
Dynamics of Cellular Regulation of Fractalkine/CX3CL1 and Its Receptor CX3CR1 in the Rat Trigeminal Subnucleus Caudalis after Unilateral Infraorbital Nerve Lesion—Extended Cellular Signaling of the CX3CL1/CX3CR1 Axis in the Development of Trigeminal Neuropathic Pain


Abstract
1. Introduction
2. Results
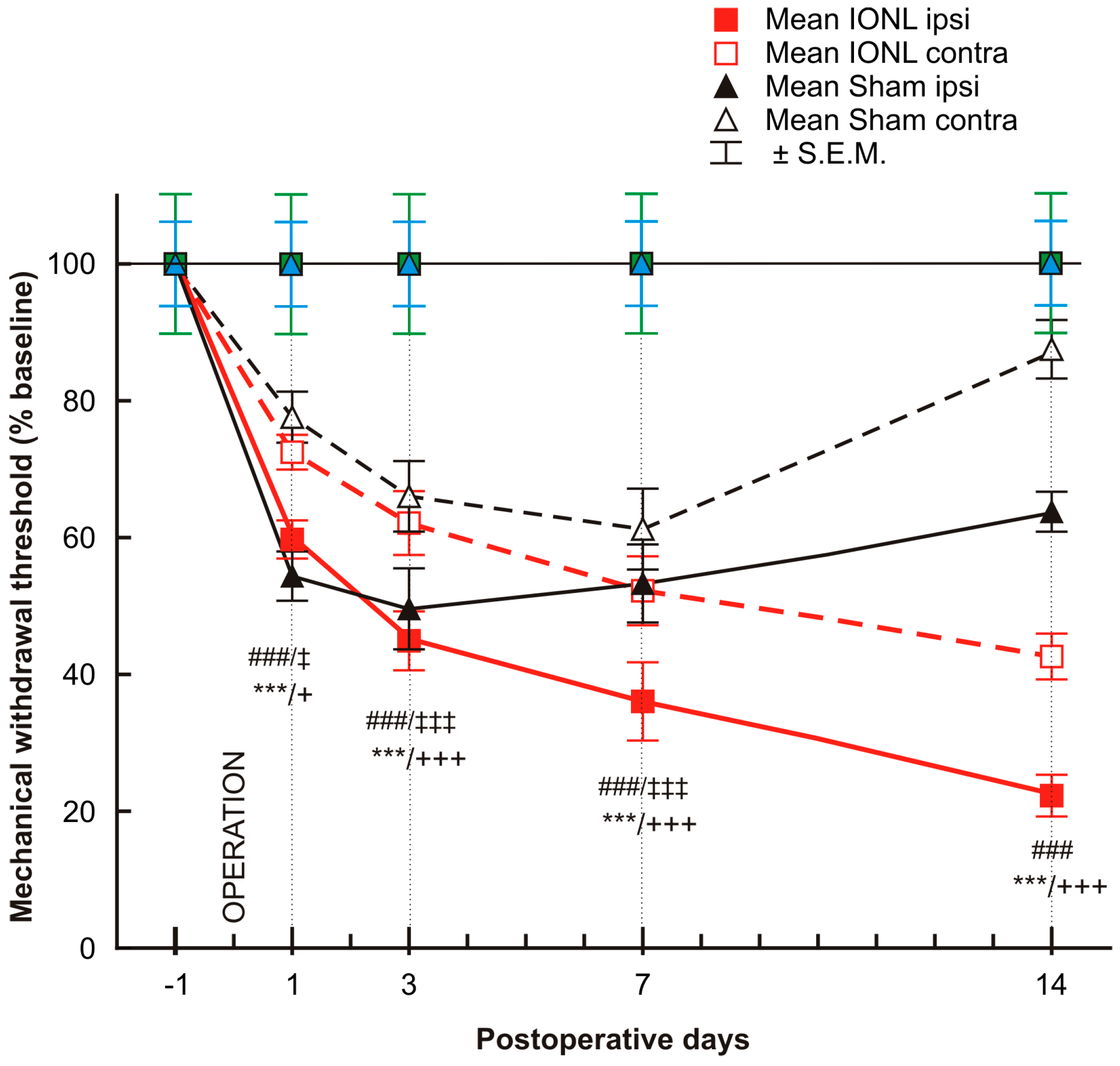
2.1. Behavioral Tests
2.2. Cellular Distribution of CX3CL1 and CX3CR1 in the TSC during the Development of Trigeminal Neuropathic Pain
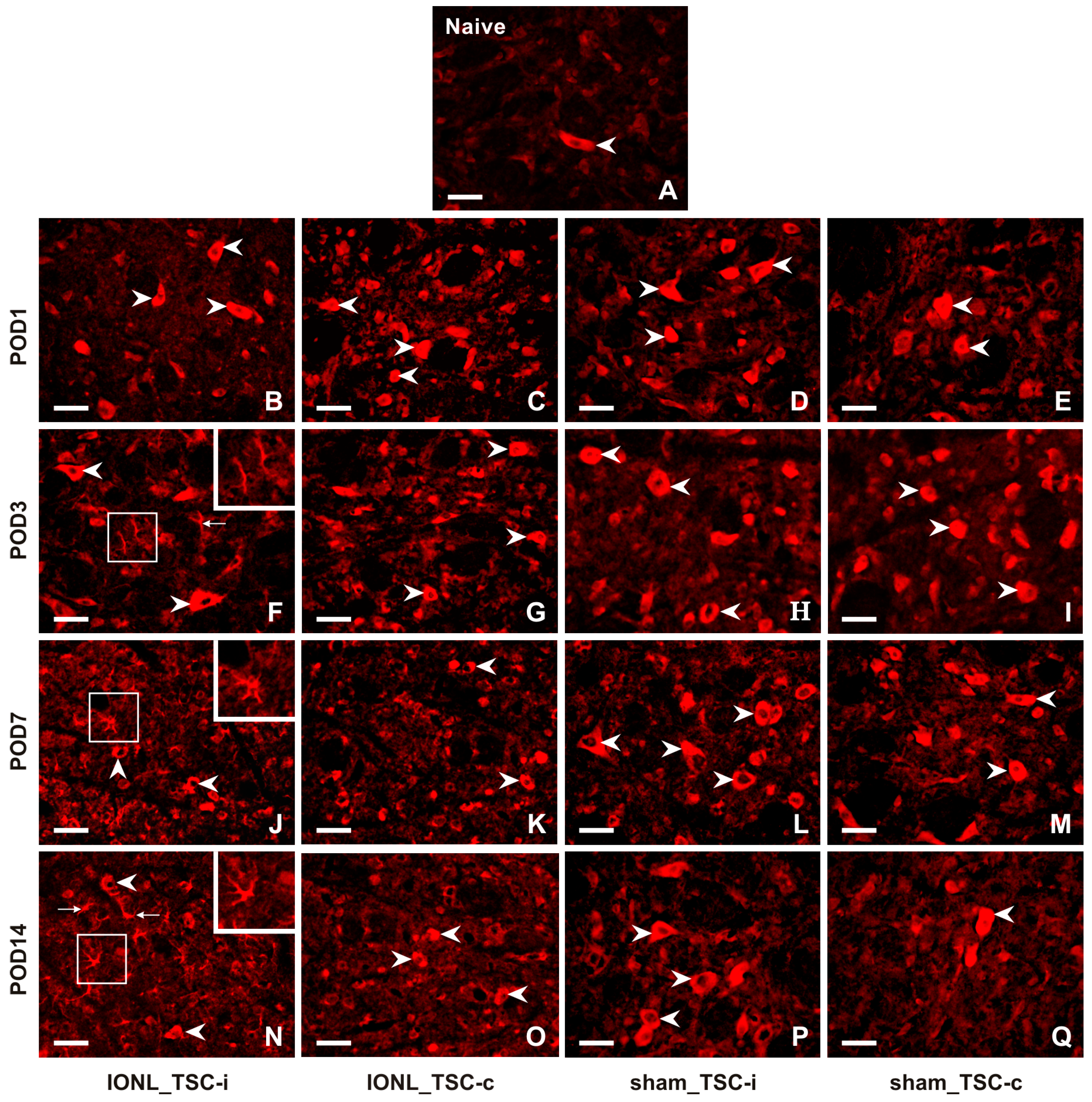
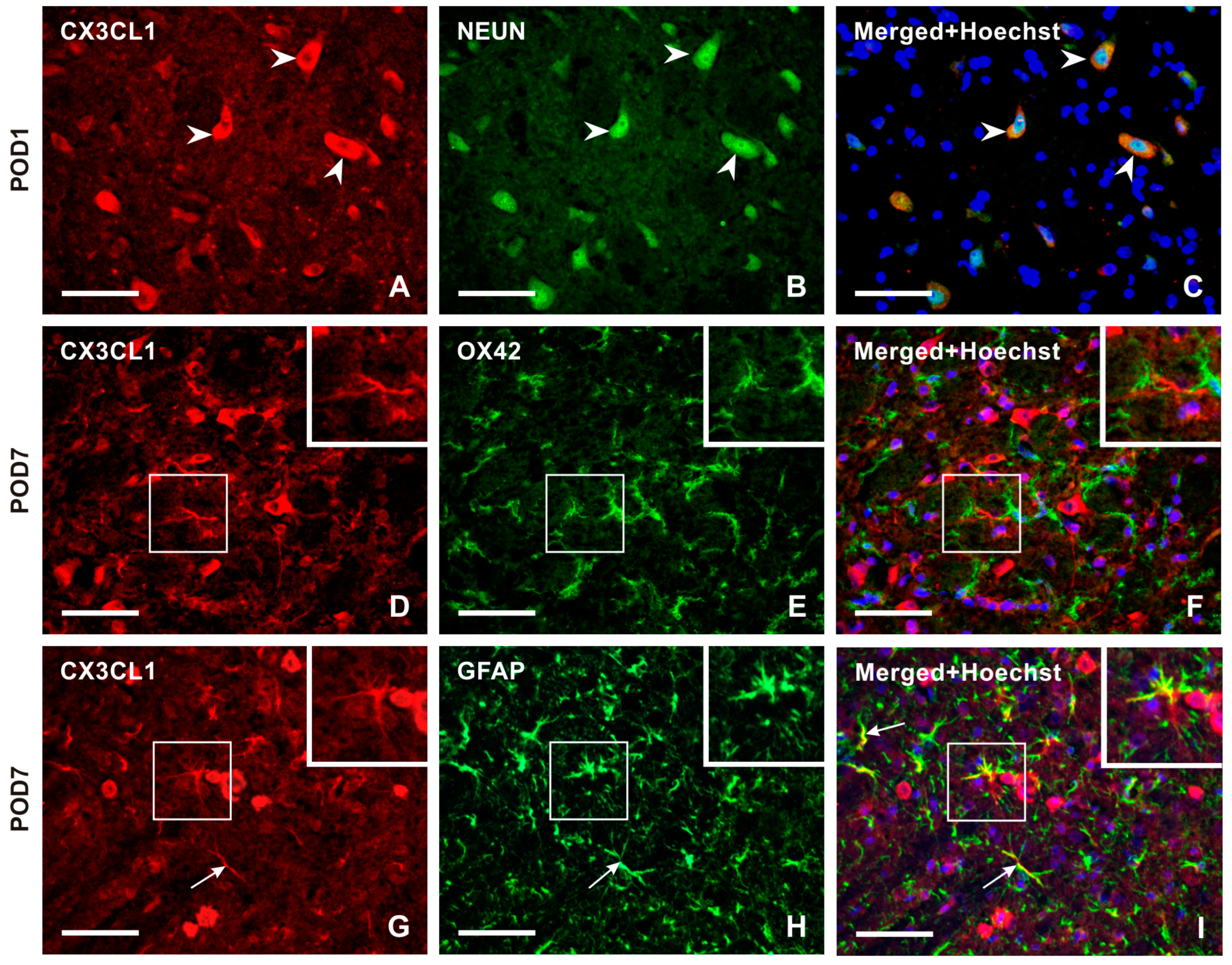
2.2.1. Cellular Distribution of CX3CL1 in the TSC of Naïve, Sham-Operated, and IONL-Operated Rats
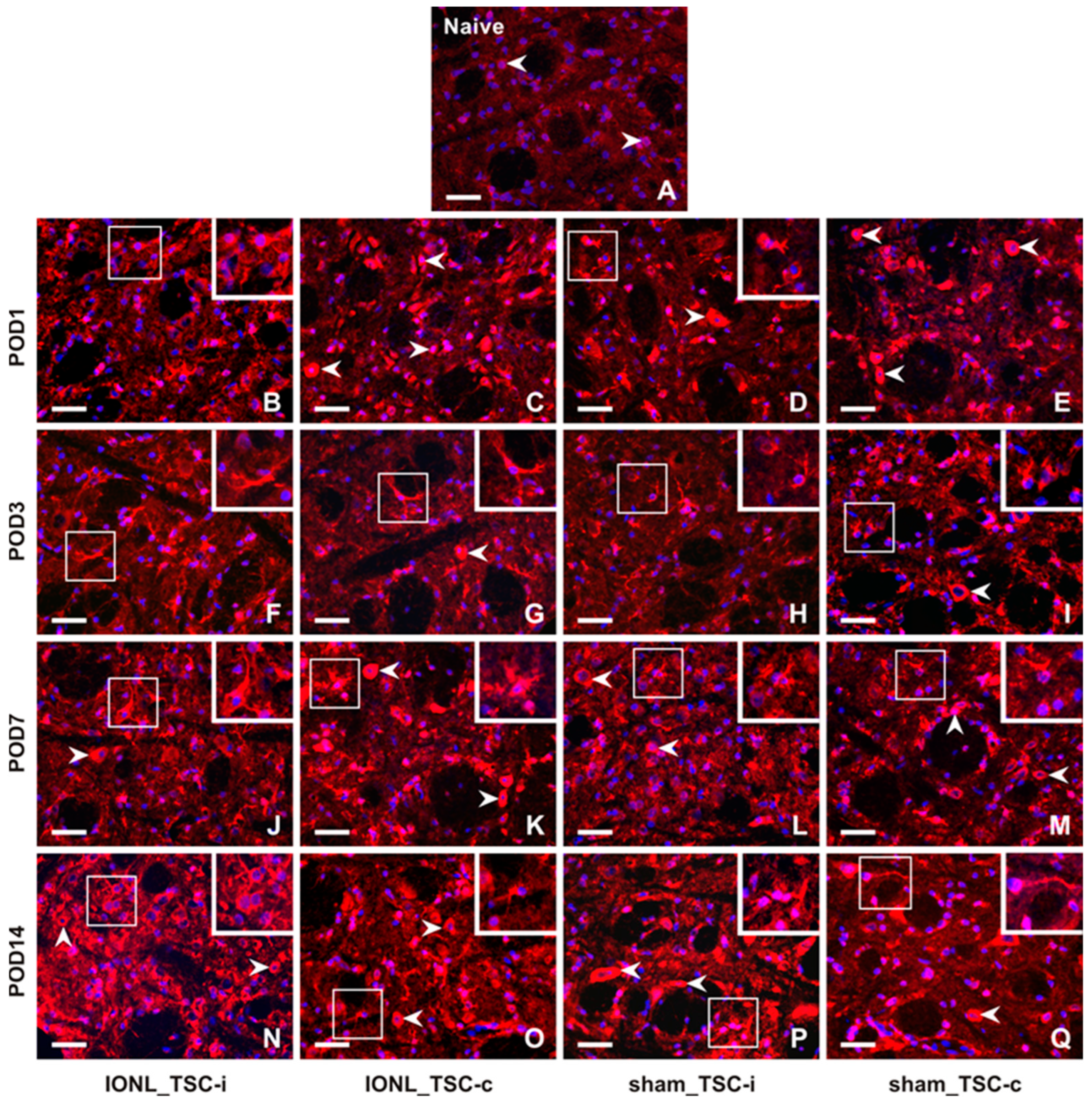
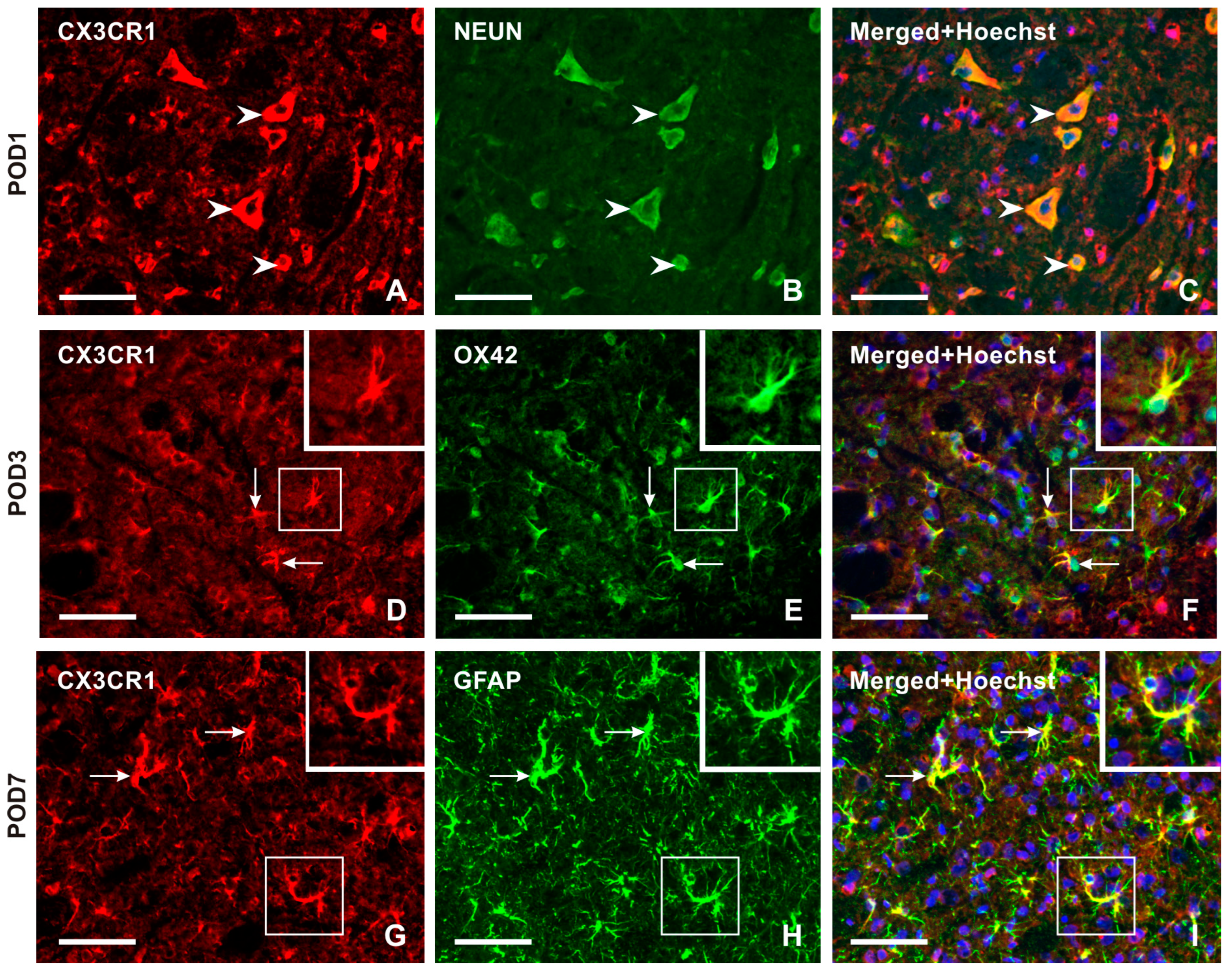
2.2.2. Cellular Distribution of CX3CR1 in the TSC of Naïve, Sham-Operated, and IONL-Operated Rats
2.3. Cellular Distribution of CatS and ADAM17 in the TSC during the Development of Trigeminal Neuropathic Pain
2.3.1. Cellular Distribution of CatS in the TSC of Naïve, Sham-Operated, and IONL-Operated Rats
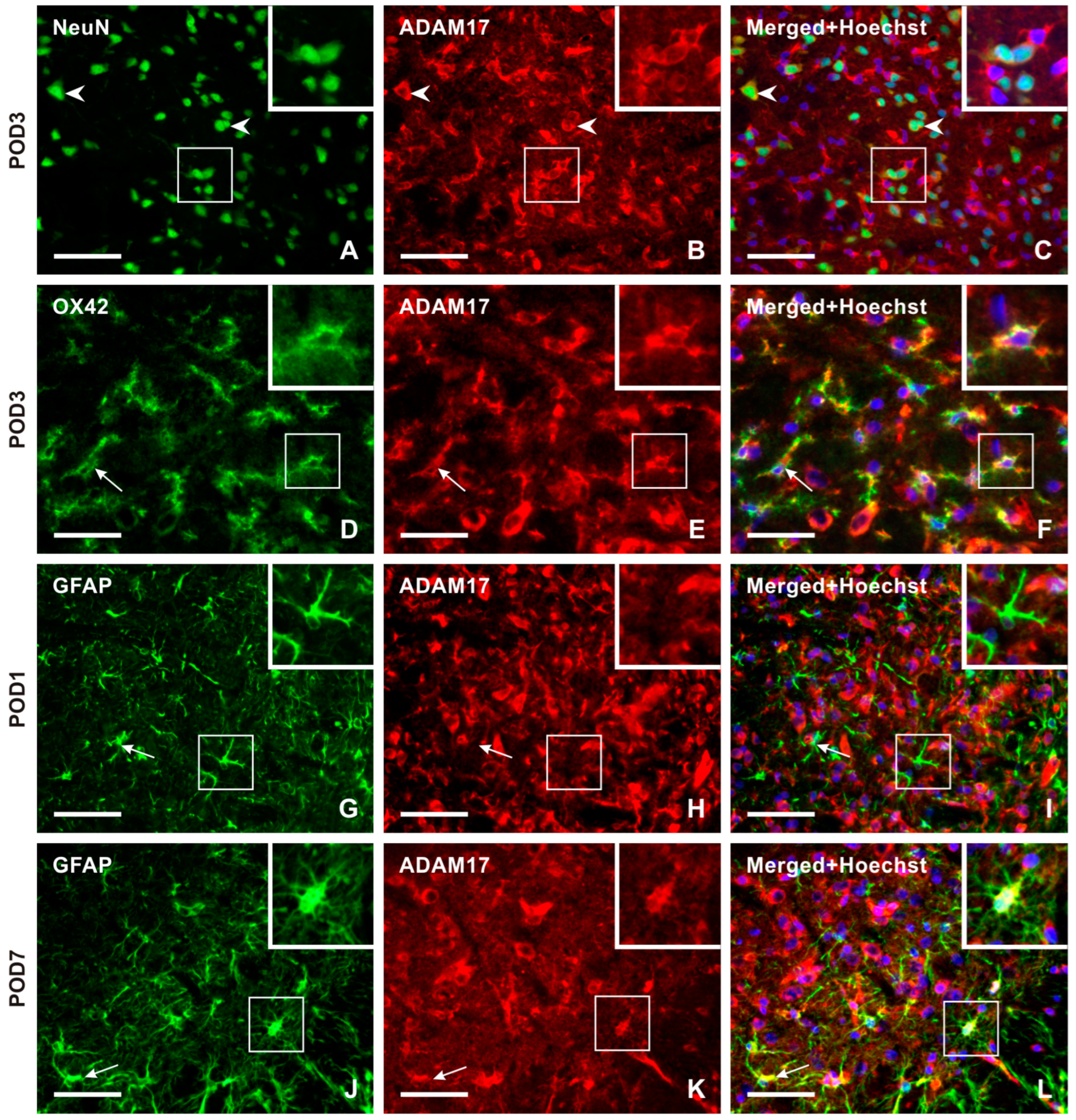
2.3.2. Cellular Distribution of ADAM17 in the TSC of Naïve, Sham-Operated, and IONL-Operated Rats
2.4. Summary of the Semiquantitative Evaluation of Immunofluorescence Intensities
3. Discussion
3.1. Cellular Distribution of CX3CL1/CX3CR1 in the TSC
3.2. Cellular Origin of Soluble CX3CL1 Production in the TSC
4. Materials and Methods
4.1. Animals and Surgical Procedures
4.2. Behavioral Tests of Mechanical Hypersensitivity
4.3. Tissue Microarrays of CX3CL1 and CX3CR1 Immunofluorescence
4.4. Double Immunostaining
4.5. Immunofluorescence Microscopy and Image Analysis
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gregg, J.M.; Dixon, A.D. Somatotopic Organization of the Trigeminal Ganglion in the Rat. Arch. Oral Biol. 1973, 18, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Chichorro, J.G.; Porreca, F.; Sessle, B. Mechanisms of Craniofacial Pain. Cephalalgia 2017, 37, 613–626. [Google Scholar] [CrossRef]
- Inoue, K.; Tsuda, M. Microglia and Neuropathic Pain. Glia 2009, 57, 1469–1479. [Google Scholar] [CrossRef]
- Kubíčková, L.; Klusáková, I.; Dubový, P. Bilateral Activation of Glial Cells and Cellular Distribution of the Chemokine CCL2 and Its Receptor CCR2 in the Trigeminal Subnucleus Caudalis of Trigeminal Neuropathic Pain Model. Histochem. Cell Biol. 2020, 153, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Okada-Ogawa, A.; Suzuki, I.; Sessle, B.J.; Chiang, C.Y.; Salter, M.W.; Dostrovsky, J.O.; Tsuboi, Y.; Kondo, M.; Kitagawa, J.; Kobayashi, A.; et al. Astroglia in Medullary Dorsal Horn (Trigeminal Spinal Subnucleus Caudalis) Are Involved in Trigeminal Neuropathic Pain Mechanisms. J. Neurosci. 2009, 29, 11161–11171. [Google Scholar] [CrossRef] [PubMed]
- Shibuta, K.; Suzuki, I.; Shinoda, M.; Tsuboi, Y.; Honda, K.; Shimizu, N.; Sessle, B.J.; Iwata, K. Organization of Hyperactive Microglial Cells in Trigeminal Spinal Subnucleus Caudalis and Upper Cervical Spinal Cord Associated with Orofacial Neuropathic Pain. Brain Res. 2012, 1451, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Dauvergne, C.; Molet, J.; Reaux-Le Goazigo, A.; Mauborgne, A.; Melik-Parsadaniantz, S.; Boucher, Y.; Pohl, M. Implication of the Chemokine CCL2 in Trigeminal Nociception and Traumatic Neuropathic Orofacial Pain. Eur. J. Pain 2014, 18, 360–375. [Google Scholar] [CrossRef]
- Takeda, M.; Nasu, M.; Kanazawa, T.; Takahashi, M.; Shimazu, Y. Chemokine Ligand 2/Chemokine Receptor 2 Signaling in the Trigeminal Ganglia Contributes to Inflammatory Hyperalgesia in Rats. Neurosci. Res. 2018, 128, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cao, D.-L.; Zhang, Z.-J.; Jiang, B.-C.; Gao, Y.-J. Chemokine CXCL13 Mediates Orofacial Neuropathic Pain via CXCR5/ERK Pathway in the Trigeminal Ganglion of Mice. J. Neuroinflamm. 2016, 13, 183. [Google Scholar] [CrossRef]
- Abbadie, C.; Bhangoo, S.; De Koninck, Y.; Malcangio, M.; Melik-Parsadaniantz, S.; White, F.A. Chemokines and Pain Mechanisms. Brain Res. Rev. 2009, 60, 125–134. [Google Scholar] [CrossRef]
- Gao, Y.-J.; Ji, R.-R. Chemokines, Neuronal–Glial Interactions, and Central Processing of Neuropathic Pain. Pharmacol. Ther. 2010, 126, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Old, E.A.; Malcangio, M. Chemokine Mediated Neuron-Glia Communication and Aberrant Signalling in Neuropathic Pain States. Curr. Opin. Pharmacol. 2012, 12, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-J.; Jiang, B.-C.; Gao, Y.-J. Chemokines in Neuron–Glial Cell Interaction and Pathogenesis of Neuropathic Pain. Cell. Mol. Life Sci. 2017, 74, 3275–3291. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.K.; Malcangio, M. Fractalkine/CX3CR1 Signaling during Neuropathic Pain. Front. Cell. Neurosci. 2014, 8, 121. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.-M.; Lee, K.-M.; Cho, H.-J. Expression of Monocyte Chemoattractant Protein-1 in Rat Dorsal Root Ganglia and Spinal Cord in Experimental Models of Neuropathic Pain. Brain Res. 2009, 1251, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Verge, G.M.; Milligan, E.D.; Maier, S.F.; Watkins, L.R.; Naeve, G.S.; Foster, A.C. Fractalkine (CX3CL1) and Fractalkine Receptor (CX3CR1) Distribution in Spinal Cord and Dorsal Root Ganglia under Basal and Neuropathic Pain Conditions. Eur. J. Neurosci. 2004, 20, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-J.; Dong, Y.-L.; Lu, Y.; Cao, S.; Zhao, Z.-Q.; Gao, Y.-J. Chemokine CCL2 and Its Receptor CCR2 in the Medullary Dorsal Horn Are Involved in Trigeminal Neuropathic Pain. J. Neuroinflamm. 2012, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, M.; Pino, P.A.; Saederup, N.; Charo, I.F.; Ransohoff, R.M.; Cardona, A.E. The Fractalkine Receptor but Not CCR2 Is Present on Microglia from Embryonic Development throughout Adulthood. J. Immunol. 2012, 188, 29–36. [Google Scholar] [CrossRef]
- Kiyomoto, M.; Shinoda, M.; Okada-Ogawa, A.; Noma, N.; Shibuta, K.; Tsuboi, Y.; Sessle, B.J.; Imamura, Y.; Iwata, K. Fractalkine Signaling in Microglia Contributes to Ectopic Orofacial Pain Following Trapezius Muscle Inflammation. J. Neurosci. 2013, 33, 7667–7680. [Google Scholar] [CrossRef]
- Clark, A.K.; Malcangio, M. Microglial Signalling Mechanisms: Cathepsin S and Fractalkine. Exp. Neurol. 2012, 234, 283–292. [Google Scholar] [CrossRef]
- Abraira, V.E.; Ginty, D.D. The Sensory Neurons of Touch. Neuron 2013, 79, 619–639. [Google Scholar] [CrossRef] [PubMed]
- Bereiter, D.A.; Hirata, H.; Hu, J.W. Trigeminal Subnucleus Caudalis: Beyond Homologies with the Spinal Dorsal Horn. Pain 2000, 88, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, J. On the Anatomical and Functional Organization of the Spinal Trigeminal Nucleus. J. Comp. Neurol. 1950, 92, 401–413. [Google Scholar] [CrossRef]
- Sessle, B.J. Acute and Chronic Craniofacial Pain: Brainstem Mechanisms of Nociceptive Transmission and Neuroplasticity, and Their Clinical Correlates. Crit. Rev. Oral Biol. Med. 2000, 11, 57–91. [Google Scholar] [CrossRef] [PubMed]
- D’amico-Martel, A.; Noden, D.M. Contributions of Placodal and Neural Crest Cells to Avian Cranial Peripheral Ganglia. Am. J. Anat. 1983, 166, 445–468. [Google Scholar] [CrossRef] [PubMed]
- Kogelman, L.J.A.; Christensen, R.E.; Pedersen, S.H.; Bertalan, M.; Hansen, T.F.; Jansen-Olesen, I.; Olesen, J. Whole Transcriptome Expression of Trigeminal Ganglia Compared to Dorsal Root Ganglia in Rattus Norvegicus. Neuroscience 2017, 350, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Korczeniewska, O.A.; Rider, G.K.; Gajra, S.; Narra, V.; Ramavajla, V.; Chang, Y.-J.; Tao, Y.; Soteropoulos, P.; Husain, S.; Khan, J.; et al. Differential Gene Expression Changes in the Dorsal Root versus Trigeminal Ganglia Following Peripheral Nerve Injury in Rats. Eur. J. Pain 2020, 24, 967–982. [Google Scholar] [CrossRef] [PubMed]
- Lindia, J.A.; McGowan, E.; Jochnowitz, N.; Abbadie, C. Induction of CX3CL1 Expression in Astrocytes and CX3CR1 in Microglia in the Spinal Cord of a Rat Model of Neuropathic Pain. J. Pain 2005, 6, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Zhu, X.; He, Y.; Zhu, M.; Wu, Z.; Huang, F.; He, H. The Role of Satellite Glial Cells in Orofacial Pain. J. Neurosci. Res. 2019, 97, 393–401. [Google Scholar] [CrossRef]
- Bonfante, R.; Napimoga, M.H.; Macedo, C.G.; Abdalla, H.B.; Pieroni, V.; Clemente-Napimoga, J.T. The P2 × 7 Receptor, Cathepsin S and Fractalkine in the Trigeminal Subnucleus Caudalis Signal Persistent Hypernociception in Temporomandibular Rat Joints. Neuroscience 2018, 391, 120–130. [Google Scholar] [CrossRef]
- Cairns, B.E.; O’Brien, M.; Dong, X.-D.; Gazerani, P. Elevated Fractalkine (CX3CL1) Levels in the Trigeminal Ganglion Mechanically Sensitize Temporalis Muscle Nociceptors. Mol. Neurobiol. 2017, 54, 3695–3706. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.K.; Yip, P.K.; Malcangio, M. The Liberation of Fractalkine in the Dorsal Horn Requires Microglial Cathepsin S. J. Neurosci. 2009, 29, 6945–6954. [Google Scholar] [CrossRef] [PubMed]
- Meucci, O.; Fatatis, A.; Simen, A.A.; Miller, R.J. Expression of CX3CR1 Chemokine Receptors on Neurons and Their Role in Neuronal Survival. Proc. Natl. Acad. Sci. USA 2000, 97, 8075–8080. [Google Scholar] [CrossRef] [PubMed]
- Ragozzino, D.; Angelantonio, S.D.; Trettel, F.; Bertollini, C.; Maggi, L.; Gross, C.; Charo, I.F.; Limatola, C.; Eusebi, F. Chemokine Fractalkine/CX3CL1 Negatively Modulates Active Glutamatergic Synapses in Rat Hippocampal Neurons. J. Neurosci. 2006, 26, 10488–10498. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.V.; Falnikar, A.; Heinsinger, N.; Cheng, L.; Andrews, C.E.; DeMarco, M.; Lepore, A.C. Cervical Spinal Cord Injury-Induced Neuropathic Pain in Male Mice Is Associated with a Persistent pro-Inflammatory Macrophage/Microglial Response in the Superficial Dorsal Horn. Exp. Neurol. 2021, 343, 113757. [Google Scholar] [CrossRef] [PubMed]
- Hulshof, S.; van Haastert, E.S.; Kuipers, H.F.; van den Elsen, P.J.; de Groot, C.J.; van der Valk, P.; Ravid, R.; Biber, K. CX3CL1 and CX3CR1 Expression in Human Brain Tissue: Noninflammatory Control versus Multiple Sclerosis. J. Neuropathol. Exp. Neurol. 2003, 62, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Catalano, M.; Lauro, C.; Cipriani, R.; Chece, G.; Ponzetta, A.; Di Angelantonio, S.; Ragozzino, D.; Limatola, C. CX3CL1 Protects Neurons against Excitotoxicity Enhancing GLT-1 Activity on Astrocytes. J. Neuroimmunol. 2013, 263, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Limatola, C.; Lauro, C.; Catalano, M.; Ciotti, M.T.; Bertollini, C.; Di Angelantonio, S.; Ragozzino, D.; Eusebi, F. Chemokine CX3CL1 Protects Rat Hippocampal Neurons against Glutamate-Mediated Excitotoxicity. J. Neuroimmunol. 2005, 166, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Song, L.; Tan, Y.; Ma, Y.; Tian, Y.; Jin, X.; Lim, G.; Zhang, S.; Chen, L.; Mao, J. A Functional Relationship Between Trigeminal Astroglial Activation and NR1 Expression in a Rat Model of Temporomandibular Joint Inflammation. Pain Med. 2012, 13, 1590–1600. [Google Scholar] [CrossRef]
- Jung, S.; Aliberti, J.; Graemmel, P.; Sunshine, M.J.; Kreutzberg, G.W.; Sher, A.; Littman, D.R. Analysis of Fractalkine Receptor CX3CR1 Function by Targeted Deletion and Green Fluorescent Protein Reporter Gene Insertion. Mol. Cell. Biol. 2000, 20, 4106–4114. [Google Scholar] [CrossRef]
- Garton, K.J.; Gough, P.J.; Blobel, C.P.; Murphy, G.; Greaves, D.R.; Dempsey, P.J.; Raines, E.W. Tumor Necrosis Factor-α-Converting Enzyme (ADAM17) Mediates the Cleavage and Shedding of Fractalkine (CX3CL1). J. Biol. Chem. 2001, 276, 37993–38001. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.K.; Gentry, C.; Bradbury, E.J.; McMahon, S.B.; Malcangio, M. Role of Spinal Microglia in Rat Models of Peripheral Nerve Injury and Inflammation. Eur. J. Pain Lond. Engl. 2007, 11, 223–230. [Google Scholar] [CrossRef]
- Luo, P.; Chu, S.; Zhang, Z.; Xia, C.; Chen, N. Fractalkine/CX3CR1 Is Involved in the Cross-Talk between Neuron and Glia in Neurological Diseases. Brain Res. Bull. 2019, 146, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Vos, B.P.; Strassman, A.M.; Maciewicz, R.J. Behavioral Evidence of Trigeminal Neuropathic Pain Following Chronic Constriction Injury to the Rat’s Infraorbital Nerve. J. Neurosci. 1994, 14, 2708–2723. [Google Scholar] [CrossRef] [PubMed]
- Kayser, V.; Latrémolière, A.; Hamon, M.; Bourgoin, S. N-Methyl-D-Aspartate Receptor-Mediated Modulations of the Anti-Allodynic Effects of 5-HT1B/1D Receptor Stimulation in a Rat Model of Trigeminal Neuropathic Pain. Eur. J. Pain Lond. Engl. 2011, 15, 451–458. [Google Scholar] [CrossRef]
- Zamboni, L.; Demartin, C. Buffered picric acid-formaldehyde—A new rapid fixative for electron microscopy. J. Cell Biol. 1967, 35, A148. [Google Scholar]
- Montague-Cardoso, K.; Mrozkova, P.; Malcangio, M. The Role of Spinal Cord CX3CL1/CX3CR1 Signalling in Chronic Pain. Curr. Tissue Microenviron. Rep. 2020, 1, 23–29. [Google Scholar] [CrossRef]
- Staniland, A.A.; Clark, A.K.; Wodarski, R.; Sasso, O.; Maione, F.; D’Acquisto, F.; Malcangio, M. Reduced Inflammatory and Neuropathic Pain and Decreased Spinal Microglial Response in Fractalkine Receptor (CX3CR1) Knockout Mice. J. Neurochem. 2010, 114, 1143–1157. [Google Scholar] [CrossRef]
- Yin, Q.; Cheng, W.; Cheng, M.-Y.; Fan, S.-Z.; Shen, W. Intrathecal Injection of Anti-CX3CR1 Neutralizing Antibody Delayed and Attenuated Pain Facilitation in Rat Tibial Bone Cancer Pain Model. Behav. Pharmacol. 2010, 21, 595–601. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Product | Dilution | Conditions | |
|---|---|---|---|---|---|
| CX3CL1 | pAb | Rabbit | Abcam | 1:200 | 18 h |
| CX3CR1 | pAb | Rabbit | Abcam | 1:200 | 18 h |
| ED1 | pAb | Mouse | Serotec | 1:300 | 18 h |
| NEUN | mAb | Mouse | Abcam | 1:500 | 240 min |
| NEUN | mAb | Mouse | Chemicon | 1:100 | 18 h |
| OX42 | mAb | Mouse | Santa Cruz | 1:50 | 18 h |
| OX42 | pAb | Chicken | MyBioSource | 1:500 | 18 h |
| GFAP | pAb | Chicken | Abcam | 1:500 | 180 min |
| CatS | pAb | Rabbit | LSbio | 1:500 | 18 h |
| ADAM17 | pAb | Rabbit | Chemicon | 1:200 | 18 h |
| Range of Pixel Intensities | Immunofluorescence Intensity |
|---|---|
| 0–9 | None |
| 10–84 | Very weak |
| 85–169 | Moderate |
| 170–255 | High |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubíčková, L.; Dubový, P. Dynamics of Cellular Regulation of Fractalkine/CX3CL1 and Its Receptor CX3CR1 in the Rat Trigeminal Subnucleus Caudalis after Unilateral Infraorbital Nerve Lesion—Extended Cellular Signaling of the CX3CL1/CX3CR1 Axis in the Development of Trigeminal Neuropathic Pain. Int. J. Mol. Sci. 2024, 25, 6069. https://doi.org/10.3390/ijms25116069
Kubíčková L, Dubový P. Dynamics of Cellular Regulation of Fractalkine/CX3CL1 and Its Receptor CX3CR1 in the Rat Trigeminal Subnucleus Caudalis after Unilateral Infraorbital Nerve Lesion—Extended Cellular Signaling of the CX3CL1/CX3CR1 Axis in the Development of Trigeminal Neuropathic Pain. International Journal of Molecular Sciences. 2024; 25(11):6069. https://doi.org/10.3390/ijms25116069
Chicago/Turabian StyleKubíčková, Lucie, and Petr Dubový. 2024. "Dynamics of Cellular Regulation of Fractalkine/CX3CL1 and Its Receptor CX3CR1 in the Rat Trigeminal Subnucleus Caudalis after Unilateral Infraorbital Nerve Lesion—Extended Cellular Signaling of the CX3CL1/CX3CR1 Axis in the Development of Trigeminal Neuropathic Pain" International Journal of Molecular Sciences 25, no. 11: 6069. https://doi.org/10.3390/ijms25116069
APA StyleKubíčková, L., & Dubový, P. (2024). Dynamics of Cellular Regulation of Fractalkine/CX3CL1 and Its Receptor CX3CR1 in the Rat Trigeminal Subnucleus Caudalis after Unilateral Infraorbital Nerve Lesion—Extended Cellular Signaling of the CX3CL1/CX3CR1 Axis in the Development of Trigeminal Neuropathic Pain. International Journal of Molecular Sciences, 25(11), 6069. https://doi.org/10.3390/ijms25116069