
Imbalance in Unc80 RNA Editing Disrupts Dynamic Neuronal Activity and Olfactory Perception
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Characterization of the Expression and RNA Editing Event of Unc80
2.2. Engineering and Phenotyping of the Unc80 Knockout and Knock-In Mouse Models
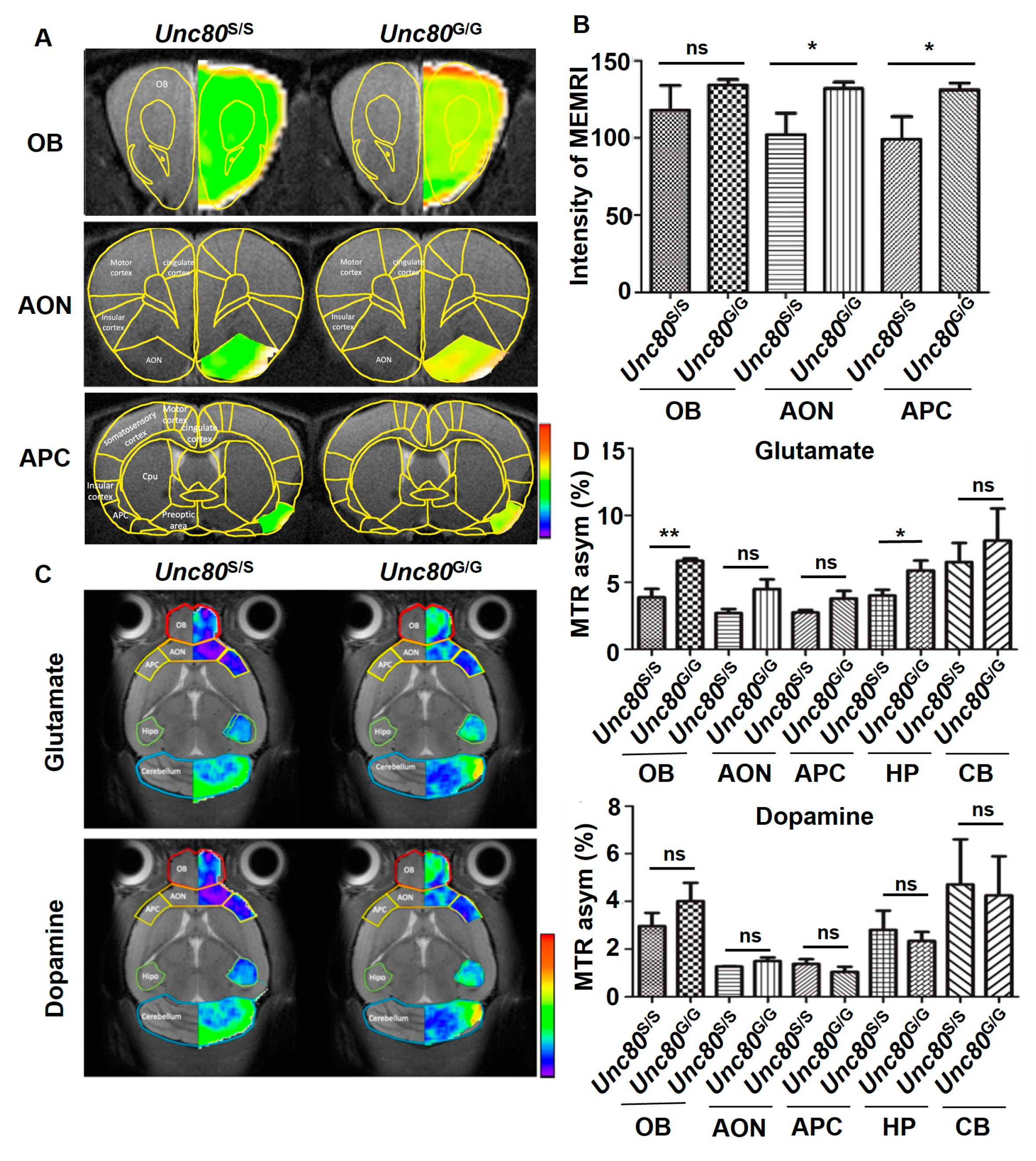
2.3. Neuronal Activity in the Unc80 Animal Models Revealed by MRI
2.4. Implication of Unc80 Editing Event in Mediating Olfactory Response
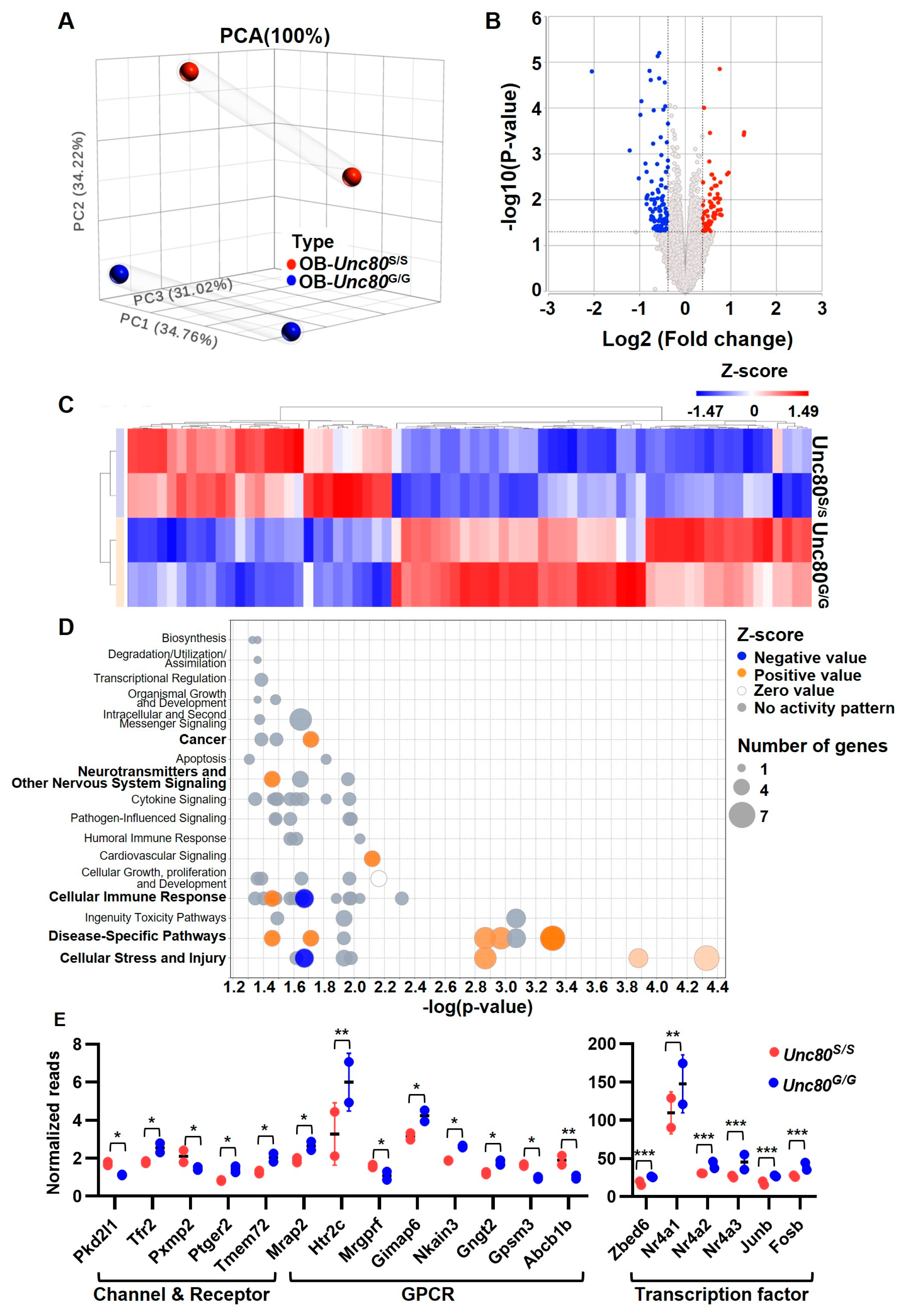
2.5. Transcriptome Profiling of Unc80 Knock-In Mice Revealed Altered Neuronal State in Olfactory Bulb
3. Discussion
4. Materials and Methods
4.1. CRISPR-Mediated Genome Editing for Generation of Point Mutant Mice
4.2. Animals
4.3. RNA Extraction, RT-PCR, and qPCR
4.4. Immunofluorescent Staining of Brain Tissue
4.5. Manganese-Enhanced Magnetic Resonance Imaging (MEMRI)
4.6. Chemical Exchange Saturation Transfer (CEST)-MRI
4.7. The Olfactory Habituation/Dishabituation Test
4.8. RNA-Sequencing
4.9. In Silico Prediction of Unc80WT and Unc80S2367G Protein Structures
4.10. Neuro 2a Cell Culture and Transfection
4.11. Indirect Immunofluorescence and Confocal Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, I.X.; So, E.; Devlin, J.L.; Zhao, Y.; Wu, M.; Cheung, V.G. ADAR regulates RNA editing, transcript stability, and gene expression. Cell Rep. 2013, 5, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Behm, M.; Wahlstedt, H.; Widmark, A.; Eriksson, M.; Ohman, M. Accumulation of nuclear ADAR2 regulates adenosine-to-inosine RNA editing during neuronal development. J. Cell Sci. 2017, 130, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Wahlstedt, H.; Daniel, C.; Enstero, M.; Ohman, M. Large-scale mRNA sequencing determines global regulation of RNA editing during brain development. Genome Res. 2009, 19, 978–986. [Google Scholar] [CrossRef]
- Konen, L.M.; Wright, A.L.; Royle, G.A.; Morris, G.P.; Lau, B.K.; Seow, P.W.; Zinn, R.; Milham, L.T.; Vaughan, C.W.; Vissel, B. A new mouse line with reduced GluA2 Q/R site RNA editing exhibits loss of dendritic spines, hippocampal CA1-neuron loss, learning and memory impairments and NMDA receptor-independent seizure vulnerability. Mol. Brain 2020, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Hideyama, T.; Yamashita, T.; Aizawa, H.; Tsuji, S.; Kakita, A.; Takahashi, H.; Kwak, S. Profound downregulation of the RNA editing enzyme ADAR2 in ALS spinal motor neurons. Neurobiol. Dis. 2012, 45, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Kwak, S. The molecular link between inefficient GluA2 Q/R site-RNA editing and TDP-43 pathology in motor neurons of sporadic amyotrophic lateral sclerosis patients. Brain Res. 2014, 1584, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Maas, S.; Single, F.N.; Hartner, J.; Rozov, A.; Burnashev, N.; Feldmeyer, D.; Sprengel, R.; Seeburg, P.H. Point mutation in an AMPA receptor gene rescues lethality in mice deficient in the RNA-editing enzyme ADAR2. Nature 2000, 406, 78–81. [Google Scholar] [CrossRef]
- Horsch, M.; Seeburg, P.H.; Adler, T.; Aguilar-Pimentel, J.A.; Becker, L.; Calzada-Wack, J.; Garrett, L.; Gotz, A.; Hans, W.; Higuchi, M.; et al. Requirement of the RNA-editing enzyme ADAR2 for normal physiology in mice. J. Biol. Chem. 2011, 286, 18614–18622. [Google Scholar] [CrossRef]
- Paschen, W.; Dux, E.; Djuricic, B. Developmental changes in the extent of RNA editing of glutamate receptor subunit GluR5 in rat brain. Neurosci. Lett. 1994, 174, 109–112. [Google Scholar] [CrossRef]
- Sommer, B.; Köhler, M.; Sprengel, R.; Seeburg, P.H. RNA editing in brain controls a determinant of ion flow in glutamate-gated channels. Cell 1991, 67, 11–19. [Google Scholar] [CrossRef]
- Niswender, C.M.; Copeland, S.C.; Herrick-Davis, K.; Emeson, R.B.; Sanders-Bush, E. RNA editing of the human serotonin 5-hydroxytryptamine 2C receptor silences constitutive activity. J. Biol. Chem. 1999, 274, 9472–9478. [Google Scholar] [CrossRef] [PubMed]
- Shumate, K.M.; Tas, S.T.; Kavalali, E.T.; Emeson, R.B. RNA editing-mediated regulation of calcium-dependent activator protein for secretion (CAPS1) localization and its impact on synaptic transmission. J. Neurochem. 2021, 158, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Khermesh, K.; D’Erchia, A.M.; Barak, M.; Annese, A.; Wachtel, C.; Levanon, E.Y.; Picardi, E.; Eisenberg, E. Reduced levels of protein recoding by A-to-I RNA editing in Alzheimer’s disease. RNA 2016, 22, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Pozdyshev, D.V.; Zharikova, A.A.; Medvedeva, M.V.; Muronetz, V.I. Differential analysis of A-to-I mRNA edited sites in Parkinson’s disease. Genes 2021, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, N.; Pandey, V.; Bhattacharyya, M.; Dey, A. Regulatory role of long non coding RNAs (lncRNAs) in neurological disorders: From novel biomarkers to promising therapeutic strategies. Asian J. Pharm. Sci. 2021, 16, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.S.; Dobbyn, A.; Li, Q.; Roussos, P.; Hoffman, G.E.; Stahl, E.; Chess, A.; Sklar, P.; Li, J.B.; Devlin, B. Global landscape and genetic regulation of RNA editing in cortical samples from individuals with schizophrenia. Nat. Neurosci. 2019, 22, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Okada, S.; Sakurai, M. Adenosine-to-inosine RNA editing in neurological development and disease. RNA Biol. 2021, 18, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, I.; Tamir, H.; Arango, V.; Dwork, A.J.; Mann, J.J.; Schmauss, C. Altered editing of serotonin 2C receptor pre-mRNA in the prefrontal cortex of depressed suicide victims. Neuron 2002, 34, 349–356. [Google Scholar] [CrossRef]
- Chen, J.Y.; Peng, Z.; Zhang, R.; Yang, X.Z.; Tan, B.C.; Fang, H.; Liu, C.J.; Shi, M.; Ye, Z.Q.; Zhang, Y.E.; et al. RNA editome in rhesus macaque shaped by purifying selection. PLoS Genet. 2014, 10, e1004274. [Google Scholar] [CrossRef]
- Cuddleston, W.H.; Li, J.; Fan, X.; Kozenkov, A.; Lalli, M.; Khalique, S.; Dracheva, S.; Mukamel, E.A.; Breen, M.S. Cellular and genetic drivers of RNA editing variation in the human brain. Nat. Commun. 2022, 13, 2997. [Google Scholar] [CrossRef]
- Lundin, E.; Wu, C.; Widmark, A.; Behm, M.; Hjerling-Leffler, J.; Daniel, C.; Ohman, M.; Nilsson, M. Spatiotemporal mapping of RNA editing in the developing mouse brain using in situ sequencing reveals regional and cell-type-specific regulation. BMC Biol. 2020, 18, 6. [Google Scholar] [CrossRef] [PubMed]
- Jospin, M.; Watanabe, S.; Joshi, D.; Young, S.; Hamming, K.; Thacker, C.; Snutch, T.P.; Jorgensen, E.M.; Schuske, K. UNC-80 and the NCA ion channels contribute to endocytosis defects in synaptojanin mutants. Curr. Biol. 2007, 17, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
- Lear, B.C.; Darrah, E.J.; Aldrich, B.T.; Gebre, S.; Scott, R.L.; Nash, H.A.; Allada, R. UNC79 and UNC80, putative auxiliary subunits of the NARROW ABDOMEN ion channel, are indispensable for robust circadian locomotor rhythms in Drosophila. PLoS ONE 2013, 8, e78147. [Google Scholar] [CrossRef]
- Yeh, E.; Ng, S.; Zhang, M.; Bouhours, M.; Wang, Y.; Wang, M.; Hung, W.; Aoyagi, K.; Melnik-Martinez, K.; Li, M.; et al. A putative cation channel, NCA-1, and a novel protein, UNC-80, transmit neuronal activity in C. elegans. PLoS Biol. 2008, 6, e55. [Google Scholar] [CrossRef]
- Lu, B.; Zhang, Q.; Wang, H.; Wang, Y.; Nakayama, M.; Ren, D. Extracellular calcium controls background current and neuronal excitability via an UNC79-UNC80-NALCN cation channel complex. Neuron 2010, 68, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ren, D. UNC80 functions as a scaffold for Src kinases in NALCN channel function. Channels 2009, 3, 161–163. [Google Scholar] [CrossRef]
- Kang, Y.; Chen, L. Structure and mechanism of NALCN-FAM155A-UNC79-UNC80 channel complex. Nat. Commun. 2022, 13, 2639. [Google Scholar] [CrossRef]
- Wie, J.; Bharthur, A.; Wolfgang, M.; Narayanan, V.; Ramsey, K.; Group, C.R.R.; Aranda, K.; Zhang, Q.; Zhou, Y.; Ren, D. Intellectual disability-associated UNC80 mutations reveal inter-subunit interaction and dendritic function of the NALCN channel complex. Nat. Commun. 2020, 11, 3351. [Google Scholar] [CrossRef] [PubMed]
- Perez, Y.; Kadir, R.; Volodarsky, M.; Noyman, I.; Flusser, H.; Shorer, Z.; Gradstein, L.; Birnbaum, R.Y.; Birk, O.S. UNC80 mutation causes a syndrome of hypotonia, severe intellectual disability, dyskinesia and dysmorphism, similar to that caused by mutations in its interacting cation channel NALCN. J. Med. Genet. 2016, 53, 397–402. [Google Scholar] [CrossRef]
- Shamseldin, H.E.; Faqeih, E.; Alasmari, A.; Zaki, M.S.; Gleeson, J.G.; Alkuraya, F.S. Mutations in UNC80, Encoding Part of the UNC79-UNC80-NALCN Channel Complex, Cause Autosomal-Recessive Severe Infantile Encephalopathy. Am. J. Hum. Genet. 2016, 98, 210–215. [Google Scholar] [CrossRef]
- Stray-Pedersen, A.; Cobben, J.M.; Prescott, T.E.; Lee, S.; Cang, C.; Aranda, K.; Ahmed, S.; Alders, M.; Gerstner, T.; Aslaksen, K.; et al. Biallelic Mutations in UNC80 Cause Persistent Hypotonia, Encephalopathy, Growth Retardation, and Severe Intellectual Disability. Am. J. Hum. Genet. 2016, 98, 202–209. [Google Scholar] [CrossRef]
- Fahad Raza, M.; Anwar, M.; Husain, A.; Rizwan, M.; Li, Z.; Nie, H.; Hlavac, P.; Ali, M.A.; Rady, A.; Su, S. Differential gene expression analysis following olfactory learning in honeybee (Apis mellifera L.). PLoS ONE 2022, 17, e0262441. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Luo, J.; He, X.; Zhou, Q.; He, Y.; Wang, X.; Ma, L. The NALCN Channel Regulator UNC-80 Functions in a Subset of Interneurons To Regulate Caenorhabditis elegans Reversal Behavior. G3 2020, 10, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Cheng, Y.; Tan, B.C.; Kang, L.; Tian, Z.; Zhu, Y.; Zhang, W.; Liang, Y.; Hu, X.; Tan, X.; et al. Comprehensive analysis of RNA-Seq data reveals extensive RNA editing in a human transcriptome. Nat. Biotechnol. 2012, 30, 253–260. [Google Scholar] [CrossRef]
- Akdel, M.; Pires, D.E.; Pardo, E.P.; Jänes, J.; Zalevsky, A.O.; Mészáros, B.; Bryant, P.; Good, L.L.; Laskowski, R.A.; Pozzati, G. A structural biology community assessment of AlphaFold2 applications. Nat. Struct. Mol. Biol. 2022, 29, 1056–1067. [Google Scholar] [CrossRef]
- Bramucci, E.; Paiardini, A.; Bossa, F.; Pascarella, S. PyMod: Sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinform. 2012, 13, 1–6. [Google Scholar] [CrossRef]
- Aqrabawi, A.J.; Kim, J.C. Olfactory memory representations are stored in the anterior olfactory nucleus. Nat. Commun. 2020, 11, 1246. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, S.; Sato, K.; Kashiwadani, H.; Tsunoda, K.; Yamasoba, T.; Mori, K. Neurons in the anterior olfactory nucleus pars externa detect right or left localization of odor sources. Proc. Natl. Acad. Sci. USA 2010, 107, 12363–12368. [Google Scholar] [CrossRef] [PubMed]
- Dikeçligil, G.N.; Yang, A.I.; Sanghani, N.; Lucas, T.; Chen, H.I.; Davis, K.A.; Gottfried, J.A. Odor representations from the two nostrils are temporally segregated in human piriform cortex. Curr. Biol. 2023, 33, 5275–5287.e5275. [Google Scholar] [CrossRef]
- Bramswig, N.C.; Bertoli-Avella, A.M.; Albrecht, B.; Al Aqeel, A.I.; Alhashem, A.; Al-Sannaa, N.; Bah, M.; Brohl, K.; Depienne, C.; Dorison, N.; et al. Genetic variants in components of the NALCN-UNC80-UNC79 ion channel complex cause a broad clinical phenotype (NALCN channelopathies). Hum. Genet. 2018, 137, 753–768. [Google Scholar] [CrossRef]
- Yu, X.; Zou, J.; Babb, J.S.; Johnson, G.; Sanes, D.H.; Turnbull, D.H. Statistical mapping of sound-evoked activity in the mouse auditory midbrain using Mn-enhanced MRI. Neuroimage 2008, 39, 223–230. [Google Scholar] [CrossRef]
- Weng, J.-C.; Chen, J.-H.; Yang, P.-F.; Tseng, W.-Y.I. Functional mapping of rat barrel activation following whisker stimulation using activity-induced manganese-dependent contrast. Neuroimage 2007, 36, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Venkataraju, K.U.; Pradhan, K.; Mende, C.; Taranda, J.; Turaga, S.C.; Arganda-Carreras, I.; Ng, L.; Hawrylycz, M.J.; Rockland, K.S.; et al. Mapping social behavior-induced brain activation at cellular resolution in the mouse. Cell Rep. 2015, 10, 292–305. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, Y.; Yu, H.; Tian, Y.; Chen, X.; Chen, C.; Ren, Y.; Chen, Z.; Ren, Y.; Gong, X.; et al. Protective effect of Nr4a2 (Nurr1) against LPS-induced depressive-like behaviors via regulating activity of microglia and CamkII neurons in anterior cingulate cortex. Pharmacol. Res. 2023, 191, 106717. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, F.; Jia, D.; Bi, S.; Gong, J.; Wu, J.J.; Mao, Y.; Chen, J.; Chai, G.S. Pathological features and molecular signatures of early olfactory dysfunction in 3xTg-AD model mice. CNS Neurosci. Ther. 2024, 30, e14632. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Su, Y.; Das, S.; Wang, H.; Wang, Y.; Liu, J.; Ren, D. Peptide neurotransmitters activate a cation channel complex of NALCN and UNC-80. Nature 2009, 457, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Gao, S.; Alcaire, S.M.; Aoyagi, K.; Wang, Y.; Griffin, J.K.; Stagljar, I.; Nagamatsu, S.; Zhen, M. NLF-1 delivers a sodium leak channel to regulate neuronal excitability and modulate rhythmic locomotion. Neuron 2013, 77, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Su, Y.; Das, S.; Liu, J.; Xia, J.; Ren, D. The neuronal channel NALCN contributes resting sodium permeability and is required for normal respiratory rhythm. Cell 2007, 129, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, N.; Twik, M.; Plaschkes, I.; Nudel, R.; Iny Stein, T.; Levitt, J.; Gershoni, M.; Morrey, C.P.; Safran, M.; Lancet, D. MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2017, 45, D877–D887. [Google Scholar] [CrossRef]
- Canet-Pons, J.; Sen, N.-E.; Arsović, A.; Almaguer-Mederos, L.-E.; Halbach, M.V.; Key, J.; Döring, C.; Kerksiek, A.; Picchiarelli, G.; Cassel, R. Atxn2-CAG100-KnockIn mouse spinal cord shows progressive TDP43 pathology associated with cholesterol biosynthesis suppression. Neurobiol. Dis. 2021, 152, 105289. [Google Scholar] [CrossRef]
- Sternberg, J.R.; Prendergast, A.E.; Brosse, L.; Cantaut-Belarif, Y.; Thouvenin, O.; Orts-Del’Immagine, A.; Castillo, L.; Djenoune, L.; Kurisu, S.; McDearmid, J.R.; et al. Pkd2l1 is required for mechanoception in cerebrospinal fluid-contacting neurons and maintenance of spine curvature. Nat. Commun. 2018, 9, 3804. [Google Scholar] [CrossRef]
- Cao, L.; Huang, M.Z.; Zhang, Q.; Luo, Z.R.; Zhang, Y.; An, P.J.; Yang, L.L.; Tan, W.; Wang, C.Q.; Dou, X.W.; et al. The neural stem cell properties of Pkd2l1(+) cerebrospinal fluid-contacting neurons in vivo. Front. Cell. Neurosci. 2022, 16, 992520. [Google Scholar] [CrossRef] [PubMed]
- Hague, C.; Hall, R.A.; Minneman, K.P. Olfactory receptor localization and function: An emerging role for GPCR heterodimerization. Mol. Interv. 2004, 4, 321–322. [Google Scholar] [CrossRef]
- Spehr, M.; Munger, S.D. Olfactory receptors: G protein-coupled receptors and beyond. J. Neurochem. 2009, 109, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Cong, X.; Ren, W.; Pacalon, J.; Xu, R.; Xu, L.; Li, X.; de March, C.A.; Matsunami, H.; Yu, H.; Yu, Y.; et al. Large-Scale G Protein-Coupled Olfactory Receptor-Ligand Pairing. ACS Cent. Sci. 2022, 8, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Peralta-Yahya, P. Olfactory Receptors as an Emerging Chemical Sensing Scaffold. Biochemistry 2023, 62, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, F.A.; Chess, A. Olfactory G proteins: Simple and complex signal transduction. Curr. Biol. 1998, 8, R431–R433. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.; Sung, Y.K.; Reddy, R.; Ronnett, G.V. Odorants induce the phosphorylation of the cAMP response element binding protein in olfactory receptor neurons. Proc. Natl. Acad. Sci. USA 1999, 96, 14605–14610. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.J.; Chesler, A.; Firestein, S. How the olfactory bulb got its glomeruli: A just so story? Nat. Rev. Neurosci. 2009, 10, 611–618. [Google Scholar] [CrossRef]
- Francia, S.; Lodovichi, C. The role of the odorant receptors in the formation of the sensory map. BMC Biol. 2021, 19, 174. [Google Scholar] [CrossRef]
- Wang, X.; Younis, S.; Cen, J.; Wang, Y.; Krizhanovskii, C.; Andersson, L.; Welsh, N. ZBED6 counteracts high-fat diet-induced glucose intolerance by maintaining beta cell area and reducing excess mitochondrial activation. Diabetologia 2021, 64, 2292–2305. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhang, J.; Jiang, X.; Wang, W.; Chen, Y.Q. Free fatty acid receptor 4 deletion attenuates colitis by modulating Treg Cells via ZBED6-IL33 pathway. EBioMedicine 2022, 80, 104060. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Pan, D.; Li, P.; Wang, D.; Xia, B.; Zhang, R.; Lu, J.; Xing, X.; Du, J.; Zhang, X.; et al. Loss of ZBED6 Protects Against Sepsis-Induced Muscle Atrophy by Upregulating DOCK3-Mediated RAC1/PI3K/AKT Signaling Pathway in Pigs. Adv. Sci. 2023, 10, e2302298. [Google Scholar] [CrossRef]
- Mignot, C.; Nahrath, P.; Sinding, C.; Hummel, T. Older and Young Adults Experience Similar Long-Term Olfactory Habituation. Chem. Senses 2021, 46, bjaa080. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.-W.; Ma, C.-P.; Chin, E.; Chen, Y.-T.; Wang, T.-C.; Kuo, Y.-P.; Su, C.-H.; Huang, P.-J.; Tan, B.C.-M. Imbalance in Unc80 RNA Editing Disrupts Dynamic Neuronal Activity and Olfactory Perception. Int. J. Mol. Sci. 2024, 25, 5985. https://doi.org/10.3390/ijms25115985
Chen H-W, Ma C-P, Chin E, Chen Y-T, Wang T-C, Kuo Y-P, Su C-H, Huang P-J, Tan BC-M. Imbalance in Unc80 RNA Editing Disrupts Dynamic Neuronal Activity and Olfactory Perception. International Journal of Molecular Sciences. 2024; 25(11):5985. https://doi.org/10.3390/ijms25115985
Chicago/Turabian StyleChen, Hui-Wen, Chung-Pei Ma, En Chin, Yi-Tung Chen, Teh-Cheng Wang, Yu-Ping Kuo, Chia-Hao Su, Po-Jung Huang, and Bertrand Chin-Ming Tan. 2024. "Imbalance in Unc80 RNA Editing Disrupts Dynamic Neuronal Activity and Olfactory Perception" International Journal of Molecular Sciences 25, no. 11: 5985. https://doi.org/10.3390/ijms25115985
APA StyleChen, H.-W., Ma, C.-P., Chin, E., Chen, Y.-T., Wang, T.-C., Kuo, Y.-P., Su, C.-H., Huang, P.-J., & Tan, B. C.-M. (2024). Imbalance in Unc80 RNA Editing Disrupts Dynamic Neuronal Activity and Olfactory Perception. International Journal of Molecular Sciences, 25(11), 5985. https://doi.org/10.3390/ijms25115985