancer. Oncogene 2023, 42, 1072–1087. [Google Scholar] [CrossRef] [PubMed]
- Vecchione, A.; Marchese, A.; Henry, P.; Rotin, D.; Morrione, A. The Grb10/Nedd4 complex regulates ligand-induced ubiquitination and stability of the insulin-like growth factor I receptor. Mol. Cell. Biol. 2003, 23, 3363–3372. [Google Scholar] [CrossRef]
- Monami, G.; Emiliozzi, V.; Morrione, A. Grb10/Nedd4-mediated multiubiquitination of the insulin-like growth factor receptor regulates receptor internalization. J. Cell Physiol. 2008, 216, 426–437. [Google Scholar] [CrossRef]
- Qiu, N.; He, Y.F.; Zhang, S.M.; Zhan, Y.T.; Han, G.D.; Jiang, M.; He, W.X.; Zhou, J.; Liang, H.L.; Ao, X.; et al. Cullin7 enhances resistance to trastuzumab therapy in Her2 positive breast cancer via degrading IRS-1 and downregulating IGFBP-3 to activate the PI3K/AKT pathway. Cancer Lett. 2019, 464, 25–36. [Google Scholar] [CrossRef]
- Avino, S.; De Marco, P.; Cirillo, F.; Santolla, M.F.; De Francesco, E.M.; Perri, M.G.; Rigiracciolo, D.; Dolce, V.; Belfiore, A.; Maggiolini, M.; et al. Stimulatory actions of IGF-I are mediated by IGF-IR cross-talk with GPER and DDR1 in mesothelioma and lung cancer cells. Oncotarget 2016, 7, 52710–52728. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, R.; Nicolosi, M.L.; Sacco, A.; Morcavallo, A.; Vella, V.; Voci, C.; Spatuzza, M.; Xu, S.Q.; Iozzo, R.V.; Vigneri, R.; et al. Novel cross talk between IGF-IR and DDR1 regulates IGF-IR trafficking, signaling and biological responses. Oncotarget 2015, 6, 16084–16105. [Google Scholar] [CrossRef] [PubMed]
- Vella, V.; Giuliano, M.; Nicolosi, M.L.; Majorana, M.G.; Marc, M.A.; Muoio, M.G.; Morrione, A.; Maggiolini, M.; Lappano, R.; De Francesco, E.M.; et al. DDR1 Affects Metabolic Reprogramming in Breast Cancer Cells by Cross-Talking to the Insulin/IGF System. Biomolecules 2021, 11, 926. [Google Scholar] [CrossRef] [PubMed]
- Morcavallo, A.; Buraschi, S.; Xu, S.Q.; Belfiore, A.; Schaefer, L.; Iozzo, R.V.; Morrione, A. Decorin differentially modulates the activity of insulin receptor isoform A ligands. Matrix Biol. 2014, 35, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Buraschi, S.; Genua, M.; Xu, S.Q.; Solomides, C.C.; Peiper, S.C.; Gomella, L.G.; Owens, R.C.; Morrione, A. Decorin antagonizes IGF receptor I (IGF-IR) function by interfering with IGF-IR activity and attenuating downstream signaling. J. Biol. Chem. 2011, 286, 34712–34721. [Google Scholar] [CrossRef] [PubMed]
- Morrione, A.; Neill, T.; Iozzo, R.V. Dichotomy of decorin activity on the insulin-like growth factor-I system. FEBS J. 2013, 280, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Galifi, C.A.; Wood, T.L. Insulin-like growth factor-1 receptor crosstalk with integrins, cadherins, and the tumor microenvironment: Sticking points in understanding IGF1R function in cancer. Endocr. Relat. Cancer 2023, 30, e230031. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Tocheny, C.E.; Shaw, L.M. The Insulin-like Growth Factor Signaling Pathway in Breast Cancer: An Elusive Therapeutic Target. Life 2022, 12, 1992. [Google Scholar] [CrossRef]
- Mayer, S.; Milo, T.; Isaacson, A.; Halperin, C.; Miyara, S.; Stein, Y.; Lior, C.; Pevsner-Fischer, M.; Tzahor, E.; Mayo, A.; et al. The tumor microenvironment shows a hierarchy of cell-cell interactions dominated by fibroblasts. Nat. Commun. 2023, 14, 5810. [Google Scholar] [CrossRef]
- Lin, J.; Asai, S.; Selicharova, I.; Mitrova, K.; Kaminsky, J.; Young, E.; Jiracek, J. Recombinant Insulin-Like Growth Factor 1 Dimers: Receptor Binding Affinities and Activation Abilities. Int. J. Pept. Res. Ther. 2023, 29, 33. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, D.; Sun, J.; Wu, Y.; Gong, J.; Li, X.; Liu, L.; Liu, S.; Liu, J.; Wu, Y.; et al. Visualization of Ligand-Bound Ectodomain Assembly in the Full-Length Human IGF-1 Receptor by Cryo-EM Single-Particle Analysis. Structure 2020, 28, 555–561.e4. [Google Scholar] [CrossRef]
- Malaguarnera, R.; Gabriele, C.; Santamaria, G.; Giuliano, M.; Vella, V.; Massimino, M.; Vigneri, P.; Cuda, G.; Gaspari, M.; Belfiore, A. Comparative proteomic analysis of insulin receptor isoform A and B signaling. Mol. Cell. Endocrinol. 2022, 557, 111739. [Google Scholar] [CrossRef]
- Xu, Y.; Margetts, M.B.; Venugopal, H.; Menting, J.G.; Kirk, N.S.; Croll, T.I.; Delaine, C.; Forbes, B.E.; Lawrence, M.C. How insulin-like growth factor I binds to a hybrid insulin receptor type 1 insulin-like growth factor receptor. Structure 2022, 30, 1098–1108.e6. [Google Scholar] [CrossRef]
- Chen, J.; Nagle, A.M.; Wang, Y.F.; Boone, D.N.; Lee, A.V. Controlled dimerization of insulin-like growth factor-1 and insulin receptors reveals shared and distinct activities of holo and hybrid receptors. J. Biol. Chem. 2018, 293, 3700–3709. [Google Scholar] [CrossRef]
- Takeda, T.; Komatsu, M.; Chiwaki, F.; Komatsuzaki, R.; Nakamura, K.; Tsuji, K.; Kobayashi, Y.; Tominaga, E.; Ono, M.; Banno, K.; et al. Upregulation of IGF2R evades lysosomal dysfunction-induced apoptosis of cervical cancer cells via transport of cathepsins. Cell Death Dis. 2019, 10, 876. [Google Scholar] [CrossRef]
- Torrente, Y.; Bella, P.; Tripodi, L.; Villa, C.; Farini, A. Role of Insulin-Like Growth Factor Receptor 2 across Muscle Homeostasis: Implications for Treating Muscular Dystrophy. Cells 2020, 9, 441. [Google Scholar] [CrossRef]
- Goryashchenko, A.S.; Mozhaev, A.A.; Serova, O.V.; Erokhina, T.N.; Orsa, A.N.; Deyev, I.E.; Petrenko, A.G. Probing Structure and Function of Alkali Sensor IRR with Monoclonal Antibodies. Biomolecules 2020, 10, 1060. [Google Scholar] [CrossRef]
- Takahashi, S.I. IGF research 2016–2018. Growth Horm. IGF Res. 2019, 48–49, 65–69. [Google Scholar] [CrossRef]
- Chung, C.L.; Chen, C.L. Fluoroquinolones upregulate insulin-like growth factor-binding protein 3, inhibit cell growth and insulin-like growth factor signaling. Eur. J. Pharmacol. 2024, 969, 176421. [Google Scholar] [CrossRef]
- Kobbero, S.D.; Gajhede, M.; Mirza, O.A.; Kloverpris, S.; Kjaer, T.R.; Mikkelsen, J.H.; Boesen, T.; Oxvig, C. Structure of the proteolytic enzyme PAPP-A with the endogenous inhibitor stanniocalcin-2 reveals its inhibitory mechanism. Nat. Commun. 2022, 13, 6084. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Pathak, K.A.; Shrivastav, A.; Varma Shrivastav, S. Insulin-Like Growth Factor Binding Protein-3 Binds to Histone 3. Int. J. Mol. Sci. 2021, 22, 407. [Google Scholar] [CrossRef]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. [Google Scholar] [CrossRef]
- Zielinska, H.A.; Daly, C.S.; Alghamdi, A.; Bahl, A.; Sohail, M.; White, P.; Dean, S.R.; Holly, J.M.P.; Perks, C.M. Interaction between GRP78 and IGFBP-3 Affects Tumourigenesis and Prognosis in Breast Cancer Patients. Cancers 2020, 12, 3821. [Google Scholar] [CrossRef]
- Cai, W.; Sakaguchi, M.; Kleinridders, A.; Gonzalez-Del Pino, G.; Dreyfuss, J.M.; O’Neill, B.T.; Ramirez, A.K.; Pan, H.; Winnay, J.N.; Boucher, J.; et al. Domain-dependent effects of insulin and IGF-1 receptors on signalling and gene expression. Nat. Commun. 2017, 8, 14892. [Google Scholar] [CrossRef]
- Nagao, H.; Cai, W.; Wewer Albrechtsen, N.J.; Steger, M.; Batista, T.M.; Pan, H.; Dreyfuss, J.M.; Mann, M.; Kahn, C.R. Distinct signaling by insulin and IGF-1 receptors and their extra- and intracellular domains. Proc. Natl. Acad. Sci. USA 2021, 118, e2019474118. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Laron, Z.; Werner, H. Congenital IGF-1 deficiency protects from cancer: Lessons from Laron syndrome. Endocr. Relat. Cancer 2023, 30, e220394. [Google Scholar] [CrossRef]
- Boguszewski, C.L.; Boguszewski, M. Growth Hormone’s Links to Cancer. Endocr. Rev. 2019, 40, 558–574. [Google Scholar] [CrossRef]
- Knuppel, A.; Fensom, G.K.; Watts, E.L.; Gunter, M.J.; Murphy, N.; Papier, K.; Perez-Cornago, A.; Schmidt, J.A.; Smith Byrne, K.; Travis, R.C.; et al. Circulating Insulin-like Growth Factor-I Concentrations and Risk of 30 Cancers: Prospective Analyses in UK Biobank. Cancer Res. 2020, 80, 4014–4021. [Google Scholar] [CrossRef]
- Qiao, C.; Huang, W.; Chen, J.; Feng, W.; Zhang, T.; Wang, Y.; Liu, D.; Ji, X.; Xie, M.; Sun, M.; et al. IGF1-mediated HOXA13 overexpression promotes colorectal cancer metastasis through upregulating ACLY and IGF1R. Cell Death Dis. 2021, 12, 564. [Google Scholar] [CrossRef]
- Riis, S.; Murray, J.B.; O’Connor, R. IGF-1 Signalling Regulates Mitochondria Dynamics and Turnover through a Conserved GSK-3beta-Nrf2-BNIP3 Pathway. Cells 2020, 9, 147. [Google Scholar] [CrossRef]
- Lyons, A.; Coleman, M.; Riis, S.; Favre, C.; O’Flanagan, C.H.; Zhdanov, A.V.; Papkovsky, D.B.; Hursting, S.D.; O’Connor, R. Insulin-like growth factor 1 signaling is essential for mitochondrial biogenesis and mitophagy in cancer cells. J. Biol. Chem. 2017, 292, 16983–16998. [Google Scholar] [CrossRef]
- Artico, L.L.; Ruas, J.S.; Teixeira Junior, J.R.; Migita, N.A.; Seguchi, G.; Shi, X.; Brandalise, S.R.; Castilho, R.F.; Yunes, J.A. IGFBP7 Fuels the Glycolytic Metabolism in B-Cell Precursor Acute Lymphoblastic Leukemia by Sustaining Activation of the IGF1R-Akt-GLUT1 Axis. Int. J. Mol. Sci. 2023, 24, 9679. [Google Scholar] [CrossRef]
- Chen, W.J.; Tsai, J.H.; Hsu, L.S.; Lin, C.L.; Hong, H.M.; Pan, M.H. Quercetin blocks the aggressive phenotype of triple-negative breast cancer by inhibiting IGF1/IGF1R-mediated EMT program. J. Food Drug Anal. 2021, 29, 98–112. [Google Scholar] [CrossRef]
- Chan, Y.T.; Lin, R.J.; Wang, Y.H.; Hung, T.H.; Huang, Y.; Yu, J.; Yu, J.C.; Yu, A.L. The interplay between IGF-1R signaling and Hippo-YAP in breast cancer stem cells. Cell Commun. Signal. 2023, 21, 81. [Google Scholar] [CrossRef]
- Nagle, A.M.; Levine, K.M.; Tasdemir, N.; Scott, J.A.; Burlbaugh, K.; Kehm, J.; Katz, T.A.; Boone, D.N.; Jacobsen, B.M.; Atkinson, J.M.; et al. Loss of E-cadherin Enhances IGF1-IGF1R Pathway Activation and Sensitizes Breast Cancers to Anti-IGF1R/InsR Inhibitors. Clin. Cancer Res. 2018, 24, 5165–5177. [Google Scholar] [CrossRef]
- Elangovan, A.; Hooda, J.; Savariau, L.; Puthanmadhomnarayanan, S.; Yates, M.E.; Chen, J.; Brown, D.D.; McAuliffe, P.F.; Oesterreich, S.; Atkinson, J.M.; et al. Loss of E-cadherin Induces IGF1R Activation and Reveals a Targetable Pathway in Invasive Lobular Breast Carcinoma. Mol. Cancer Res. 2022, 20, 1405–1419. [Google Scholar] [CrossRef]
- Papoutsidakis, A.; Giatagana, E.M.; Berdiaki, A.; Spyridaki, I.; Spandidos, D.A.; Tsatsakis, A.; Tzanakakis, G.N.; Nikitovic, D. Lumican mediates HTB94 chondrosarcoma cell growth via an IGF-IR/Erk1/2 axis. Int. J. Oncol. 2020, 57, 791–803. [Google Scholar] [CrossRef]
- Stueven, N.A.; Beauvais, D.M.; Hu, R.; Kimple, R.J.; Rapraeger, A.C. Inhibiting IGF1R-mediated Survival Signaling in Head and Neck Cancer with the Peptidomimetic SSTN(IGF1R). Cancer Res. Commun. 2023, 3, 97–108. [Google Scholar] [CrossRef]
- Shi, Z.D.; Pang, K.; Wu, Z.X.; Dong, Y.; Hao, L.; Qin, J.X.; Wang, W.; Chen, Z.S.; Han, C.H. Tumor cell plasticity in targeted therapy-induced resistance: Mechanisms and new strategies. Signal Transduct. Target. Ther. 2023, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, B.; Li, H.; Wang, Y.; Liu, X.; Wong, K.Y.; Chan, W.N.; Chan, A.K.; Cheung, A.H.; Leung, K.T.; et al. Cancer-associated fibroblasts potentiate colorectal cancer progression by crosstalk of the IGF2-IGF1R and Hippo-YAP1 signaling pathways. J. Pathol. 2023, 259, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Aguilar-Mahecha, A.; Krzemien, U.; Hosein, A.; Buchanan, M.; Lafleur, J.; Pollak, M.; Ferrario, C.; Basik, M. Metastatic Breast Carcinoma-Associated Fibroblasts Have Enhanced Protumorigenic Properties Related to Increased IGF2 Expression. Clin. Cancer Res. 2019, 25, 7229–7242. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Li, Z.; Zhou, R.; Feng, W.; Yi, L.; Zhang, H.; Chen, B.; Li, Q.; Huang, S.; Zhu, X. Single cell and bulk RNA sequencing identifies tumor microenvironment subtypes and chemoresistance-related IGF1(+) cancer-associated fibroblast in gastric cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2024, 1870, 167123. [Google Scholar] [CrossRef] [PubMed]
- Tommelein, J.; De Vlieghere, E.; Verset, L.; Melsens, E.; Leenders, J.; Descamps, B.; Debucquoy, A.; Vanhove, C.; Pauwels, P.; Gespach, C.P.; et al. Radiotherapy-Activated Cancer-Associated Fibroblasts Promote Tumor Progression through Paracrine IGF1R Activation. Cancer Res. 2018, 78, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Remsing Rix, L.L.; Sumi, N.J.; Hu, Q.; Desai, B.; Bryant, A.T.; Li, X.; Welsh, E.A.; Fang, B.; Kinose, F.; Kuenzi, B.M.; et al. IGF-binding proteins secreted by cancer-associated fibroblasts induce context-dependent drug sensitization of lung cancer cells. Sci. Signal. 2022, 15, eabj5879. [Google Scholar] [CrossRef] [PubMed]
- Cambria, D.; Longhitano, L.; La Spina, E.; Giallongo, S.; Orlando, L.; Giuffrida, R.; Tibullo, D.; Fontana, P.; Barbagallo, I.; Nicoletti, V.G.; et al. IGFBP-6 Alters Mesenchymal Stromal Cell Phenotype Driving Dasatinib Resistance in Chronic Myeloid Leukemia. Life 2023, 13, 259. [Google Scholar] [CrossRef]
- Lv, J.; Liu, C.; Chen, F.K.; Feng, Z.P.; Jia, L.; Liu, P.J.; Yang, Z.X.; Hou, F.; Deng, Z.Y. M2-like tumour-associated macrophage-secreted IGF promotes thyroid cancer stemness and metastasis by activating the PI3K/AKT/mTOR pathway. Mol. Med. Rep. 2021, 24, 604. [Google Scholar] [CrossRef] [PubMed]
- Ireland, L.; Santos, A.; Campbell, F.; Figueiredo, C.; Hammond, D.; Ellies, L.G.; Weyer-Czernilofsky, U.; Bogenrieder, T.; Schmid, M.; Mielgo, A. Blockade of insulin-like growth factors increases efficacy of paclitaxel in metastatic breast cancer. Oncogene 2018, 37, 2022–2036. [Google Scholar] [CrossRef]
- Ma, J.; Chen, Z.; Li, Q.; Wang, L.; Chen, J.; Yang, X.; Yang, C.; Quan, Z. RARRES2 is involved in the “lock-and-key” interactions between osteosarcoma stem cells and tumor-associated macrophages. Sci. Rep. 2024, 14, 2267. [Google Scholar] [CrossRef]
- Du, L.; Lin, L.; Li, Q.; Liu, K.; Huang, Y.; Wang, X.; Cao, K.; Chen, X.; Cao, W.; Li, F.; et al. IGF-2 Preprograms Maturing Macrophages to Acquire Oxidative Phosphorylation-Dependent Anti-inflammatory Properties. Cell Metab. 2019, 29, 1363–1375.e8. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, L.; Cao, J.; Ge, J.; Liu, X.; Liu, P.; Teng, Y.; Wang, S.; Sun, Y.; Liu, M.; et al. DACH1 regulates macrophage activation and tumour progression in hypopharyngeal squamous cell carcinoma. Immunology 2023, 170, 253–269. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, S.Q.; Hang, L.; Zhang, C.F.; Wang, L.; Duan, C.J.; Cheng, Y.D.; Wu, D.K.; Chen, R. GRP78 facilitates M2 macrophage polarization and tumour progression. Cell. Mol. Life Sci. 2021, 78, 7709–7732. [Google Scholar] [CrossRef] [PubMed]
- Alturki, N.A. Review of the Immune Checkpoint Inhibitors in the Context of Cancer Treatment. J. Clin. Med. 2023, 12, 4301. [Google Scholar] [CrossRef]
- Ajona, D.; Ortiz-Espinosa, S.; Lozano, T.; Exposito, F.; Calvo, A.; Valencia, K.; Redrado, M.; Remirez, A.; Lecanda, F.; Alignani, D.; et al. Short-term starvation reduces IGF-1 levels to sensitize lung tumors to PD-1 immune checkpoint blockade. Nat. Cancer 2020, 1, 75–85. [Google Scholar] [CrossRef]
- Durfort, T.; Tkach, M.; Meschaninova, M.I.; Rivas, M.A.; Elizalde, P.V.; Venyaminova, A.G.; Schillaci, R.; Francois, J.C. Small interfering RNA targeted to IGF-IR delays tumor growth and induces proinflammatory cytokines in a mouse breast cancer model. PLoS ONE 2012, 7, e29213. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Tian, A.L.; Li, B.; Leduc, M.; Forveille, S.; Hamley, P.; Galloway, W.; Xie, W.; Liu, P.; Zhao, L.; et al. IGF1 receptor inhibition amplifies the effects of cancer drugs by autophagy and immune-dependent mechanisms. J. Immunother. Cancer 2021, 9, e002722. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, S.; Lee, C.; Brandhorst, S.; Manes, B.; Buono, R.; Cheng, C.W.; Cacciottolo, M.; Martin-Montalvo, A.; de Cabo, R.; Wei, M.; et al. Fasting-Mimicking Diet Reduces HO-1 to Promote T Cell-Mediated Tumor Cytotoxicity. Cancer Cell 2016, 30, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Pol, J.; Vacchelli, E.; Rao, S.; Enot, D.P.; Baracco, E.E.; Levesque, S.; Castoldi, F.; Jacquelot, N.; Yamazaki, T.; et al. Caloric Restriction Mimetics Enhance Anticancer Immunosurveillance. Cancer Cell 2016, 30, 147–160. [Google Scholar] [CrossRef]
- Heckl, S.M.; Mau, F.; Senftleben, A.; Daunke, T.; Beckinger, S.; Abdullazade, S.; Schreiber, S.; Rocken, C.; Sebens, S.; Schafer, H. Programmed Death-Ligand 1 (PD-L1) Expression Is Induced by Insulin in Pancreatic Ductal Adenocarcinoma Cells Pointing to Its Role in Immune Checkpoint Control. Med. Sci. 2021, 9, 48. [Google Scholar] [CrossRef]
- Peng, M.; Fan, S.; Li, J.; Zhou, X.; Liao, Q.; Tang, F.; Liu, W. Programmed death-ligand 1 signaling and expression are reversible by lycopene via PI3K/AKT and Raf/MEK/ERK pathways in tongue squamous cell carcinoma. Genes Nutr. 2022, 17, 3. [Google Scholar] [CrossRef]
- Yue, M.; Hu, S.; Sun, H.; Tuo, B.; Jia, B.; Chen, C.; Wang, W.; Liu, J.; Liu, Y.; Sun, Z.; et al. Extracellular vesicles remodel tumor environment for cancer immunotherapy. Mol. Cancer 2023, 22, 203. [Google Scholar] [CrossRef]
- Bhatta, R.; Han, J.; Liu, Y.; Bo, Y.; Lee, D.; Zhou, J.; Wang, Y.; Nelson, E.R.; Chen, Q.; Zhang, X.S.; et al. Metabolic tagging of extracellular vesicles and development of enhanced extracellular vesicle based cancer vaccines. Nat. Commun. 2023, 14, 8047. [Google Scholar] [CrossRef]
- DeRita, R.M.; Zerlanko, B.; Singh, A.; Lu, H.; Iozzo, R.V.; Benovic, J.L.; Languino, L.R. c-Src, Insulin-Like Growth Factor I Receptor, G-Protein-Coupled Receptor Kinases and Focal Adhesion Kinase are Enriched Into Prostate Cancer Cell Exosomes. J. Cell. Biochem. 2017, 118, 66–73. [Google Scholar] [CrossRef]
- He, M.; Crow, J.; Roth, M.; Zeng, Y.; Godwin, A.K. Integrated immunoisolation and protein analysis of circulating exosomes using microfluidic technology. Lab Chip 2014, 14, 3773–3780. [Google Scholar] [CrossRef]
- Saftics, A.; Abuelreich, S.; Romano, E.; Ghaeli, I.; Jiang, N.; Spanos, M.; Lennon, K.M.; Singh, G.; Das, S.; Van Keuren-Jensen, K.; et al. Single Extracellular VEsicle Nanoscopy. J. Extracell. Vesicles 2023, 12, e12346. [Google Scholar] [CrossRef]
- Stiller, C.; Viktorsson, K.; Paz Gomero, E.; Haag, P.; Arapi, V.; Kaminskyy, V.O.; Kamali, C.; De Petris, L.; Ekman, S.; Lewensohn, R.; et al. Detection of Tumor-Associated Membrane Receptors on Extracellular Vesicles from Non-Small Cell Lung Cancer Patients via Immuno-PCR. Cancers 2021, 13, 922. [Google Scholar] [CrossRef]
- Wang, X.; Chai, Z.; Pan, G.; Hao, Y.; Li, B.; Ye, T.; Li, Y.; Long, F.; Xia, L.; Liu, M. ExoBCD: A comprehensive database for exosomal biomarker discovery in breast cancer. Brief. Bioinform. 2021, 22, bbaa088. [Google Scholar] [CrossRef]
- Chen, C.; Yang, C.; Tian, X.; Liang, Y.; Wang, S.; Wang, X.; Shou, Y.; Li, H.; Xiao, Q.; Shu, J.; et al. Downregulation of miR-100-5p in cancer-associated fibroblast-derived exosomes facilitates lymphangiogenesis in esophageal squamous cell carcinoma. Cancer Med. 2023, 12, 14468–14483. [Google Scholar] [CrossRef]
- Hsu, X.R.; Wu, J.E.; Wu, Y.Y.; Hsiao, S.Y.; Liang, J.L.; Wu, Y.J.; Tung, C.H.; Huang, M.F.; Lin, M.S.; Yang, P.C.; et al. Exosomal long noncoding RNA MLETA1 promotes tumor progression and metastasis by regulating the miR-186-5p/EGFR and miR-497-5p/IGF1R axes in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2023, 42, 283. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, H.; He, K.; Wang, J. Human bone marrow mesenchymal stem cells-derived exosomes attenuated prostate cancer progression via the miR-99b-5p/IGF1R axis. Bioengineered 2022, 13, 2004–2016. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, V.; Xu, B.; Akers, J.; Nguyen, T.; Ma, J.; Dhawan, S.; Ning, J.; Mao, Y.; Hua, W.; Kokkoli, E.; et al. Radiation-induced extracellular vesicle (EV) release of miR-603 promotes IGF1-mediated stem cell state in glioblastomas. EBioMedicine 2020, 55, 102736. [Google Scholar] [CrossRef] [PubMed]
- Mancarella, C.; Giusti, V.; Caldoni, G.; Laginestra, M.A.; Parra, A.; Toracchio, L.; Giordano, G.; Roncuzzi, L.; Piazzi, M.; Blalock, W.; et al. Extracellular vesicle-associated IGF2BP3 tunes Ewing sarcoma cell migration and affects PI3K/Akt pathway in neighboring cells. Cancer Gene Ther. 2023, 30, 1285–1295. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Sang, H.; Zhou, Y.; Shang, C.; Wang, Y.; Zhu, H. Effects of hyperglycemia on the progression of tumor diseases. J. Exp. Clin. Cancer Res. 2019, 38, 327. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.; Kaminsky-Forrett, M.C.; Henry, S.; Zanetta, S.; Geoffrois, L.; Bompas, E.; Moxhon, A.; Mignion, L.; Guigay, J.; Knoops, L.; et al. Phase II study of figitumumab in patients with recurrent and/or metastatic squamous cell carcinoma of the head and neck: Clinical activity and molecular response (GORTEC 2008-02). Ann. Oncol. 2012, 23, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Palanissami, G.; Paul, S.F.D. AGEs and RAGE: Metabolic and molecular signatures of the glycation-inflammation axis in malignant or metastatic cancers. Explor. Target. Antitumor Ther. 2023, 4, 812–849. [Google Scholar] [CrossRef] [PubMed]
- Muoio, M.G.; Talia, M.; Lappano, R.; Sims, A.H.; Vella, V.; Cirillo, F.; Manzella, L.; Giuliano, M.; Maggiolini, M.; Belfiore, A.; et al. Activation of the S100A7/RAGE Pathway by IGF-1 Contributes to Angiogenesis in Breast Cancer. Cancers 2021, 13, 621. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, M.; Aston, C.; Hintz, R.; Vlassara, H. Receptor-specific induction of insulin-like growth factor I in human monocytes by advanced glycosylation end product-modified proteins. J. Clin. Investig. 1992, 90, 439–446. [Google Scholar] [CrossRef]
- Yang, S.J.; Chen, C.Y.; Chang, G.D.; Wen, H.C.; Chen, C.Y.; Chang, S.C.; Liao, J.F.; Chang, C.H. Activation of Akt by advanced glycation end products (AGEs): Involvement of IGF-1 receptor and caveolin-1. PLoS ONE 2013, 8, e58100. [Google Scholar] [CrossRef]
- Monden, M.; Koyama, H.; Otsuka, Y.; Morioka, T.; Mori, K.; Shoji, T.; Mima, Y.; Motoyama, K.; Fukumoto, S.; Shioi, A.; et al. Receptor for advanced glycation end products regulates adipocyte hypertrophy and insulin sensitivity in mice: Involvement of Toll-like receptor 2. Diabetes 2013, 62, 478–489. [Google Scholar] [CrossRef]
- Chen, Z.; Hong, Q. Correlation of serum IGF-1, AGEs and their receptors with the risk of colorectal cancer in patients with type 2 diabetes mellitus. Front. Oncol. 2023, 13, 1125745. [Google Scholar] [CrossRef] [PubMed]
- Poljicanin, A.; Filipovic, N.; Vukusic Pusic, T.; Soljic, V.; Caric, A.; Saraga-Babic, M.; Vukojevic, K. Expression pattern of RAGE and IGF-1 in the human fetal ovary and ovarian serous carcinoma. Acta Histochem. 2015, 117, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Mak, V.C.; Cheung, L.W. Drugging IGF-1R in cancer: New insights and emerging opportunities. Genes Dis. 2023, 10, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Davidson, T.M.; Lebreton, C.L.; Hendricksen, A.E.W.; Atkinson, H.J.; Larson, M.C.; Oberg, A.L.; Provencher, D.M.; Glaspy, J.A.; Karlan, B.Y.; Slamon, D.J.; et al. Results of TRIO-15, a multicenter, open-label, phase II study of the efficacy and safety of ganitumab in patients with recurrent platinum-sensitive ovarian cancer. Gynecol. Oncol. 2023, 170, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.M.; Bielack, S.S.; Gorlick, R.G.; Skubitz, K.; Daw, N.C.; Herzog, C.E.; Monge, O.R.; Lassaletta, A.; Boldrini, E.; Papai, Z.; et al. A phase II study of clinical activity of SCH 717454 (robatumumab) in patients with relapsed osteosarcoma and Ewing sarcoma. Pediatr. Blood Cancer 2016, 63, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Akshintala, S.; Sundby, R.T.; Bernstein, D.; Glod, J.W.; Kaplan, R.N.; Yohe, M.E.; Gross, A.M.; Derdak, J.; Lei, H.; Pan, A.; et al. Phase I trial of Ganitumab plus Dasatinib to Cotarget the Insulin-Like Growth Factor 1 Receptor and Src Family Kinase YES in Rhabdomyosarcoma. Clin. Cancer Res. 2023, 29, 3329–3339. [Google Scholar] [CrossRef] [PubMed]
- Shulman, D.S.; Merriam, P.; Choy, E.; Guenther, L.M.; Cavanaugh, K.L.; Kao, P.C.; Posner, A.; Bhushan, K.; Fairchild, G.; Barker, E.; et al. Phase 2 trial of palbociclib and ganitumab in patients with relapsed Ewing sarcoma. Cancer Med. 2023, 12, 15207–15216. [Google Scholar] [CrossRef] [PubMed]
- Scagliotti, G.V.; Bondarenko, I.; Blackhall, F.; Barlesi, F.; Hsia, T.C.; Jassem, J.; Milanowski, J.; Popat, S.; Sanchez-Torres, J.M.; Novello, S.; et al. Randomized, phase III trial of figitumumab in combination with erlotinib versus erlotinib alone in patients with nonadenocarcinoma nonsmall-cell lung cancer. Ann. Oncol. 2015, 26, 497–504. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.J.; Chudow, J.; DeNunzio, M.; Slovin, S.F.; Danila, D.C.; Morris, M.J.; Scher, H.I.; Rathkopf, D.E. A Phase I Trial of IGF-1R Inhibitor Cixutumumab and mTOR Inhibitor Temsirolimus in Metastatic Castration-resistant Prostate Cancer. Clin. Genitourin. Cancer 2020, 18, 171–178.e2. [Google Scholar] [CrossRef]
- Haddad, T.C.; He, J.; O’Sullivan, C.C.; Chen, B.; Northfelt, D.; Dueck, A.C.; Ballman, K.V.; Tenner, K.S.; Linden, H.; Sparano, J.A.; et al. Randomized Phase II Trial of Capecitabine and Lapatinib with or without IMC-A12 (Cituxumumab) in Patients with HER2-Positive Advanced Breast Cancer Previously Treated with Trastuzumab and Chemotherapy: NCCTG N0733 (Alliance). Breast Cancer Res. Treat. 2021, 188, 477–487. [Google Scholar] [CrossRef]
- Khan, S.; LeBlanc, R.; Gyger, M.; White, D.; Kaufman, J.; Jazubowiak, A.; Gul, E.; Paul, H.; Le, L.W.; Lau, A.; et al. A phase-1 trial of linsitinib (OSI-906) in combination with bortezomib and dexamethasone for the treatment of relapsed/refractory multiple myeloma. Leuk. Lymphoma 2021, 62, 1721–1729. [Google Scholar] [CrossRef] [PubMed]
- von Mehren, M.; George, S.; Heinrich, M.C.; Schuetze, S.M.; Yap, J.T.; Yu, J.Q.; Abbott, A.; Litwin, S.; Crowley, J.; Belinsky, M.; et al. Linsitinib (OSI-906) for the Treatment of Adult and Pediatric Wild-Type Gastrointestinal Stromal Tumors, a SARC Phase II Study. Clin. Cancer Res. 2020, 26, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Bergqvist, M.; Holgersson, G.; Bondarenko, I.; Grechanaya, E.; Maximovich, A.; Andor, G.; Klockare, M.; Thureson, M.; Jerling, M.; Harmenberg, J. Phase II randomized study of the IGF-1R pathway modulator AXL1717 compared to docetaxel in patients with previously treated, locally advanced or metastatic non-small cell lung cancer. Acta Oncol. 2017, 56, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Haluska, P.; Menefee, M.; Plimack, E.R.; Rosenberg, J.; Northfelt, D.; LaVallee, T.; Shi, L.; Yu, X.Q.; Burke, P.; Huang, J.; et al. Phase I dose-escalation study of MEDI-573, a bispecific, antiligand monoclonal antibody against IGFI and IGFII, in patients with advanced solid tumors. Clin. Cancer Res. 2014, 20, 4747–4757. [Google Scholar] [CrossRef]
- Iguchi, H.; Nishina, T.; Nogami, N.; Kozuki, T.; Yamagiwa, Y.; Yagawa, K. Phase I dose-escalation study evaluating safety, tolerability and pharmacokinetics of MEDI-573, a dual IGF-I/II neutralizing antibody, in Japanese patients with advanced solid tumours. Investig. New Drugs 2015, 33, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Macaulay, V.M.; Lord, S.; Hussain, S.; Maroto, J.P.; Jones, R.H.; Climent, M.A.; Cook, N.; Lin, C.C.; Wang, S.S.; Bianchini, D.; et al. A Phase Ib/II study of IGF-neutralising antibody xentuzumab with enzalutamide in metastatic castration-resistant prostate cancer. Br. J. Cancer 2023, 129, 965–973. [Google Scholar] [CrossRef]
- de Bono, J.; Lin, C.C.; Chen, L.T.; Corral, J.; Michalarea, V.; Rihawi, K.; Ong, M.; Lee, J.H.; Hsu, C.H.; Yang, J.C.; et al. Two first-in-human studies of xentuzumab, a humanised insulin-like growth factor (IGF)-neutralising antibody, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1324–1332. [Google Scholar] [CrossRef]
- Schmid, P.; Cortes, J.; Joaquim, A.; Janez, N.M.; Morales, S.; Diaz-Redondo, T.; Blau, S.; Neven, P.; Lemieux, J.; Garcia-Saenz, J.A.; et al. XENERA-1: A randomised double-blind Phase II trial of xentuzumab in combination with everolimus and exemestane versus everolimus and exemestane in patients with hormone receptor-positive/HER2-negative metastatic breast cancer and non-visceral disease. Breast Cancer Res. 2023, 25, 67. [Google Scholar] [CrossRef]
- Chen, Y.M.; Leibovitch, M.; Zeinieh, M.; Jabado, N.; Brodt, P. Targeting the IGF-Axis in Cultured Pediatric High-Grade Glioma Cells Inhibits Cell Cycle Progression and Survival. Pharmaceuticals 2023, 16, 297. [Google Scholar] [CrossRef]
- Wang, N.; Rayes, R.F.; Elahi, S.M.; Lu, Y.; Hancock, M.A.; Massie, B.; Rowe, G.E.; Aomari, H.; Hossain, S.; Durocher, Y.; et al. The IGF-Trap: Novel Inhibitor of Carcinoma Growth and Metastasis. Mol. Cancer Ther. 2015, 14, 982–993. [Google Scholar] [CrossRef]
- Vaniotis, G.; Moffett, S.; Sulea, T.; Wang, N.; Elahi, S.M.; Lessard, E.; Baardsnes, J.; Perrino, S.; Durocher, Y.; Frystyk, J.; et al. Enhanced anti-metastatic bioactivity of an IGF-TRAP re-engineered to improve physicochemical properties. Sci. Rep. 2018, 8, 17361. [Google Scholar] [CrossRef] [PubMed]
- Tsui, J.; Qi, S.; Perrino, S.; Leibovitch, M.; Brodt, P. Identification of a Resistance Mechanism to IGF-IR Targeting in Human Triple Negative MDA-MB-231 Breast Cancer Cells. Biomolecules 2021, 11, 527. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Konda, J.D.; Perrino, S.; Celia Fernandez, M.; Lowy, A.M.; Brodt, P. Targeting the IGF-Axis Potentiates Immunotherapy for Pancreatic Ductal Adenocarcinoma Liver Metastases by Altering the Immunosuppressive Microenvironment. Mol. Cancer Ther. 2021, 20, 2469–2482. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Mancarella, C.; Grilli, A.; Manara, M.C.; Astolfi, A.; Marino, M.T.; Conte, A.; Sigismund, S.; Care, A.; Belfiore, A.; et al. Identification of common and distinctive mechanisms of resistance to different anti-IGF-IR agents in Ewing’s sarcoma. Mol. Endocrinol. 2012, 26, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, D.; Siroky, M.D.; Miller, S.M. Next-generation biological vector platforms for in vivo delivery of genome editing agents. Curr. Opin. Biotechnol. 2023, 85, 103040. [Google Scholar] [CrossRef]
- Samani, A.A.; Nalbantoglu, J.; Brodt, P. Glioma Cells With Genetically Engineered IGF-I Receptor Downregulation Can Persist in the Brain in a Dormant State. Front. Oncol. 2020, 10, 555945. [Google Scholar] [CrossRef]
- Samani, A.A.; Fallavollita, L.; Jaalouk, D.E.; Galipeau, J.; Brodt, P. Inhibition of carcinoma cell growth and metastasis by a vesicular stomatitis virus G-pseudotyped retrovector expressing type I insulin-like growth factor receptor antisense. Hum. Gene Ther. 2001, 12, 1969–1977. [Google Scholar] [CrossRef] [PubMed]
- Trojan, A.; Chun Lone, Y.; Briceno, I.; Trojan, J. Anti-Gene IGF-I Vaccines in Cancer Gene Therapy: A Review of a Case of Glioblastoma. Curr. Med. Chem. 2023, 31, 1983–2002. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Ma, J.; Zeng, B.J.; Catanese, V.M.; Samuels, S.; Gama Sosa, M.A.; Kolodny, E.H. Correlation of glioma cell regression with inhibition of insulin-like growth factor 1 and insulin-like growth factor-binding protein-2 expression. Neuroendocrinology 1997, 66, 203–211. [Google Scholar] [CrossRef]
- Dale, B.; Cheng, M.; Park, K.S.; Kaniskan, H.U.; Xiong, Y.; Jin, J. Advancing targeted protein degradation for cancer therapy. Nat. Rev. Cancer 2021, 21, 638–654. [Google Scholar] [CrossRef]
- Mancarella, C.; Morrione, A.; Scotlandi, K. PROTAC-Based Protein Degradation as a Promising Strategy for Targeted Therapy in Sarcomas. Int. J. Mol. Sci. 2023, 24, 16346. [Google Scholar] [CrossRef]
- Wan, X.; Yeung, C.; Heske, C.; Mendoza, A.; Helman, L.J. IGF-1R Inhibition Activates a YES/SFK Bypass Resistance Pathway: Rational Basis for Co-Targeting IGF-1R and Yes/SFK Kinase in Rhabdomyosarcoma. Neoplasia 2015, 17, 358–366. [Google Scholar] [CrossRef]
- Manda, S.; Lee, N.K.; Oh, D.C.; Lee, J. Design, Synthesis, and Biological Evaluation of Proteolysis Targeting Chimeras (PROTACs) for the Dual Degradation of IGF-1R and Src. Molecules 2020, 25, 1948. [Google Scholar] [CrossRef]
- Yu, X.; Xu, J.; Cahuzac, K.M.; Xie, L.; Shen, Y.; Chen, X.; Liu, J.; Parsons, R.E.; Jin, J. Novel Allosteric Inhibitor-Derived AKT Proteolysis Targeting Chimeras (PROTACs) Enable Potent and Selective AKT Degradation in KRAS/BRAF Mutant Cells. J. Med. Chem. 2022, 65, 14237–14260. [Google Scholar] [CrossRef]
- Yu, X.; Xu, J.; Shen, Y.; Cahuzac, K.M.; Park, K.S.; Dale, B.; Liu, J.; Parsons, R.E.; Jin, J. Discovery of Potent, Selective, and In Vivo Efficacious AKT Kinase Protein Degraders via Structure-Activity Relationship Studies. J. Med. Chem. 2022, 65, 3644–3666. [Google Scholar] [CrossRef]
- Yu, X.; Xu, J.; Xie, L.; Wang, L.; Shen, Y.; Cahuzac, K.M.; Chen, X.; Liu, J.; Parsons, R.E.; Jin, J. Design, Synthesis, and Evaluation of Potent, Selective, and Bioavailable AKT Kinase Degraders. J. Med. Chem. 2021, 64, 18054–18081. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.L.; Luo, X.; Tian, T.; Rao, Z.; Wang, H.; Zhou, Z.; Mi, T.; Chen, D.; Xu, Y.; Wu, Y.; et al. Structure-based rational design enables efficient discovery of a new selective and potent AKT PROTAC degrader. Eur. J. Med. Chem. 2022, 238, 114459. [Google Scholar] [CrossRef]
- Jauslin, W.T.; Schild, M.; Schaefer, T.; Borsari, C.; Orbegozo, C.; Bissegger, L.; Zhanybekova, S.; Ritz, D.; Schmidt, A.; Wymann, M.; et al. A high affinity pan-PI3K binding module supports selective targeted protein degradation of PI3Kalpha. Chem. Sci. 2024, 15, 683–691. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Therapy | Drug | Phase | Disease | Outcomes | References |
|---|---|---|---|---|---|
| Monoclonal antibodies | Robatumumab | II | Relapsed Ewing sarcoma and osteosarcoma | Osteosarcoma: 3/80 CR or PR; 23/80 SD Ewing sarcoma: 6/84 CR or PR; 23/84 SD | [115] |
| Ganitumab (+ dasatinib) | I | Rhabdomyosarcoma | 1/9 PR, 1/9 SD | [116] | |
| Ganitumab (+ palbociclib) | II | Relapsed Ewing sarcoma | 2/10 SD | [117] | |
| Figitumumab (+ erlotinib) | III | Non adenocarcinoma non-small-cell lung cancer | 16/293 PR, 113/293 SD | [118] | |
| Figitumumab | II | Squamous cell carcinoma of the head and neck | 2/17 SD | [105] | |
| Cixutumumab (+ Temsirolimus) | I | Castration-resistant prostate cancer | 3/16 SD | [119] | |
| Cixutumumab (+ capecitabine, lapatinib) | II | HER2-positive advanced breast cancer | No objective response | [120] | |
| Tyrosine kinase inhibitors | Linsitinib (+ bortezomib and dexamethasone) | I | Relapsed/refractory multiple myeloma | No clinical benefit | [121] |
| Linsitinib | II | Gastrointestinal stromal tumors | No objective responses | [122] | |
| AXL-1717 | II | Non-small-cell lung cancer | 24% CR + PR + SD | [123] | |
| Neutralizing antibodies | Dusigitumab | I | Advanced solid tumors | 13/39 SD | [124] |
| Dusigitumab | I | Advanced solid tumors | 4/10 SD | [125] | |
| Xentuzumab (+ enzalutamide) | Ib/II | Castration-resistant prostate cancer | No antitumor activity | [126] | |
| Xentuzumab | I | Advanced solid tumors | 2/61 PR, 3/61 SD | [127] | |
| Xentuzumab (+ everolimus) | II | Breast cancer with non-visceral disease | No clinical benefit | [128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancarella, C.; Morrione, A.; Scotlandi, K. Extracellular Interactors of the IGF System: Impact on Cancer Hallmarks and Therapeutic Approaches. Int. J. Mol. Sci. 2024, 25, 5915. https://doi.org/10.3390/ijms25115915
Mancarella C, Morrione A, Scotlandi K. Extracellular Interactors of the IGF System: Impact on Cancer Hallmarks and Therapeutic Approaches. International Journal of Molecular Sciences. 2024; 25(11):5915. https://doi.org/10.3390/ijms25115915
Chicago/Turabian StyleMancarella, Caterina, Andrea Morrione, and Katia Scotlandi. 2024. "Extracellular Interactors of the IGF System: Impact on Cancer Hallmarks and Therapeutic Approaches" International Journal of Molecular Sciences 25, no. 11: 5915. https://doi.org/10.3390/ijms25115915
APA StyleMancarella, C., Morrione, A., & Scotlandi, K. (2024). Extracellular Interactors of the IGF System: Impact on Cancer Hallmarks and Therapeutic Approaches. International Journal of Molecular Sciences, 25(11), 5915. https://doi.org/10.3390/ijms25115915