
Next-Generation Sequencing of the Human Aqueous Humour Microbiome

 ,
,
Abstract
1. Introduction
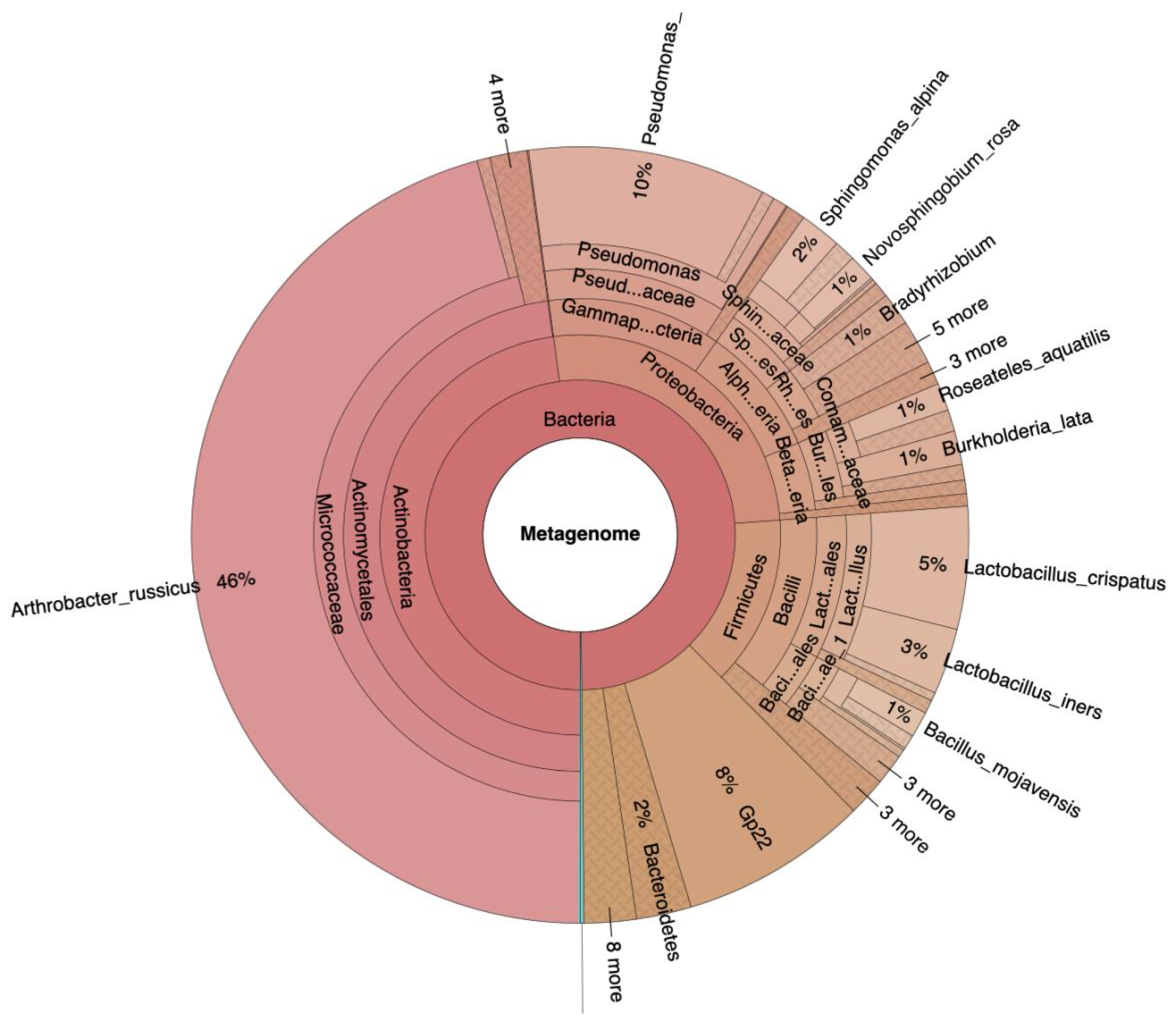
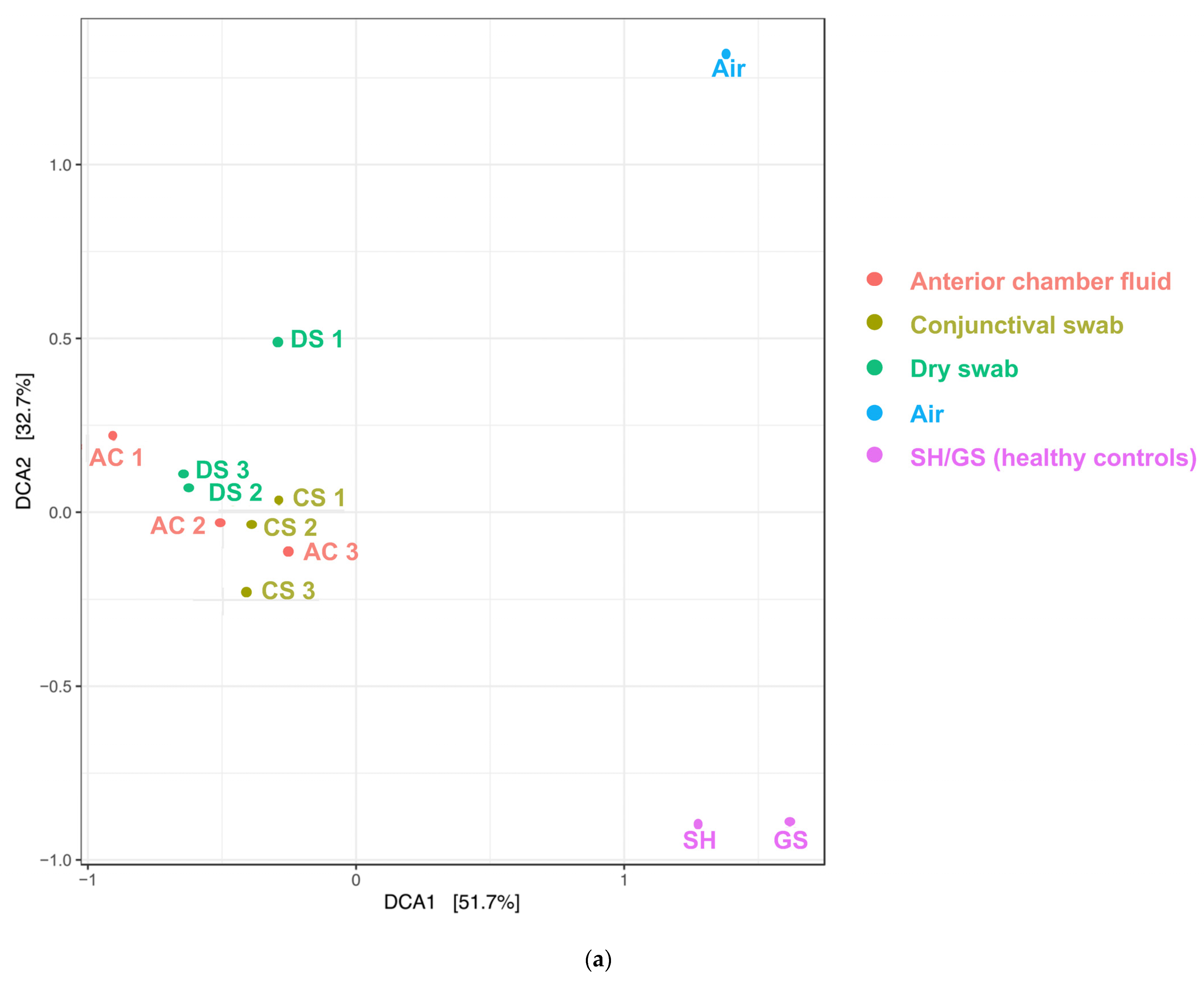
2. Results
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Sampling
4.3. Sample Processing
4.4. Biostatistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed]
- de Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C.S. Human Placenta Has No Microbiome but Can Contain Potential Pathogens. Nature 2019, 572, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Characterization of the Bacterial Microbiome in First-Pass Meconium Using Propidium Monoazide (PMA) to Exclude Nonviable Bacterial DNA. Lett. Appl. Microbiol. 2019, 68, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Doan, T.; Akileswaran, L.; Andersen, D.; Johnson, B.; Ko, N.; Shrestha, A.; Shestopalov, V.; Lee, C.S.; Lee, A.Y.; Van Gelder, R.N. Paucibacterial Microbiome and Resident DNA Virome of the Healthy Conjunctiva. Investig. Opthalmol. Vis. Sci. 2016, 57, 5116. [Google Scholar] [CrossRef] [PubMed]
- Delbeke, H.; Younas, S.; Casteels, I.; Joossens, M. Current Knowledge on the Human Eye Microbiome: A Systematic Review of Available Amplicon and Metagenomic Sequencing Data. Acta Ophthalmol. 2021, 99, 16–25. [Google Scholar] [CrossRef]
- Dong, Q.; Brulc, J.M.; Iovieno, A.; Bates, B.; Garoutte, A.; Miller, D.; Revanna, K.V.; Gao, X.; Antonopoulos, D.A.; Slepak, V.Z.; et al. Diversity of Bacteria at Healthy Human Conjunctiva. Investig. Opthalmol. Vis. Sci. 2011, 52, 5408. [Google Scholar] [CrossRef]
- Katzka, W.; Dong, T.S.; Luu, K.; Lagishetty, V.; Sedighian, F.; Arias-Jayo, N.; Jacobs, J.P.; Hsu, H.Y. The Ocular Microbiome Is Altered by Sampling Modality and Age. Trans. Vis. Sci. Technol. 2021, 10, 24. [Google Scholar] [CrossRef]
- Ozkan, J.; Nielsen, S.; Diez-Vives, C.; Coroneo, M.; Thomas, T.; Willcox, M. Temporal Stability and Composition of the Ocular Surface Microbiome. Sci. Rep. 2017, 7, 9880. [Google Scholar] [CrossRef]
- Wen, X.; Miao, L.; Deng, Y.; Bible, P.W.; Hu, X.; Zou, Y.; Liu, Y.; Guo, S.; Liang, J.; Chen, T.; et al. The Influence of Age and Sex on Ocular Surface Microbiota in Healthy Adults. Investig. Ophthalmol. Vis. Sci. 2017, 58, 6030–6037. [Google Scholar] [CrossRef]
- Matysiak, A.; Kabza, M.; Karolak, J.A.; Jaworska, M.M.; Rydzanicz, M.; Ploski, R.; Szaflik, J.P.; Gajecka, M. Characterization of Ocular Surface Microbial Profiles Revealed Discrepancies between Conjunctival and Corneal Microbiota. Pathogens 2021, 10, 405. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, J.; Coroneo, M.; Willcox, M.; Wemheuer, B.; Thomas, T. Identification and Visualization of a Distinct Microbiome in Ocular Surface Conjunctival Tissue. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4268–4276. [Google Scholar] [CrossRef]
- Borroni, D.; Romano, V.; Kaye, S.B.; Somerville, T.; Napoli, L.; Fasolo, A.; Gallon, P.; Ponzin, D.; Esposito, A.; Ferrari, S. Metagenomics in Ophthalmology: Current Findings and Future Prospectives. BMJ Open Ophthalmol. 2019, 4, e000248. [Google Scholar] [CrossRef]
- Ander, S.E.; Diamond, M.S.; Coyne, C.B. Immune Responses at the Maternal-Fetal Interface. Sci. Immunol. 2019, 4, eaat6114. [Google Scholar] [CrossRef]
- Kniesel, U.; Wolburg, H. Tight Junctions of the Blood–Brain Barrier. Cell. Mol. Neurobiol. 2000, 20, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Mölzer, C.; Heissigerova, J.; Wilson, H.M.; Kuffova, L.; Forrester, J.V. Immune Privilege: The Microbiome and Uveitis. Front. Immunol. 2020, 11, 608377. [Google Scholar] [CrossRef]
- Al-Obaidi, M.M.J.; Desa, M.N.M. Mechanisms of Blood Brain Barrier Disruption by Different Types of Bacteria, and Bacterial–Host Interactions Facilitate the Bacterial Pathogen Invading the Brain. Cell. Mol. Neurobiol. 2018, 38, 1349–1368. [Google Scholar] [CrossRef] [PubMed]
- Kirstahler, P.; Bjerrum, S.S.; Friis-Møller, A.; la Cour, M.; Aarestrup, F.M.; Westh, H.; Pamp, S.J. Genomics-Based Identification of Microorganisms in Human Ocular Body Fluid. Sci. Rep. 2018, 8, 4126. [Google Scholar] [CrossRef]
- Mazoteras, P.; Quiles, M.G.; Martins Bispo, P.J.; Höfling-Lima, A.L.; Pignatari, A.C.; Casaroli-Marano, R.P. Analysis of Intraocular Lens Biofilms and Fluids After Long-Term Uncomplicated Cataract Surgery. Am. J. Ophthalmol. 2016, 169, 46–57. [Google Scholar] [CrossRef]
- Rosenbaum, J.T.; Asquith, M. The Microbiome and HLA-B27-Associated Acute Anterior Uveitis. Nat. Rev. Rheumatol. 2018, 14, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Vendomèle, J.; Khebizi, Q.; Fisson, S. Cellular and Molecular Mechanisms of Anterior Chamber-Associated Immune Deviation (ACAID): What We Have Learned from Knockout Mice. Front. Immunol. 2017, 8, 1686. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, B.; Li, W. Defining the Normal Core Microbiome of Conjunctival Microbial Communities. Clin. Microbiol. Infect. 2016, 22, 643.e7–643.e12. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Yang, B.; Huang, Y. Efficacy of 0.5% Levofloxacin and 5.0% Povidone-Iodine Eyedrops in Reducing Conjunctival Bacterial Flora: Metagenomic Analysis. J. Ophthalmol. 2020, 2020, 1780498. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kawamura, Y.; Fujiwara, N.; Naka, T.; Liu, H.; Huang, X.; Kobayashi, K.; Ezaki, T. Rothia Aeria Sp. Nov., Rhodococcus Baikonurensis Sp. Nov. and Arthrobacter Russicus Sp. Nov., Isolated from Air in the Russian Space Laboratory Mir. Int. J. Syst. Evol. Microbiol. 2004, 54, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Segata, N. No Bacteria Found in Healthy Placentas. Nature 2019, 572, 317–318. [Google Scholar] [CrossRef]
- Perlejewski, K.; Bukowska-Ośko, I.; Nakamura, S.; Motooka, D.; Stokowy, T.; Płoski, R.; Rydzanicz, M.; Zakrzewska-Pniewska, B.; Podlecka-Piętowska, A.; Nojszewska, M.; et al. Metagenomic Analysis of Cerebrospinal Fluid from Patients with Multiple Sclerosis. Adv. Exp. Med. Biol. 2016, 935, 89–98. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Age (years) | Sex | Ethnicity | Eye | |
|---|---|---|---|---|
| Patient 1 | 70 | Female | Caucasian | Left |
| Patient 2 | 73 | Male | Caucasian | Right |
| Patient 3 | 83 | Female | Caucasian | Right |
| Control 1 | 56 | Male | Caucasian | Right |
| Control 2 | 43 | Female | Caucasian | Right |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlunck, G.; Maier, P.; Maier, B.; Maier, W.; Strempel, S.; Reinhard, T.; Heinzelmann, S. Next-Generation Sequencing of the Human Aqueous Humour Microbiome. Int. J. Mol. Sci. 2024, 25, 6128. https://doi.org/10.3390/ijms25116128
Schlunck G, Maier P, Maier B, Maier W, Strempel S, Reinhard T, Heinzelmann S. Next-Generation Sequencing of the Human Aqueous Humour Microbiome. International Journal of Molecular Sciences. 2024; 25(11):6128. https://doi.org/10.3390/ijms25116128
Chicago/Turabian StyleSchlunck, Günther, Philip Maier, Barbara Maier, Wolfgang Maier, Sebastian Strempel, Thomas Reinhard, and Sonja Heinzelmann. 2024. "Next-Generation Sequencing of the Human Aqueous Humour Microbiome" International Journal of Molecular Sciences 25, no. 11: 6128. https://doi.org/10.3390/ijms25116128
APA StyleSchlunck, G., Maier, P., Maier, B., Maier, W., Strempel, S., Reinhard, T., & Heinzelmann, S. (2024). Next-Generation Sequencing of the Human Aqueous Humour Microbiome. International Journal of Molecular Sciences, 25(11), 6128. https://doi.org/10.3390/ijms25116128