


Zebrafish as a Human Muscle Model for Studying Age-Dependent Sarcopenia and Frailty

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Results
2.1. Morphometric and Motor Activity Analysis Revealed a Decline in Physical Condition and a Concurrent Increase in Frailty with Aging
2.2. The Structure and Organization of the Zebrafish Skeletal Muscle Are Significantly Compromised with Age
2.3. Pathways of Muscle Growth and Differentiation undergo Alterations with Aging in Zebrafish
2.4. Zebrafish Aging Revealed Alterations in the Ultrastructure of Muscle and Mitochondria
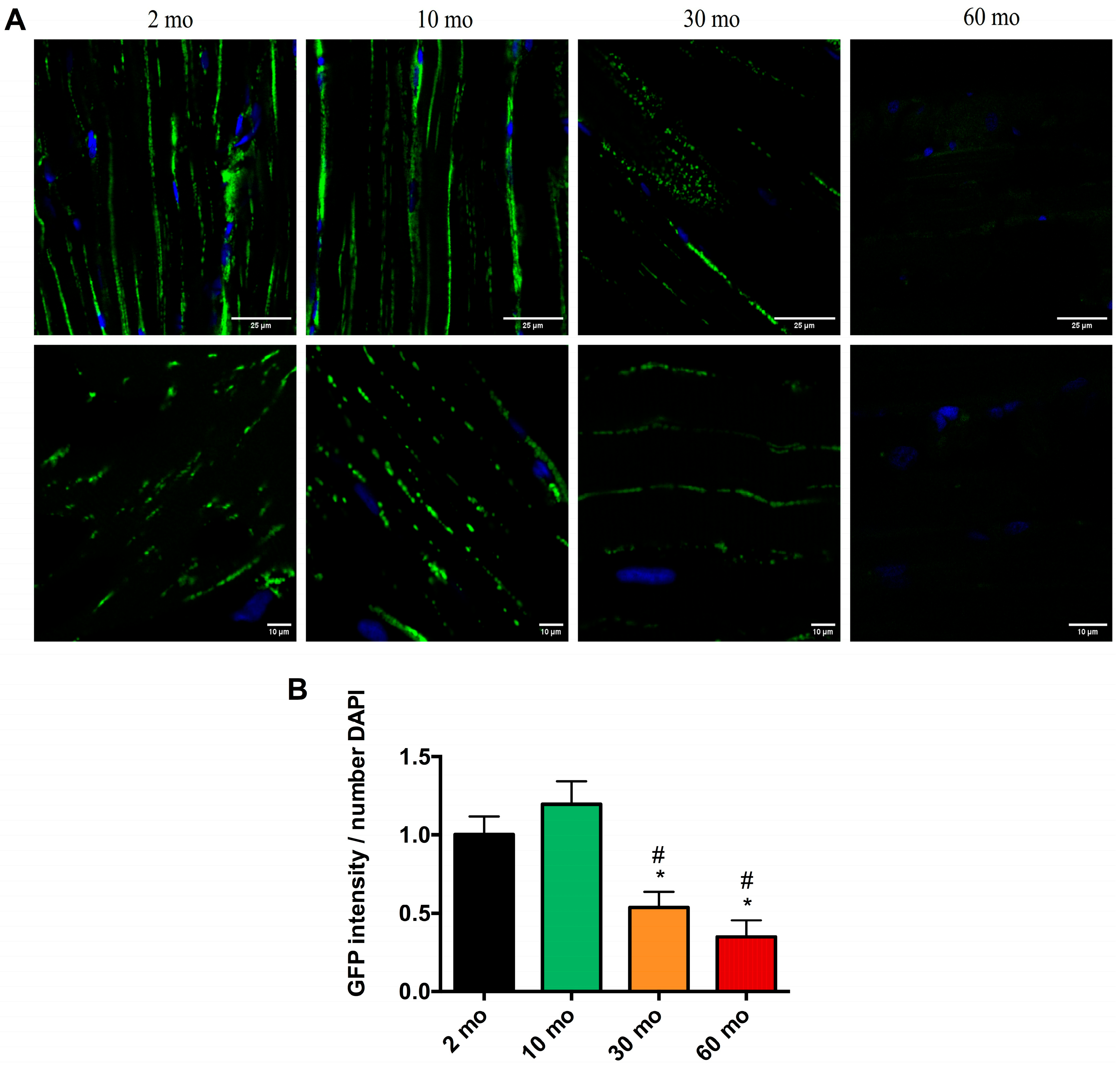
2.5. Confocal Analysis of Skeletal Muscle in Mito-GFP Zebrafish Shows Consistent Loss of Mitochondria with Age
3. Discussion
4. Materials and Methods
4.1. Fish Maintenance and Experimental Groups
4.2. Assessment of Locomotor Activity
4.3. Frailty Index Calculation
4.4. Gene Expression Analyses
4.5. Skeletal Muscle Histology and Morphometric Analysis
4.6. Transmission Electron Microscopy Analysis
4.7. Confocal Microscopy Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; El Assar, M.; Rodríguez-Mañas, L. Frailty and Sarcopenia as the Basis for the Phenotypic Manifestation of Chronic Diseases in Older Adults. Mol. Asp. Med. 2016, 50, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Rogeri, P.S.; Zanella, R.; Martins, G.L.; Garcia, M.D.A.; Leite, G.; Lugaresi, R.; Gasparini, S.O.; Sperandio, G.A.; Ferreira, L.H.B.; Souza-Junior, T.P.; et al. Strategies to Prevent Sarcopenia in the Aging Process: Role of Protein Intake and Exercise. Nutrients 2021, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Christian, C.J.; Benian, G.M. Animal Models of Sarcopenia. Aging Cell 2020, 19, e13223. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.K.A.; de Leonardis, E.C.; Guerrero-Martínez, J.A.; Rahim, I.; Mokhtar, D.M.; Saleh, A.M.; Abdalla, K.E.H.; Pozo, M.J.; Escames, G.; López, L.C.; et al. Identification of Morphological Markers of Sarcopenia at Early Stage of Aging in Skeletal Muscle of Mice. Exp. Gerontol. 2016, 83, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Rusanova, I.; Rahim, I.; Escames, G.; López, L.C.; Mokhtar, D.M.; Acuña-Castroviejo, D. The Protective Effect of Melatonin Against Age-Associated, Sarcopenia-Dependent Tubular Aggregate Formation, Lactate Depletion, and Mitochondrial Changes. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Aranda-Martínez, P.; Fernández-Martínez, J.; Guerra-Librero, A.; Escames, G.; López, L.C.; Alsaadawy, R.M.; Acuña-Castroviejo, D. Lack of NLRP3 Inflammasome Activation Reduces Age-Dependent Sarcopenia and Mitochondrial Dysfunction, Favoring the Prophylactic Effect of Melatonin. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Martínez, J.; Ramírez-Casas, Y.; Aranda-Martínez, P.; López-Rodríguez, A.; Sayed, R.K.A.; Escames, G.; Acuña-Castroviejo, D. IMS-Bmal1−/− Mice Show Evident Signs of Sarcopenia That Are Counteracted by Exercise and Melatonin Therapies. J. Pineal Res. 2023, 76, e12912. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.L.; Zhang, X.; McCarthy, J.J.; McDearmon, E.L.; Hornberger, T.A.; Russell, B.; Campbell, K.S.; Arbogast, S.; Reid, M.B.; Walker, J.R.; et al. CLOCK and BMAL1 Regulate MyoD and Are Necessary for Maintenance of Skeletal Muscle Phenotype and Function. Proc. Natl. Acad. Sci. USA 2010, 107, 19090–19095. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Hamlish, N.X.; Thakkar, A.V.; Steffeck, A.W.T.; Rendleman, E.J.; Khan, N.H.; Waldeck, N.J.; DeVilbiss, A.W.; Martin-Sandoval, M.S.; Mathews, T.P.; et al. BMAL1 Drives Muscle Repair through Control of Hypoxic NAD+ Regeneration in Satellite Cells. Genes Dev. 2022, 36, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Aranda-martínez, P.; Fernández-martínez, J.; Ramírez-casas, Y.; Guerra-librero, A.; Rodríguez-santana, C.; Escames, G.; Acuña-castroviejo, D. The Zebrafish, an Outstanding Model for Biomedical Research in the Field of Melatonin and Human Diseases. Int. J. Mol. Sci. 2022, 23, 7438. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, G.S.; Kauffman, E.J.; Wang, X.; Stewart, R.; Moore, J.L.; Kasales, C.J.; Demidenko, E.; Cheng, K.C. Life Spans and Senescent Phenotypes in Two Strains of Zebrafish (Danio Rerio). Exp. Gerontol. 2002, 37, 1055–1068. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M.; Elizondo, M.R.; Mills, M.G.; Gordon, T.N.; Engeszer, R.E. Normal table of postembryonic zebrafish development: Staging by externally visible anatomy of the living fish. Dev. Dyn. 2009, 238, 2975–3015. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, G.S. Comparative aspects of zebrafish (Danio rerio) as a model for aging research. Exp. Gerontol. 2003, 38, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S. The search for evolutionary developmental origins of aging in zebrafish: A novel intersection of developmental and senescence biology in the zebrafish model system. Birth Defects Res. C Embryo Today 2011, 93, 229–248. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, G.S.; Cheng, K.C. A call to fins! Zebrafish as a gerontological model. Aging Cell 2002, 1, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.C.; Yang, D.; Chen, Z.L.; Xiao, J.L.; Xiao, Q.; Li, C.L.; Zhou, Z.Q.; Peng, X.Y.; Tang, C.F.; Zheng, L. Exercise Intervention Mitigates Zebrafish Age-Related Sarcopenia via Alleviating Mitochondrial Dysfunction. FEBS J. 2023, 290, 1519–1530. [Google Scholar] [CrossRef]
- Rutkove, S.B.; Chen, Z.Z.; Pandeya, S.; Callegari, S.; Mourey, T.; Nagy, J.A.; Nath, A.K. Surface Electrical Impedance Myography Detects Skeletal Muscle Atrophy in Aged Wildtype Zebrafish and Aged gpr27 Knockout Zebrafish. Biomedicines 2023, 11, 1938. [Google Scholar] [CrossRef]
- Gilbert, M.J.; Zerulla, T.C.; Tierney, K.B. Zebrafish (Danio rerio) as a model for the study of aging and exercise: Physical ability and trainability decrease with age. Exp. Gerontol. 2014, 50, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Daya, A.; Donaka, R.; Karasik, D. Zebrafish Models of Sarcopenia. Dis. Models Mech. 2020, 13, dmm042689. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.L.; Guo, C.; Zou, Y.Y.; Feng, C.; Yang, D.X.; Sun, C.C.; Wen, W.; Jian, Z.J.; Zhao, Z.; Xiao, Q.; et al. Aerobic Exercise Enhances Mitochondrial Homeostasis to Counteract D-Galactose-Induced Sarcopenia in Zebrafish. Exp. Gerontol. 2023, 180, 112265. [Google Scholar] [CrossRef] [PubMed]
- Martinez De Toda, I.; Garrido, A.; Vida, C.; Gomez-Cabrera, M.C.; Viña, J.; De La Fuente, M. Frailty Quantified by the “Valencia Score” as a Potential Predictor of Lifespan in Mice. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic Signaling Deficits Underlie Amino Acid Resistance of Wasting, Aging Muscle. FASEB J. 2005, 19, 1–22. [Google Scholar] [CrossRef]
- Paturi, S.; Gutta, A.K.; Katta, A.; Kakarla, S.K.; Arvapalli, R.K.; Gadde, M.K.; Nalabotu, S.K.; Rice, K.M.; Wu, M.; Blough, E. Effects of Aging and Gender on Muscle Mass and Regulation of Akt-MTOR-P70s6k Related Signaling in the F344BN Rat Model. Mech. Ageing Dev. 2010, 131, 202–209. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Ganassi, M.; Badodi, S.; Wanders, K.; Zammit, P.S. Myogenin Is an Essential Regulator of Adult Myofibre Growth and Muscle Stem Cell Homeostasis. Elife 2020, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.E.; Ingham, P.W. Control of Muscle Fibre-Type Diversity during Embryonic Development: The Zebrafish Paradigm. Mech. Dev. 2013, 130, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.E.; Ono, Y.; Wang, X.; Elworthy, S.; Cunliffe, V.T.; Ingham, P.W. The Role of Sox6 in Zebrafish Muscle Fiber Type Specification. Skelet Muscle 2015, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Short, K.R.; Bigelow, M.L.; Kahl, J.; Singh, R.; Coenen-Schimke, J.; Raghavakaimal, S.; Nair, K.S. Decline in skeletal muscle mitochondrial function with aging in humans. Proc. Natl. Acad. Sci. USA 2005, 102, 5618–5623. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhao, H.; Li, Y. Mitochondrial dynamics in health and disease: Mechanisms and potential targets. Signal Transduct Target. Ther. 2023, 8, 333. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Slack, B.E.; Uchiyama, J.; Zhdanova, I.V. Zebrafish as a Genetic Model in Biological and Behavioral Gerontology: Where Development Meets Aging in Vertebrates-A Mini-Review. Gerontology 2009, 55, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.B. Weight and Body Mass Index in Old Age: Do They Still Matter? J. Am. Geriatr. Soc. 2017, 6, 1898–1899. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.S.; Leurgans, S.E.; Wang, T.; Schnaider-Beeri, M.; Agarwal, P.; Dawe, R.J.; Delbono, O.; Bennett, D.A. Motor function is the primary driver of the associations of sarcopenia and physical frailty with adverse health outcomes in community-dwelling older adults. PLoS. ONE 2021, 16, e0245680. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.; Kwon, E.-S.; Kwon, K.-S. Molecular Mechanisms and Therapeutic Interventions in Sarcopenia. Osteoporos. Sarcopenia 2017, 3, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Zheng, K.; Li, W.; An, K.; Liu, Y.; Xiao, X.; Hai, S.; Dong, B.; Li, S.; An, Z.; et al. Post-Translational Regulation of Muscle Growth, Muscle Aging and Sarcopenia. J. Cachexia Sarcopenia Muscle 2023, 14, 1212–1227. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- Bano, G.; Trevisan, C.; Carraro, S.; Solmi, M.; Luchini, C.; Stubbs, B.; Manzato, E.; Sergi, G.; Veronese, N. Inflammation and Sarcopenia: A Systematic Review and Meta-Analysis. Maturitas 2017, 96, 10–15. [Google Scholar] [CrossRef]
- Pan, L.; Xie, W.; Fu, X.; Lu, W.; Jin, H.; Lai, J.; Zhang, A.; Yu, Y.; Li, Y.; Xiao, W. Inflammation and Sarcopenia: A Focus on Circulating Inflammatory Cytokines. Exp. Gerontol. 2021, 154, 111544. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhang, T.; Liu, H.; Li, Z.; Peng, L.; Wang, C.; Wang, T. Inflammaging: The Ground for Sarcopenia? Exp. Gerontol. 2022, 168, 111931. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Diaz, M.; Comai, G.; Di Girolamo, D.; Langa, F.; Tajbakhsh, S. Dynamics of Myogenic Differentiation Using a Novel Myogenin Knock-in Reporter Mouse. Skelet. Muscle 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Xu, C.; Wu, G.; Zohar, Y.; Du, S.J. Analysis of myostatin gene structure, expression and function in zebrafish. J. Exp. Biol. 2003, 206 Pt 22, 4067–4079. [Google Scholar] [CrossRef] [PubMed]
- Purves-Smith, F.M.; Sgarioto, N.; Hepple, R.T. Fiber Typing in Aging Muscle. Exerc. Sport Sci. Rev. 2014, 42, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Von Hofsten, J.; Elworthy, S.; Gilchrist, M.J.; Smith, J.C.; Wardle, F.C.; Ingham, P.W. Prdm1- and Sox6-Mediated Transcriptional Repression Specifies Muscle Fibre Type in the Zebrafish Embryo. EMBO. Rep. 2008, 9, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, F.; Buglio, A.L.; Vendemiale, G. Mitochondrial Impairment in Sarcopenia. Biology 2021, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Castroviejo, D. Melatonin Role in the Mitochondrial Function. Front. Biosci. 2007, 12, 947. [Google Scholar] [CrossRef] [PubMed]
- Ichii, S.; Matsuoka, I.; Okazaki, F.; Shimada, Y. Zebrafish Models for Skeletal Muscle Senescence: Lessons from Cell Cultures and Rodent Models. Molecules 2022, 27, 8625. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kang, K.H.; Kim, C.H.; Choi, S.Y. Real-Time Imaging of Mitochondria in Transgenic Zebrafish Expressing Mitochondrially Targeted GFP. Biotechniques 2008, 45, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book, A Guide for the Laboratory Use of Zebrafish (Danio Rerio); Univ Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| mfn1 | AACGAAGTGTGCTCTGCTCA | GGATTCAGAGTTCGCCACCA |
| opa1 | AGACTGGAAGCAGAGGTGGA | GGAAGTGACGTCGAAAGAGC |
| drp1 | AACATCCAGGACAGCGTACC | TCACCACAAGTGCGTCTCTC |
| dyn2 | CGCAGATAGCAGTTGTCGGA | TCTGCTTCAATCTCCTGCCG |
| akt | TGCTGAAGAGTGACGGTACG | CTTTCTTCAGGCGTCTCCAC |
| p70s6k | CAGACTCCCGTTGACAGTCC | ATTGGACTGAGAGGCGTTCG |
| myogenin | CTCCACATACTGGGGTGTCG | GTCGTTCAGCAGATCCTCGT |
| prdm1a | TTGAACGCTTTGACATCAGC | GCTGCGATGAACTTTGATGA |
| gapdh | TCACACCAAGTGTCAGGACG | CGCCTTCTGCCTTAACCTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aranda-Martínez, P.; Sayed, R.K.A.; Fernández-Martínez, J.; Ramírez-Casas, Y.; Yang, Y.; Escames, G.; Acuña-Castroviejo, D. Zebrafish as a Human Muscle Model for Studying Age-Dependent Sarcopenia and Frailty. Int. J. Mol. Sci. 2024, 25, 6166. https://doi.org/10.3390/ijms25116166
Aranda-Martínez P, Sayed RKA, Fernández-Martínez J, Ramírez-Casas Y, Yang Y, Escames G, Acuña-Castroviejo D. Zebrafish as a Human Muscle Model for Studying Age-Dependent Sarcopenia and Frailty. International Journal of Molecular Sciences. 2024; 25(11):6166. https://doi.org/10.3390/ijms25116166
Chicago/Turabian StyleAranda-Martínez, Paula, Ramy K. A. Sayed, José Fernández-Martínez, Yolanda Ramírez-Casas, Yang Yang, Germaine Escames, and Darío Acuña-Castroviejo. 2024. "Zebrafish as a Human Muscle Model for Studying Age-Dependent Sarcopenia and Frailty" International Journal of Molecular Sciences 25, no. 11: 6166. https://doi.org/10.3390/ijms25116166
APA StyleAranda-Martínez, P., Sayed, R. K. A., Fernández-Martínez, J., Ramírez-Casas, Y., Yang, Y., Escames, G., & Acuña-Castroviejo, D. (2024). Zebrafish as a Human Muscle Model for Studying Age-Dependent Sarcopenia and Frailty. International Journal of Molecular Sciences, 25(11), 6166. https://doi.org/10.3390/ijms25116166