
Human Brain In Vitro Model for Pathogen Infection-Related Neurodegeneration Study


Abstract
1. Introduction
2. Cellular Population of Central Nervous System
2.1. Subtype Neurons
2.2. Astrocyte
2.3. Oligodendrocyte
2.4. Microglia
2.5. Brain Microvascular Endothelial Cells and Stromal Cells
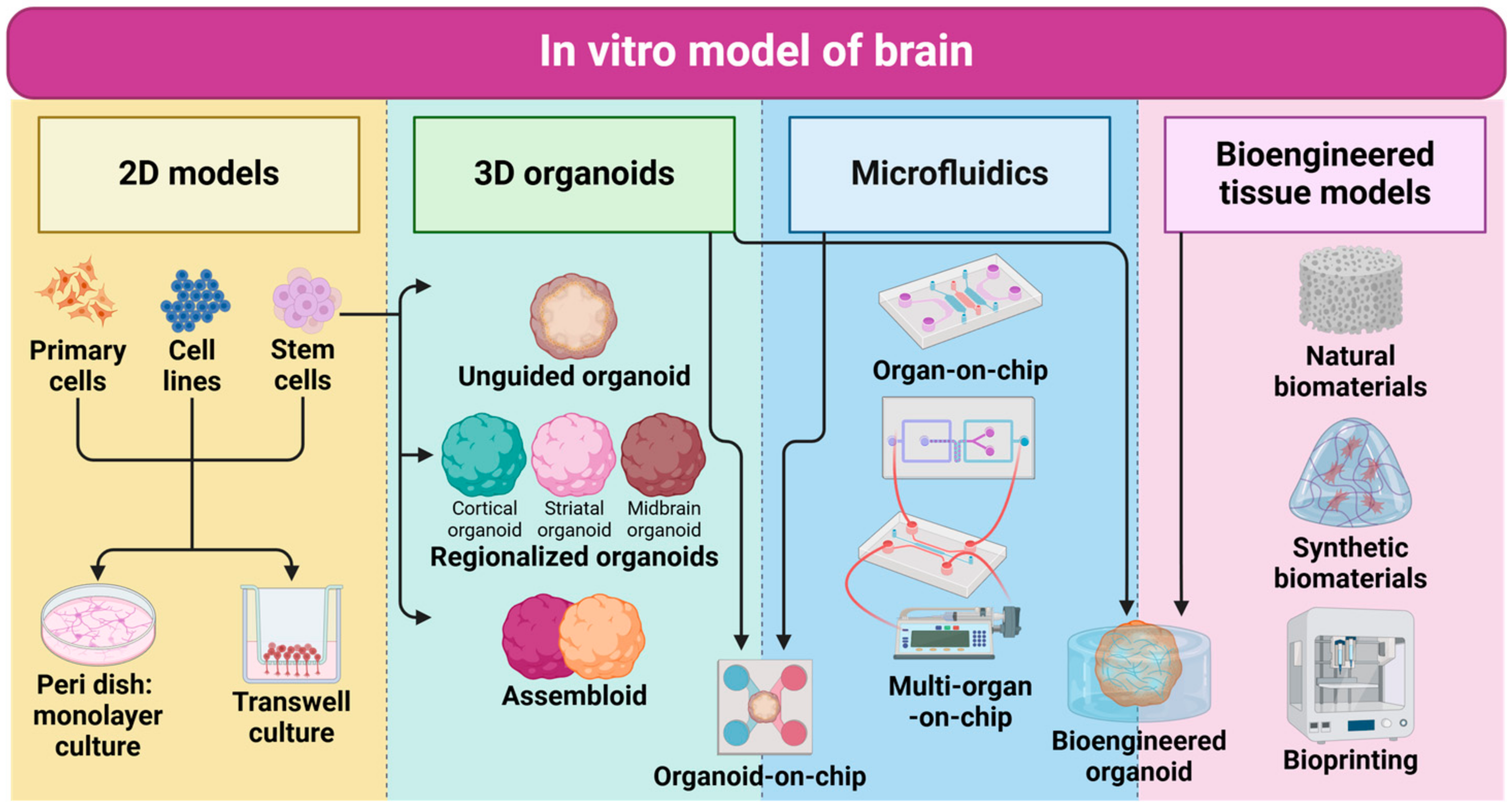
3. In Vitro Model of Human Brain
3.1. 2D Monolayer Cultures
3.2. 3D Organoids
3.2.1. Unguided (Cerebral Organoid)
3.2.2. Regionalized (Cortical Organoids, Assembloids)
3.2.3. Bioengineered
3.3. Organ-on-Chip
3.3.1. Blood–Brain Barrier
3.3.2. Neuroinflammation
3.3.3. Multi-Organ Crosstalk
3.4. Bioengineered Tissue Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Vitro Model | Advantages | Disadvantages | Citations | |
|---|---|---|---|---|
| 2D Monolayer Cultures | Primary Cells | Properties of native brain tissues. | Challenges of accessibility, limited proliferation, inconsistent results, and lower growth rates. | Danz et al. [94] |
| Immortal Cell Lines | Indefinite proliferation, robust, and easy to manipulate. | Alter physiological properties due to genetic modification. | He et al. [97], Chua et al. [98], Zhang et al. [99], Slanzi et al. [100] | |
| Stem Cells (NSCs, hPSCs) | Differentiation into various specialized cell types, personalized genetic information (hiPSCs). | Complicated cultural conditions, and potential variability in differentiation outcomes. | Pietilainen et al. [110], Drager et al. [111], Stebbins et al. [112] | |
| 3D Organoids | Unguided Brain Organoids | Spontaneous formation of diverse brain regions, suitable model for studying brain development. | Variability between samples, lack of external guidance. | Lancaster et al. [116], Giandomenico et al. [122], Pellegrini et al. [123], Bagley et al. [124] |
| Regionalized Organoids | Specific growth factors combined for region-specific differentiation, form functional synapses, and model disease-specific neural activity. | Skillset of precise control of differentiation protocols. | Pasca et al. [128], Jo et al. [130] | |
| Assembloids | Recapitulation of interactions and connectivity between various brain regions, involving multiple brain regions. | Batch variation between samples, inconsistent producibility, and unstable long-term cultures. | Birey et al. [115], Sloan et al. [114], Miura et al. [134] | |
| Bioengineered Organoids | Improved 3D structural support, and complex microenvironment modeling. | Complex to develop, limited nutrient and waste exchange. | Lancaster et al. [137], Cho et al. [138], Cakir et al. [139] | |
| Brain-on-a-Chip (BoC) | BBB-on-a-Chip | Functional BBB microenvironment, useful for drug testing. | Limitations in replicating the entire BBB functionality. | Griep et al. [146], Yu et al. [147] |
| Neuroinflammation Models | Residual and peripheral inflammatory responses. | Requires precise manipulation, variability due to complex mixture of different cells. | Pediaditakis et al. [149], Herland et al. [151] | |
| Multi-Organ Chip Models | Mimics organ crosstalk including gut–brain and lung–brain interactions. | Complex and high-cost to fabricate, limited ability to fully emulate the systemic interactions. | Kim et al. [154], Wang et al. [7] | |
| Bioengineered Tissue Models | Biomaterials | Enhanced neurogenesis and neural differentiation, variable choice of materials, reduced cell death, and vascularization in brain models. | Potential biocompatibility issues, variation due to manipulation of biomaterials. | Shin et al. [161] |
| Bioprinting | Precise spatial control, improved integration of multiple cell types and materials, allows for high reproducibility and scalability. | Technical complexity, high cost of equipment and materials, potential issues with cell viability and functionality post-printing, and requires specialized knowledge and skills for operation. | Roth et. al. [162], Park et al. [163], Jin et al. [164], Grebenyuk et al. [165] | |
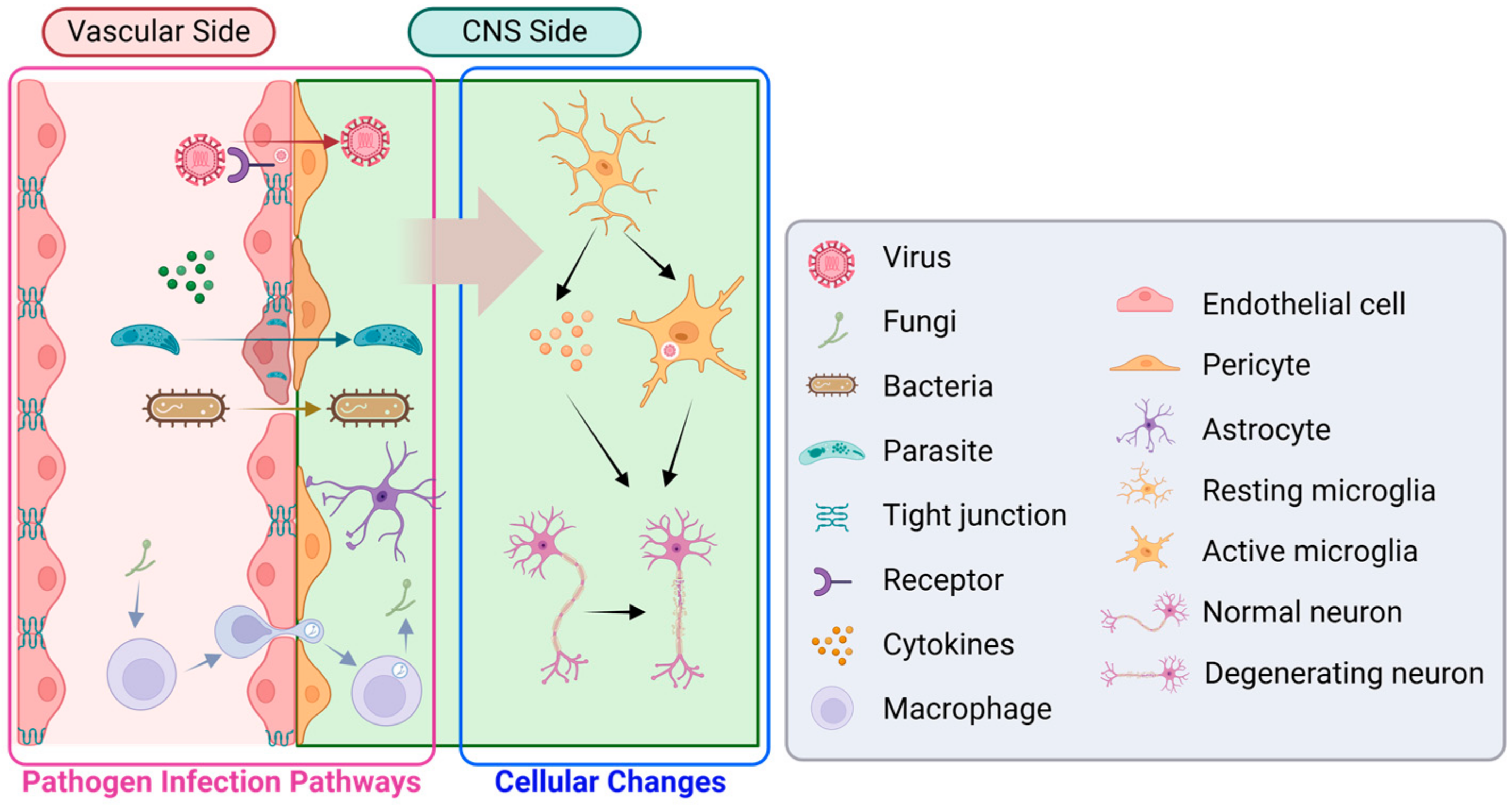
4. Application of Infection Model for Neurodegenerative Diseases and Neurological Disorder
4.1. Virus
4.1.1. ZIKV
4.1.2. SARS-CoV-2
4.1.3. Other Viruses
4.2. Fungi
4.3. Bacteria
4.4. Parasite
| Pathogens | Cell Resources | In Vitro Models | Insights | Citations | |
|---|---|---|---|---|---|
| Virus | Zika Virus (ZIKV) | Neural progenitor cells, mature neurons, BMECs | Region-specific brain organoids, hiPSC-derived brain organoids, 2D transwell system | ZIKV infection leads to microcephaly-like symptoms, disrupts neurogenesis, and can cross the BBB without compromising the integrity of tight junctions. Enoxacin treatment prevents ZIKV-associated damage [117,170,171]. | Xu et al. [117], Garcez et al. [170], Alimonti et al. [171]. |
| SARS-CoV-2 | Astrocytes, microglia, neurons, BMECs | 2D static and 3D microfluidic BBB models, lung–brain MPS, brain organoids, hiPSC-derived monolayer neural cells, choroid plexus organoids, pericyte-containing cortical organoids | SARS-CoV-2 affects brain function through direct infection and secondary effects. It primarily affects vascular and immune cells, leading to neurological symptoms through inflammatory processes. The spike protein disrupts BBB integrity [176,177,178,179]. | Buzhdygan et al. [168], Wang et al. [7], Ramani et al. [176], Jacob et al. [177], Martínez-Mármo et al. [178], Aguado et al. [179], Wang et al. [181]. | |
| Tick-borne encephalitis virus (TBEV) | Neurons, astrocytes | 2D co-culture model | TBEV leads to neuronal death and neurite loss, with astrocytes showing limited infection but exhibiting astrogliosis to protect neurons [182]. | Fares et al. [182]. | |
| Human immunodeficiency virus (HIV) | Neurons, microglia, astrocytes | 3D human brain organoid | HIV infection leads to increased inflammatory response, neuronal loss, astrocytosis, and compromised synaptic integrity [183]. | Reis et al. [183]. | |
| Influenza virus (H1N1-WSN strain) | Neurons, NSCs, astrocytes | hiPSC-derived brain organoids in a spinning bioreactor | The virus causes widespread infection, apoptosis in neurons and NSCs, and reduced organoid size [184]. | Zhang et al. [184]. | |
| Herpes simplex virus type 1 (HSV-1) | Neurons, glial cells | 3D bioengineered human brain model, scaffold-free 3D human neuronal cultures | HSV-1 infection shows amyloid plaque-like structures, gliosis, and neuroinflammation, and is effectively inhibited by acyclovir [185,187]. | Cairns et al. [185], D’Aiuto et al. [187]. | |
| John Cunningham virus (JCV) | Oligodendrocytes, astrocytes, neurons | iPSC-derived 3D brain organoid | JCV infects oligodendrocytes and astrocytes, not neurons, leading to PML [186]. | Barreras et al. [186]. | |
| Fungi | Cryptococcus neoformans | Murine BV-2 cell lines, microglia, NSCs, BMECs, brain vascular pericytes | 2D static model, human-neurovascular-unit-on-a-chip (hNVU chip) | The fungus causes microglial activation, and phagocytosis, and crosses the BBB without disrupting tight junctions [188,189]. | Koutsouras et al. [188], Kim et al. [189]. |
| Bacteria | Staphylococcus aureus (SA) | BMECs | 2D static BBB model | SA infection increases paracellular permeability and cytokine release, reducing the expression of interendothelial junction proteins [192]. | McLoughlin et al. [192]. |
| Neisseria meningitidis | BMECs, leptomeningeal cells | iPSC-derived BMECs co-cultured with leptomeningeal cells on transwell system | The bacteria adhere to BMECs, cross the barrier transcellularly, and compromise barrier integrity over time [193]. | Endres et al. [193]. | |
| Parasite | Toxoplasma gondii (T. gondii) | Neurons, astrocytes, oligodendrocytes | Human cerebral organoids, 3D brain spheroids | T. gondii infects various brain cells, causing neural cell death, size reduction in organoids, and cellular damage [197]. | Seo et al. [196], Leite et al. [197]. |
| Naegleria fowleri | Rat BMECs | 2D static BBB model | The amoeba decreases TEER, degrades tight junction proteins, and disrupts the BBB, leading to potential CNS inflammation [198]. | Coronado-Velazquez et al. [198]. | |
5. Conclusions and Future Directions
5.1. Drug Development
5.2. Gene Therapy
5.3. Vaccine
Funding
Conflicts of Interest
References
- Lynn, C.W.; Bassett, D.S. The physics of brain network structure, function and control. Nat. Rev. Phys. 2019, 1, 318–332. [Google Scholar] [CrossRef]
- Winnubst, J.; Arber, S. A census of cell types in the brain’s motor cortex. Nature 2021, 598, 33–34. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Song, H.; Ming, G.L. Genetics of human brain development. Nat. Rev. Genet. 2024, 25, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Kim, P. Biomimetic Strategies for the Glioblastoma Microenvironment. Front. Mater. 2017, 4, 45. [Google Scholar] [CrossRef]
- Wichmann, T.O.; Damkier, H.H.; Pedersen, M. A Brief Overview of the Cerebrospinal Fluid System and Its Implications for Brain and Spinal Cord Diseases. Front. Hum. Neurosci. 2022, 15, 737217. [Google Scholar] [CrossRef] [PubMed]
- Molnár, Z.; Clowry, G.J.; Šestan, N.; Alzu’Bi, A.; Bakken, T.; Hevner, R.F.; Hüppi, P.S.; Kostović, I.; Rakic, P.; Anton, E.S.; et al. New insights into the development of the human cerebral cortex. J. Anat. 2019, 235, 432–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jin, L.; Zhang, M.; Wu, Y.; Duan, Z.; Guo, Y.; Wang, C.; Guo, Y.; Chen, W.; Liao, Z.; et al. Blood–brain barrier injury and neuroinflammation induced by SARS-CoV-2 in a lung–brain microphysiological system. Nat. Biomed. Eng. 2023, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dixon, T.A.; Muotri, A.R. Advancing preclinical models of psychiatric disorders with human brain organoid cultures. Mol. Psychiatry 2022, 28, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Loewa, A.; Feng, J.J.; Hedtrich, S. Human disease models in drug development. Nat. Rev. Bioeng. 2023, 1, 545–559. [Google Scholar] [CrossRef] [PubMed]
- Eichmüller, O.L.; Knoblich, J.A. Human cerebral organoids—A new tool for clinical neurology research. Nat. Rev. Neurol. 2022, 18, 661–680. [Google Scholar] [CrossRef]
- Siletti, K.; Hodge, R.D.; Alejandro Mossi, A.; Ka Wai, L.; Ding, S.L.; Hu, L.; Lönnerberg, P.; Bakken, T.E.; Casper, T.; Clark, M.; et al. Transcriptomic diversity of cell types across the adult human brain. Science 2023, 382, 7046. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. The Human Brain in numbers: A Linearly scaled-up Primate Brain. Front. Hum. Neurosci. 2009, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Herculano-Houzel, S. The remarkable, yet not extraordinary, human brain as a scaled-up primate brain and its associated cost. Proc. Natl. Acad. Sci. USA 2012, 109 (Suppl. S1), 10661–10668. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, F.A.C.; Carvalho, L.R.B.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.L.; Leite, R.E.P.; Filho, W.J.; Lent, R.; Herculano-Houzel, S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C. Towards an Understanding of Synapse Formation. Neuron 2018, 100, 276–293. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.N.; Allen, D.E.; Keefe, M.G.; Mancia Leon, W.R.; Ziffra, R.S.; Crouch, E.E.; Alvarez-Buylla, A.; Nowakowski, T.J. Individual human cortical progenitors can produce excitatory and inhibitory neurons. Nature 2022, 601, 397–403. [Google Scholar] [CrossRef]
- He, H.-y.; Cline, H.T. What Is Excitation/Inhibition and How Is It Regulated? A Case of the Elephant and the Wisemen. J. Exp. Neurosci. 2019, 13, 117906951985937. [Google Scholar] [CrossRef] [PubMed]
- Bruining, H.; Hardstone, R.; Juarez-Martinez, E.L.; Sprengers, J.; Avramiea, A.-E.; Simpraga, S.; Houtman, S.J.; Poil, S.-S.; Dallares, E.; Palva, S.; et al. Measurement of excitation-inhibition ratio in autism spectrum disorder using critical brain dynamics. Sci. Rep. 2020, 10, 9195. [Google Scholar] [CrossRef] [PubMed]
- Wester, J.C.; Mahadevan, V.; Rhodes, C.T.; Calvigioni, D.; Venkatesh, S.; Maric, D.; Hunt, S.; Yuan, X.; Zhang, Y.; Petros, T.J.; et al. Neocortical Projection Neurons Instruct Inhibitory Interneuron Circuit Development in a Lineage-Dependent Manner. Neuron 2019, 102, 960–975.e6. [Google Scholar] [CrossRef] [PubMed]
- Musall, S.; Sun, X.R.; Mohan, H.; An, X.; Gluf, S.; Li, S.-J.; Drewes, R.; Cravo, E.; Lenzi, I.; Yin, C.; et al. Pyramidal cell types drive functionally distinct cortical activity patterns during decision-making. Nat. Neurosci. 2023, 26, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Penzes, P. Common Mechanisms of Excitatory and Inhibitory Imbalance in Schizophrenia and Autism Spectrum Disorders. Curr. Mol. Med. 2015, 15, 146–167. [Google Scholar] [CrossRef] [PubMed]
- Wonders, C.P.; Anderson, S.A. The origin and specification of cortical interneurons. Nat. Rev. Neurosci. 2006, 7, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Cao, Q.; Moser, M.-B.; Moser, E.I. Parvalbumin and Somatostatin Interneurons Control Different Space-Coding Networks in the Medial Entorhinal Cortex. Cell 2017, 171, 507–521.e17. [Google Scholar] [CrossRef] [PubMed]
- Defelipe, J.; González-Albo, M.C.; Del Río, M.R.; Elston, G.N. Distribution and patterns of connectivity of interneurons containing calbindin, calretinin, and parvalbumin in visual areas of the occipital and temporal lobes of the macaque monkey. J. Comp. Neurol. 1999, 412, 515–526. [Google Scholar] [CrossRef]
- Georgiou, C.; Kehayas, V.; Lee, K.S.; Brandalise, F.; Sahlender, D.A.; Blanc, J.; Knott, G.; Holtmaat, A. A subpopulation of cortical VIP-expressing interneurons with highly dynamic spines. Commun. Biol. 2022, 5, 352. [Google Scholar] [CrossRef] [PubMed]
- Llorca, A.; Deogracias, R. Origin, Development, and Synaptogenesis of Cortical Interneurons. Front. Neurosci. 2022, 16, 929469. [Google Scholar] [CrossRef] [PubMed]
- Udakis, M.; Pedrosa, V.; Chamberlain, S.E.L.; Clopath, C.; Mellor, J.R. Interneuron-specific plasticity at parvalbumin and somatostatin inhibitory synapses onto CA1 pyramidal neurons shapes hippocampal output. Nat. Commun. 2020, 11, 4395. [Google Scholar] [CrossRef] [PubMed]
- Urban-Ciecko, J.; Barth, A.L. Somatostatin-expressing neurons in cortical networks. Nat. Rev. Neurosci. 2016, 17, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Chung, H.; Rowland, J.M.; Richards, B.A.; Kohl, M.M.; Kwag, J. Distinct roles of parvalbumin and somatostatin interneurons in gating the synchronization of spike times in the neocortex. Sci. Adv. 2020, 6, eaay5333. [Google Scholar] [CrossRef] [PubMed]
- Romero-Sosa, J.L.; Motanis, H.; Buonomano, D.V. Differential Excitability of PV and SST Neurons Results in Distinct Functional Roles in Inhibition Stabilization of Up States. J. Neurosci. 2021, 41, 7182–7196. [Google Scholar] [CrossRef] [PubMed]
- Hattori, R.; Kuchibhotla, K.V.; Froemke, R.C.; Komiyama, T. Functions and dysfunctions of neocortical inhibitory neuron subtypes. Nat. Neurosci. 2017, 20, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.A. Inhibitory neurons in human cortical circuits: Substrate for cognitive dysfunction in schizophrenia. Curr. Opin. Neurobiol. 2014, 26, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2009, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Fusco, R.; Cuzzocrea, S. Astrocytes: Role and Functions in Brain Pathologies. Front. Pharmacol. 2019, 10, 1114. [Google Scholar] [CrossRef] [PubMed]
- Bellot-Saez, A.; Kekesi, O.; Morley, J.W.; Buskila, Y. Astrocytic modulation of neuronal excitability through K+ spatial buffering. Neurosci. Biobehav. Rev. 2017, 77, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes Maintain Glutamate Homeostasis in the CNS by Controlling the Balance between Glutamate Uptake and Release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Toth, A.E.; Klepe, A.; Lipka, D.V.; Goldeman, C.; Brodin, B.; Nielsen, M.S. SorLA in astrocytes regulates blood-brain barrier integrity. Front. Drug Deliv. 2023, 2, 1082689. [Google Scholar] [CrossRef]
- Li, C.; Zhao, R.; Gao, K.; Wei, Z.; Yaoyao Yin, M.; Ting Lau, L.; Chui, D.; Cheung Hoi Yu, A. Astrocytes: Implications for Neuroinflammatory Pathogenesis of Alzheimers Disease. Curr. Alzheimer Res. 2011, 8, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Alexei, V.; Butt, A.M.; Li, B.; Illéš, P.; Zorec, R.; Alexey, S.; Tang, Y.; Sofroniew, M.V. Astrocytes in human central nervous system diseases: A frontier for new therapies. Signal Transduct. Target. Ther. 2023, 8, 396. [Google Scholar]
- Eugenin, E.A.; Clements, J.E.; Zink, M.C.; Berman, J.W. Human immunodeficiency virus infection of human astrocytes disrupts blood-brain barrier integrity by a gap junction-dependent mechanism. J. Neurosci. 2011, 31, 9456–9465. [Google Scholar] [CrossRef] [PubMed]
- Bohmwald, K.; Soto, J.A.; Andrade-Parra, C.; Fernandez-Fierro, A.; Espinoza, J.A.; Rios, M.; Eugenin, E.A.; Gonzalez, P.A.; Opazo, M.C.; Riedel, C.A.; et al. Lung pathology due to hRSV infection impairs blood-brain barrier permeability enabling astrocyte infection and a long-lasting inflammation in the CNS. Brain. Behav. Immun. 2021, 91, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.-P.; Kothary, R. Oligodendrocytes in a Nutshell. Front. Cell. Neurosci. 2015, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef] [PubMed]
- Munyeshyaka, M.; Fields, R.D. Oligodendroglia are emerging players in several forms of learning and memory. Commun. Biol. 2022, 5, 1148. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Nave, K.-A. Oligodendrocytes: Myelination and Axonal Support. Cold Spring Harb. Perspect. Biol. 2019, 8, a020479. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.S.; Zdunek, S.; Bergmann, O.; Bernard, S.; Salehpour, M.; Alkass, K.; Perl, S.; Tisdale, J.; Possnert, G.; Brundin, L.; et al. Dynamics of Oligodendrocyte Generation and Myelination in the Human Brain. Cell 2014, 159, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.S.Y.; Djelloul, M.; Steiner, E.; Bernard, S.; Salehpour, M.; Possnert, G.; Brundin, L.; Frisén, J. Dynamics of oligodendrocyte generation in multiple sclerosis. Nature 2019, 566, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.C.; Mierzwa, A.J.; Sullivan, G.M.; Sanchez, M.A. Myelin and oligodendrocyte lineage cells in white matter pathology and plasticity after traumatic brain injury. Neuropharmacology 2016, 110, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Yuen, T.J.; Silbereis, J.C.; Griveau, A.; Chang, S.M.; Daneman, R.; Fancy, S.P.; Zahed, H.; Maltepe, E.; Rowitch, D.H. Oligodendrocyte-Encoded HIF Function Couples Postnatal Myelination and White Matter Angiogenesis. Cell 2014, 158, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Nasrabady, S.E.; Rizvi, B.; Goldman, J.E.; Brickman, A.M. White matter changes in Alzheimer’s disease: A focus on myelin and oligodendrocytes. Acta Neuropathol. Commun. 2018, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Zhang, Q.; Anthony, S.M.; Zhou, Y.; Zou, X.; Cassell, M.; Perlman, S. Oligodendrocytes that survive acute coronavirus infection induce prolonged inflammatory responses in the CNS. Proc. Natl. Acad. Sci. USA 2020, 117, 15902–15910. [Google Scholar] [CrossRef]
- Schultz, V.; Barrie, J.A.; Donald, C.L.; Crawford, C.L.; Mullin, M.; Anderson, T.J.; Solomon, T.; Barnett, S.C.; Linington, C.; Kohl, A.; et al. Oligodendrocytes are susceptible to Zika virus infection in a mouse model of perinatal exposure: Implications for CNS complications. Glia 2021, 69, 2023–2036. [Google Scholar] [CrossRef]
- Peferoen, L.A.N.; Vogel, D.Y.S.; Ummenthum, K.; Breur, M.; Heijnen, P.D.A.M.; Gerritsen, W.H.; Peferoen-Baert, R.M.B.; Van Der Valk, P.; Dijkstra, C.D.; Amor, S. Activation Status of Human Microglia Is Dependent on Lesion Formation Stage and Remyelination in Multiple Sclerosis. J. Neuropathol. Exp. Neurol. 2015, 74, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Tarnawsky, S.P.; Yoder, M.C. From embryo mutation to adult degeneration. Nature 2017, 549, 340–342. [Google Scholar] [CrossRef] [PubMed]
- Hickman, S.; Izzy, S.; Sen, P.; Morsett, L.; El Khoury, J. Microglia in neurodegeneration. Nat. Neurosci. 2018, 21, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Félix, D.-G.; Bitarafan, S.; Lacroix, S.; Gosselin, D. Roles and regulation of microglia activity in multiple sclerosis: Insights from animal models. Nat. Rev. Neurosci. 2023, 24, 397–415. [Google Scholar]
- Kent, S.; Miron, V.E. Microglia regulation of central nervous system myelin health and regeneration. Nat. Rev. Immunol. 2023, 24, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Priller, J. Microglia and brain macrophages in the molecular age: From origin to neuropsychiatric disease. Nat. Rev. Neurosci. 2014, 15, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.C.; Partridge, A.T.; Sell, C.; Torres, C.; Martin-Garcia, J. Fate of microglia during HIV-1 infection: From activation to senescence? Glia 2017, 65, 431–446. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.U.; Lyu, J.; Kim, K.-D.; Chung, Y.C.; Yoon, G.Y.; Lee, S.; Hwang, I.; Shin, W.-H.; Ko, J.; Lee, J.-Y.; et al. SARS-CoV-2 Infection of Microglia Elicits Proinflammatory Activation and Apoptotic Cell Death. Microbiol. Spectr. 2022, 10, 01091-22. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, G.L.; Ballard, V.A.; Glausen, T.; Boone, Z.; Teamer, J.; Hinkson, C.L.; Wohlfert, E.A.; Blader, I.J.; Fox, M.A. Toxoplasma infection induces microglia-neuron contact and the loss of perisomatic inhibitory synapses. Glia 2020, 68, 1968–1986. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.F.; Chandrasegaran, P.; Sinton, M.C.; Briggs, E.M.; Otto, T.D.; Heslop, R.; Bentley-Abbot, C.; Loney, C.; De Lecea, L.; Mabbott, N.A.; et al. Single cell and spatial transcriptomic analyses reveal microglia-plasma cell crosstalk in the brain during Trypanosoma brucei infection. Nat. Commun. 2022, 13, 5752. [Google Scholar] [CrossRef] [PubMed]
- Simard, A.R.; Soulet, D.; Gowing, G.; Julien, J.-P.; Rivest, S. Bone Marrow-Derived Microglia Play a Critical Role in Restricting Senile Plaque Formation in Alzheimer’s Disease. Neuron 2006, 49, 489–502. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.-j.; Li, M.-c.; Tao, H.; Wang, X. Targeting brain microvascular endothelial cells: A therapeutic approach to neuroprotection against stroke. Neural Regen. Res. 2015, 10, 1882. [Google Scholar] [CrossRef] [PubMed]
- Pong, S.; Karmacharya, R.; Sofman, M.; Bishop, J.R.; Lizano, P. The Role of Brain Microvascular Endothelial Cell and Blood-Brain Barrier Dysfunction in Schizophrenia. Complex Psychiatry 2020, 6, 30–46. [Google Scholar] [CrossRef] [PubMed]
- Takata, F.; Nakagawa, S.; Matsumoto, J.; Dohgu, S. Blood-Brain Barrier Dysfunction Amplifies the Development of Neuroinflammation: Understanding of Cellular Events in Brain Microvascular Endothelial Cells for Prevention and Treatment of BBB Dysfunction. Front. Cell. Neurosci. 2021, 15, 661838. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.-C.; Hsieh, Y.C.; Hu, C.J.; Yong Kwang, T. Endothelial Dysfunction in Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 2909. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, J.; Wang, J.; He, L.; Lai, H.; Zhang, T.; Wang, X.; Li, W. Mitochondrial Oxidative Stress in Brain Microvascular Endothelial Cells: Triggering Blood-Brain Barrier Disruption. Mitochondrion 2023, 69, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Körbelin, J.; Dogbevia, G.; Michelfelder, S.; Ridder, D.A.; Hunger, A.; Wenzel, J.; Seismann, H.; Lampe, M.; Bannach, J.; Pasparakis, M.; et al. A brain microvasculature endothelial cell-specific viral vector with the potential to treat neurovascular and neurological diseases. EMBO Mol. Med. 2016, 8, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Liu, Y.; Qi, H.; Ma, Q.; Xu, L.; Chen, W.; Chen, G.; Xu, X. A Novel Brain Neurovascular Unit Model with Neurons, Astrocytes and Microvascular Endothelial Cells of Rat. Int. J. Biol. Sci. 2013, 9, 174–189. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, B.B.; Grevesse, T.; Zimmerman, J.F.; Ardoña, H.A.M.; Jimenez, J.A.; Bitounis, D.; Demokritou, P.; Parker, K.K. Human brain microvascular endothelial cell pairs model tissue-level blood–brain barrier function. Integr. Biol. 2020, 12, 64–79. [Google Scholar] [CrossRef] [PubMed]
- Pikor, N.B.; Cupovic, J.; Onder, L.; Gommerman, J.L.; Ludewig, B. Stromal Cell Niches in the Inflamed Central Nervous System. J. Immunol. 2017, 198, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Do, P.T.; Wu, C.-C.; Chiang, Y.-H.; Hu, C.-J.; Chen, K.-Y. Mesenchymal Stem/Stromal Cell Therapy in Blood–Brain Barrier Preservation Following Ischemia: Molecular Mechanisms and Prospects. Int. J. Mol. Sci. 2021, 22, 10045. [Google Scholar] [CrossRef] [PubMed]
- Ramaglia, V.; Florescu, A.; Zuo, M.; Sheikh-Mohamed, S.; Gommerman, J.L. Stromal Cell–Mediated Coordination of Immune Cell Recruitment, Retention, and Function in Brain-Adjacent Regions. J. Immunol. 2021, 206, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W.; Faull, R.L.M.; Dragunow, M. A role for human brain pericytes in neuroinflammation. J. Neuroinflamm. 2014, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Kosyakova, N.; Kao, D.D.; Figetakis, M.; Francesc, L.-G.; Spindler, S.; Graham, M.; James, K.J.; Jee Won, S.; Liu, X.; Tietjen, G.T.; et al. Differential functional roles of fibroblasts and pericytes in the formation of tissue-engineered microvascular networks in vitro. NPJ Regen. Med. 2020, 5, 1038. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.D.; Winkler, E.A.; Sagare, A.P.; Singh, I.; LaRue, B.; Deane, R.; Zlokovic, B.V. Pericytes Control Key Neurovascular Functions and Neuronal Phenotype in the Adult Brain and during Brain Aging. Neuron 2010, 68, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Castro, V.; Toborek, M. Infection of human pericytes by HIV-1 disrupts the integrity of the blood–brain barrier. J. Mol. Cell Biol. 2012, 16, 2950–2957. [Google Scholar] [CrossRef] [PubMed]
- Bocci, M.; Oudenaarden, C.; Sàenz-Sardà, X.; Simrén, J.; Edén, A.; Sjölund, J.; Möller, C.; Gisslén, M.; Zetterberg, H.; Englund, E.; et al. Infection of Brain Pericytes Underlying Neuropathology of COVID-19 Patients. Int. J. Mol. Sci. 2021, 22, 11622. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.P.; Toledo, F.A.O.; Bautista, D.F.A.; Silva, M.F.; Oliveira, J.B.S.; Lima, P.A.; Costa, F.B.; Ribeiro, N.Q.; Lee, J.-Y.; Birbrair, A.; et al. Pericytes modulate endothelial inflammatory response during bacterial infection. mBio 2024, 15, e03252-23. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.A.; Bell, R.D.; Zlokovic, B.V. Central nervous system pericytes in health and disease. Nat. Neurosci. 2011, 14, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Rustenhoven, J.; Jansson, D.; Smyth, L.C.; Dragunow, M. Brain Pericytes As Mediators of Neuroinflammation. Trends Pharmacol. Sci. 2017, 38, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Hirunpattarasilp, C.; Attwell, D.; Freitas, F. The role of pericytes in brain disorders: From the periphery to the brain. J. Neurochem. 2019, 150, 648–665. [Google Scholar] [CrossRef] [PubMed]
- ElAli, A.; Thériault, P.; Rivest, S. The Role of Pericytes in Neurovascular Unit Remodeling in Brain Disorders. Int. J. Mol. Sci. 2014, 15, 6453–6474. [Google Scholar] [CrossRef] [PubMed]
- Velasco, S.; Kedaigle, A.J.; Simmons, S.K.; Nash, A.; Rocha, M.; Quadrato, G.; Paulsen, B.; Nguyen, L.; Adiconis, X.; Regev, A.; et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Paşca, S.P. Assembling human brain organoids. Science 2019, 363, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Liao, W.; Feng, N.-H.; Lou, Y.-L.; Niu, X.; Zhang, A.-J.; Wang, Y.; Deng, Z.-F. Human induced pluripotent stem cell-derived neural stem cells survive, migrate, differentiate, and improve neurologic function in a rat model of middle cerebral artery occlusion. Stem Cell. Res. Ther. 2013, 4, 73. [Google Scholar] [CrossRef] [PubMed]
- Wren, M.C.; Zhao, J.; Liu, C.-C.; Murray, M.E.; Atagi, Y.; Davis, M.D.; Fu, Y.; Okano, H.J.; Ogaki, K.; Strongosky, A.J.; et al. Frontotemporal dementia-associated N279K tau mutant disrupts subcellular vesicle trafficking and induces cellular stress in iPSC-derived neural stem cells. Mol. Neurodegener. 2015, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Piwocka, O.; Musielak, M.; Piotrowski, I.; Suchorska, W.M.; Trzeciak, T. From Donor to the Lab: A Fascinating Journey of Primary Cell Lines. Front. Cell Dev. Biol. 2021, 9, 711381. [Google Scholar] [CrossRef] [PubMed]
- Lopes, F.M.; Bristot, I.J.; da Motta, L.L.; Parsons, R.B.; Klamt, F. Mimicking Parkinson’s Disease in a Dish: Merits and Pitfalls of the Most Commonly used Dopaminergic In Vitro Models. Neuromol. Med. 2017, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Danz, K.; Höcherl, T.; Wien, S.L.; Wien, L.; Von Briesen, H.; Wagner, S. Experimental Comparison of Primary and hiPS-Based In Vitro Blood–Brain Barrier Models for Pharmacological Research. Pharmaceutics 2022, 14, 737. [Google Scholar] [CrossRef] [PubMed]
- Sivandzade, F.; Cucullo, L. In-vitro blood–brain barrier modeling: A review of modern and fast-advancing technologies. J. Cereb. Blood Flow Metab. 2018, 38, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Katsumiti, A.; Ruenraroengsak, P.; Cajaraville, M.P.; Thorley, A.J.; Tetley, T.D. Immortalisation of primary human alveolar epithelial lung cells using a non-viral vector to study respiratory bioreactivity in vitro. Sci. Rep. 2020, 10, 20486. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Liu, J.; Cheng, S.; Xing, Y.; Suo, W.Z. Differentiation renders susceptibility to excitotoxicity in HT22 neurons. Neural Regen. Res. 2013, 8, 1297–1306. [Google Scholar] [PubMed]
- Chua, P.; Lim, W.K. Optimisation of a PC12 cell-based in vitro stroke model for screening neuroprotective agents. Sci. Rep. 2021, 11, 8096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-m.; Yin, M.; Zhang, M.-h. Cell-based assays for Parkinson’s disease using differentiated human LUHMES cells. Acta Pharmacol. Sin. 2014, 35, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Slanzi, A.; Iannoto, G.; Rossi, B.; Zenaro, E.; Constantin, G. In vitro Models of Neurodegenerative Diseases. Front. Cell Dev. Biol. 2020, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Dufour, J.M. Cell Lines. Spermatogenesis 2012, 2, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Cattaneo, E. Neural stem cell systems: Physiological players or in vitro entities? Nat. Rev. Neurosci. 2010, 11, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Martino, G.; Pluchino, S. The therapeutic potential of neural stem cells. Nat. Rev. Neurosci. 2006, 7, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Johansson, C.B.; Momma, S.; Clarke, D.L.; Risling, M.; Lendahl, U.; Frisén, J. Identification of a Neural Stem Cell in the Adult Mammalian Central Nervous System. Cell 1999, 96, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Kang, E.; Liu, C.Y.; Ming, G.-L.; Song, H. Development of neural stem cell in the adult brain. Curr. Opin. Neurobiol. 2008, 18, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Huangfu, D. Human pluripotent stem cells: An emerging model in developmental biology. Development 2013, 140, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Bates, N.; Soteriou, D.; Grady, L.; Edmond, C.; Ross, A.; Kerby, A.; Lewis, P.A.; Adeniyi, T.; Wright, R.; et al. High quality clinical grade human embryonic stem cell lines derived from fresh discarded embryos. Stem Cell. Res. Ther. 2017, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Romito, A.; Cobellis, G. Pluripotent Stem Cells: Current Understanding and Future Directions. Stem Cells Int. 2016, 2016, 9451492. [Google Scholar] [CrossRef]
- Tristán-Noguero, A.; Fernández-Carasa, I.; Calatayud, C.; Bermejo-Casadesús, C.; Pons-Espinal, M.; Colini Baldeschi, A.; Campa, L.; Artigas, F.; Bortolozzi, A.; Domingo-Jiménez, R.; et al. iPSC-based modeling of THD recapitulates disease phenotypes and reveals neuronal malformation. EMBO Mol. Med. 2023, 15, e15847. [Google Scholar] [CrossRef] [PubMed]
- Pietiläinen, O.; Trehan, A.; Meyer, D.; Mitchell, J.; Tegtmeyer, M.; Valakh, V.; Gebre, H.; Chen, T.; Vartiainen, E.; Farhi, S.L.; et al. Astrocytic cell adhesion genes linked to schizophrenia correlate with synaptic programs in neurons. Cell Rep. 2023, 42, 111988. [Google Scholar] [CrossRef] [PubMed]
- Dräger, N.M.; Sattler, S.M.; Huang, C.T.-L.; Teter, O.M.; Leng, K.; Hashemi, S.H.; Hong, J.; Aviles, G.; Clelland, C.D.; Zhan, L.; et al. A CRISPRi/a platform in human iPSC-derived microglia uncovers regulators of disease states. Nat. Neurosci. 2022, 25, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, M.J.; Gastfriend, B.D.; Canfield, S.G.; Lee, M.-S.; Richards, D.; Faubion, M.G.; Li, W.-J.; Daneman, R.; Palecek, S.P.; Shusta, E.V. Human pluripotent stem cell–derived brain pericyte–like cells induce blood-brain barrier properties. Sci. Adv. 2019, 5, eaau7375. [Google Scholar] [CrossRef] [PubMed]
- Di Lullo, E.; Kriegstein, A.R. The use of brain organoids to investigate neural development and disease. Nat. Rev. Neurosci. 2017, 18, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Sloan, S.A.; Andersen, J.; Pașca, A.M.; Birey, F.; Pașca, S.P. Generation and assembly of human brain region–specific three-dimensional cultures. Nat. Protoc. 2018, 13, 2062–2085. [Google Scholar] [CrossRef] [PubMed]
- Birey, F.; Andersen, J.; Makinson, C.D.; Islam, S.; Wei, W.; Huber, N.; Fan, H.C.; Metzler, K.R.C.; Panagiotakos, G.; Thom, N.; et al. Assembly of functionally integrated human forebrain spheroids. Nature 2017, 545, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-P.; Qiu, Y.; Zhang, B.; Chen, G.; Chen, Q.; Wang, M.; Mo, F.; Xu, J.; Wu, J.; Zhang, R.-R.; et al. Zika virus infection induces RNAi-mediated antiviral immunity in human neural progenitors and brain organoids. Cell Res. 2019, 29, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef]
- Qian, X.; Song, H.; Ming, G.-l. Brain organoids: Advances, applications and challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef] [PubMed]
- Lokai, T.; Albin, B.; Qubbaj, K.; Tiwari, A.P.; Adhikari, P.; Yang, I.H. A review on current brain organoid technologies from a biomedical engineering perspective. Exp. Neurol. 2023, 367, 114461. [Google Scholar] [CrossRef]
- Renner, M.; Lancaster, M.A.; Bian, S.; Choi, H.; Ku, T.; Peer, A.; Chung, K.; Knoblich, J.A. Self-organized developmental patterning and differentiation in cerebral organoids. EMBO J. 2017, 36, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.D.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E.; et al. Cerebral organoids at the air–liquid interface generate diverse nerve tracts with functional output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, L.; Bonfio, C.; Chadwick, J.; Begum, F.; Skehel, M.; Lancaster, M.A. Human CNS barrier-forming organoids with cerebrospinal fluid production. Science 2020, 369, eaaz5626. [Google Scholar] [CrossRef] [PubMed]
- Bagley, J.A.; Reumann, D.; Bian, S.; Lévi-Strauss, J.; Knoblich, J.A. Fused cerebral organoids model interactions between brain regions. Nat. Methods 2017, 14, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Tambalo, M.; Lodato, S. Brain organoids: Human 3D models to investigate neuronal circuits assembly, function and dysfunction. Brain Res. 2020, 1746, 147028. [Google Scholar] [CrossRef] [PubMed]
- Lindborg, B.A.; Brekke, J.H.; Vegoe, A.L.; Ulrich, C.B.; Haider, K.T.; Subramaniam, S.; Venhuizen, S.L.; Eide, C.R.; Orchard, P.J.; Chen, W.; et al. Rapid Induction of Cerebral Organoids from Human Induced Pluripotent Stem Cells Using a Chemically Defined Hydrogel and Defined Cell Culture Medium. Stem Cells Transl. Med. 2016, 5, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-J.; Elahi, L.S.; Pașca, A.M.; Marton, R.M.; Gordon, A.; Revah, O.; Miura, Y.; Walczak, E.M.; Holdgate, G.M.; Fan, H.C.; et al. Reliability of human cortical organoid generation. Nat. Methods 2019, 16, 75–78. [Google Scholar] [CrossRef]
- Paşca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Chou, V.T.; Johnson, S.A.; Van Vactor, D. Synapse development and maturation at the drosophila neuromuscular junction. Neural Dev. 2020, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Xiao, Y.; Sun, A.X.; Cukuroglu, E.; Tran, H.-D.; Göke, J.; Tan, Z.Y.; Saw, T.Y.; Tan, C.-P.; Lokman, H.; et al. Midbrain-like Organoids from Human Pluripotent Stem Cells Contain Functional Dopaminergic and Neuromelanin-Producing Neurons. Cell Stem Cell 2016, 19, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.V.; Emre, M.; Jenner, P.; Poewe, W. Levodopa in the treatment of Parkinson’s disease. Eur. J. Neurol. 2009, 16, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Splawski, I.; Timothy, K.W.; Sharpe, L.M.; Decher, N.; Kumar, P.; Bloise, R.; Napolitano, C.; Schwartz, P.J.; Joseph, R.M.; Condouris, K.; et al. CaV1.2 Calcium Channel Dysfunction Causes a Multisystem Disorder Including Arrhythmia and Autism. Cell 2004, 119, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Bortone, D.; Polleux, F. KCC2 expression promotes the termination of cortical interneuron migration in a voltage-sensitive calcium-dependent manner. Neuron 2009, 62, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Li, M.-Y.; Birey, F.; Ikeda, K.; Revah, O.; Thete, M.V.; Park, J.-Y.; Puno, A.; Lee, S.H.; Porteus, M.H.; et al. Generation of human striatal organoids and cortico-striatal assembloids from human pluripotent stem cells. Nat. Biotechnol. 2020, 38, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Revah, O.; Miura, Y.; Thom, N.; Amin, N.D.; Kelley, K.W.; Singh, M.; Chen, X.; Thete, M.V.; Walczak, E.M.; et al. Generation of Functional Human 3D Cortico-Motor Assembloids. Cell 2020, 183, 1913–1929.e26. [Google Scholar] [CrossRef]
- Miura, Y.; Li, M.-Y.; Revah, O.; Yoon, S.-J.; Narazaki, G.; Pașca, S.P. Engineering brain assembloids to interrogate human neural circuits. Nat. Protoc. 2022, 17, 15–35. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-N.; Jin, Y.; An, Y.; Kim, J.; Choi, Y.S.; Lee, J.S.; Kim, J.; Choi, W.-Y.; Koo, D.-J.; Yu, W.; et al. Microfluidic device with brain extracellular matrix promotes structural and functional maturation of human brain organoids. Nat. Commun. 2021, 12, 4730. [Google Scholar] [CrossRef] [PubMed]
- Cakir, B.; Xiang, Y.; Tanaka, Y.; Kural, M.H.; Parent, M.; Kang, Y.-J.; Chapeton, K.; Patterson, B.; Yuan, Y.; He, C.-S.; et al. Engineering of human brain organoids with a functional vascular-like system. Nat. Methods 2019, 16, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Amirifar, L.; Shamloo, A.; Nasiri, R.; de Barros, N.R.; Wang, Z.Z.; Unluturk, B.D.; Libanori, A.; Ievglevskyi, O.; Diltemiz, S.E.; Sances, S.; et al. Brain-on-a-chip: Recent advances in design and techniques for microfluidic models of the brain in health and disease. Biomaterials 2022, 285, 121531. [Google Scholar] [CrossRef] [PubMed]
- Maoz, B.M. Brain-on-a-Chip: Characterizing the next generation of advanced in vitro platforms for modeling the central nervous system. APL Bioeng. 2021, 5, 030902. [Google Scholar] [CrossRef] [PubMed]
- Oldendorf, W.H. The blood-brain barrier. Exp. Eye Res. 1977, 25, 177–190. [Google Scholar] [CrossRef] [PubMed]
- de Vries, H.E.; Blom-Roosemalen, M.C.M.; Oosten, M.v.; de Boer, A.G.; van Berkel, T.J.C.; Breimer, D.D.; Kuiper, J. The influence of cytokines on the integrity of the blood-brain barrier in vitro. J. Neuroimmunol. 1996, 64, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Wevers, N.R.; Kasi, D.G.; Gray, T.; Wilschut, K.J.; Smith, B.; van Vught, R.; Shimizu, F.; Sano, Y.; Kanda, T.; Marsh, G.; et al. A perfused human blood–brain barrier on-a-chip for high-throughput assessment of barrier function and antibody transport. Fluids Barriers CNS 2018, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Cho, S.-W. Blood-brain barrier-on-a-chip for brain disease modeling and drug testing. BMB Rep. 2022, 55, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Griep, L.M.; Wolbers, F.; De Wagenaar, B.; Ter Braak, P.M.; Weksler, B.B.; Romero, I.A.; Couraud, P.O.; Vermes, I.; Van Der Meer, A.D.; Van Den Berg, A. BBB ON CHIP: Microfluidic platform to mechanically and biochemically modulate blood-brain barrier function. Biomed. Microdevices 2013, 15, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Nivasini; Foo, L.C.; Ng, S.H.; Hunziker, W.; Choudhury, D. A pump-free tricellular blood–brain barrier on-a-chip model to understand barrier property and evaluate drug response. Biotechnol. Bioeng. 2020, 117, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.J.; Diep, Y.N.; Tran, M.; Tran, V.T.A.; Ambrin, G.; Ngo, H.; Cho, H. Three-dimensional human neural culture on a chip recapitulating neuroinflammation and neurodegeneration. Nat. Protoc. 2023, 18, 2838–2867. [Google Scholar] [CrossRef]
- Pediaditakis, I.; Kodella, K.R.; Manatakis, D.V.; Le, C.Y.; Barthakur, S.; Sorets, A.; Gravanis, A.; Ewart, L.; Rubin, L.L.; Manolakos, E.S.; et al. A microengineered Brain-Chip to model neuroinflammation in humans. iScience 2022, 25, 104813. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Chu, S.; Yang, Y.; Zhang, Z.; Pang, Z.; Chen, N. Neuroinflammatory In Vitro Cell Culture Models and the Potential Applications for Neurological Disorders. Front. Pharmacol. 2021, 12, 671734. [Google Scholar] [CrossRef] [PubMed]
- Herland, A.; van der Meer, A.D.; FitzGerald, E.A.; Park, T.-E.; Sleeboom, J.J.F.; Ingber, D.E. Distinct Contributions of Astrocytes and Pericytes to Neuroinflammation Identified in a 3D Human Blood-Brain Barrier on a Chip. PLoS ONE 2016, 11, e0150360. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, I.; Izzo, L.; Tunesi, M.; Comar, M.; Albani, D.; Giordano, C. Organ-On-A-Chip in vitro Models of the Brain and the Blood-Brain Barrier and Their Value to Study the Microbiota-Gut-Brain Axis in Neurodegeneration. Front. Bioeng. Biotechnol. 2020, 7, 435. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, M.T.; Albani, D.; Giordano, C. An Organ-On-A-Chip Engineered Platform to Study the Microbiota–Gut–Brain Axis in Neurodegeneration. Trends Mol. Med. 2019, 25, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H.; Kim, D.; Sung, J.H. A Gut-Brain Axis-on-a-Chip for studying transport across epithelial and endothelial barriers. J. Ind. Eng. Chem. 2021, 101, 126–134. [Google Scholar] [CrossRef]
- Kim, M.-H.; van Noort, D.; Sung, J.H.; Park, S. Organ-on-a-Chip for Studying Gut-Brain Interaction Mediated by Extracellular Vesicles in the Gut Microenvironment. Int. J. Mol. Sci. 2021, 22, 13513. [Google Scholar] [CrossRef] [PubMed]
- Azzoni, R.; Marsland, B.J. The lung-brain axis: A new frontier in host-microbe interactions. Immunity 2022, 55, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, T.; Yuan, C.; Zhong, J.-S.; Huang, J.-D.; Ke, Y.; Sun, H. The Lung Microbiome: A New Frontier for Lung and Brain Disease. Int. J. Mol. Sci. 2023, 24, 2170. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Bajinka, O.; Tang, Z.; Qiu, X.; Tan, Y. Lung–brain axis: Metabolomics and pathological changes in lungs and brain of respiratory syncytial virus-infected mice. J. Med. Virol. 2022, 94, 5885–5893. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-S.; Baez, C.E.; Atala, A. Biomaterials for tissue engineering. World J. Urol. 2000, 18, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.P.; Appel, E.A.; Ashby, P.D.; Baker, B.M.; Franco, E.; Gu, L.; Haynes, K.; Joshi, N.S.; Kloxin, A.M.; Kouwer, P.H.J.; et al. The living interface between synthetic biology and biomaterial design. Nat. Mater. 2022, 21, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Choi, E.J.; Cho, J.H.; Cho, A.-N.; Jin, Y.; Yang, K.; Song, C.; Cho, S.-W. Three-Dimensional Electroconductive Hyaluronic Acid Hydrogels Incorporated with Carbon Nanotubes and Polypyrrole by Catechol-Mediated Dispersion Enhance Neurogenesis of Human Neural Stem Cells. Biomacromolecules 2017, 18, 3060–3072. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.G.; Brunel, L.G.; Huang, M.S.; Liu, Y.; Cai, B.; Sinha, S.; Yang, F.; Pașca, S.P.; Shin, S.; Heilshorn, S.C. Spatially controlled construction of assembloids using bioprinting. Nat. Commun. 2023, 14, 4346. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Lee, J.-S.; Gao, G.; Byoung Soo, K.; Cho, D.-W. 3D bioprinted multilayered cerebrovascular conduits to study cancer extravasation mechanism related with vascular geometry. Nat. Commun. 2023, 14, 7696. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Mikhailova, E.; Lei, M.; Cowley, S.A.; Sun, T.; Yang, X.; Zhang, Y.; Liu, K.; Catarino da Silva, D.; Campos Soares, L.; et al. Integration of 3D-printed cerebral cortical tissue into an ex vivo lesioned brain slice. Nat. Commun. 2023, 14, 5986. [Google Scholar] [CrossRef] [PubMed]
- Grebenyuk, S.; Abdel Fattah, A.R.; Kumar, M.; Toprakhisar, B.; Rustandi, G.; Vananroye, A.; Salmon, I.; Verfaillie, C.; Grillo, M.; Ranga, A. Large-scale perfused tissues via synthetic 3D soft microfluidics. Nat. Commun. 2023, 14, 193. [Google Scholar] [CrossRef] [PubMed]
- Abdel Fattah, A.R.; Daza, B.; Rustandi, G.; Berrocal-Rubio, M.Á.; Gorissen, B.; Poovathingal, S.; Davie, K.; Barrasa-Fano, J.; Cóndor, M.; Cao, X.; et al. Actuation enhances patterning in human neural tube organoids. Nat. Commun. 2021, 12, 3192. [Google Scholar] [CrossRef] [PubMed]
- Rifes, P.; Isaksson, M.; Rathore, G.S.; Aldrin-Kirk, P.; Møller, O.K.; Barzaghi, G.; Lee, J.; Egerod, K.L.; Rausch, D.M.; Parmar, M.; et al. Modeling neural tube development by differentiation of human embryonic stem cells in a microfluidic WNT gradient. Nat. Biotechnol. 2020, 38, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood–brain barrier. Neurobiol. Dis. 2020, 146, 105131. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, P.; Cricca, M.; Nastasi, C. Are the Organoid Models an Invaluable Contribution to ZIKA Virus Research? Pathogens 2021, 10, 1233. [Google Scholar] [CrossRef] [PubMed]
- Garcez, P.P.; Loiola, E.C.; Madeiro da Costa, R.; Higa, L.M.; Trindade, P.; Delvecchio, R.; Nascimento, J.M.; Brindeiro, R.; Tanuri, A.; Rehen, S.K. Zika virus impairs growth in human neurospheres and brain organoids. Science 2016, 352, 816–818. [Google Scholar] [CrossRef] [PubMed]
- Alimonti, J.B.; Ribecco-Lutkiewicz, M.; Sodja, C.; Jezierski, A.; Stanimirovic, D.B.; Liu, Q.; Haqqani, A.S.; Conlan, W.; Bani-Yaghoub, M. Zika virus crosses an in vitro human blood brain barrier model. Fluids Barriers CNS 2018, 15, 15. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.A.; Rhea, E.M.; Knopp, R.C.; Banks, W.A. Interactions of SARS-CoV-2 with the Blood–Brain Barrier. Int. J. Mol. Sci. 2021, 22, 2681. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, W.; Chen, F.; Cui, L. COVID-19 and cognitive impairment: Neuroinvasive and blood-brain barrier dysfunction. J. Neuroinflamm. 2022, 19, 222. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.-E.; Madore, C.; Bordeleau, M.; Tian, L.; Verkhratsky, A. Neuropathobiology of COVID-19: The Role for Glia. Front. Cell. Neurosci. 2020, 14, 592214. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T. Neurological infection with SARS-CoV-2—The story so far. Nat. Rev. Neurol. 2021, 17, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Ramani, A.; Müller, L.; Ostermann, P.N.; Gabriel, E.; Abida-Islam, P.; Müller-Schiffmann, A.; Mariappan, A.; Goureau, O.; Gruell, H.; Walker, A.; et al. SARS-CoV-2 targets neurons of 3D human brain organoids. EMBO J. 2020, 39, e106230. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Pather, S.R.; Huang, W.-K.; Zhang, F.; Wong, S.Z.H.; Zhou, H.; Cubitt, B.; Fan, W.; Chen, C.Z.; Xu, M.; et al. Human Pluripotent Stem Cell-Derived Neural Cells and Brain Organoids Reveal SARS-CoV-2 Neurotropism Predominates in Choroid Plexus Epithelium. Cell Stem Cell 2020, 27, 937–950.e9. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Mármol, R.; Giordano-Santini, R.; Kaulich, E.; Cho, A.N.; Przybyla, M.; Riyadh, M.A.; Robinson, E.; Chew, K.Y.; Amor, R.; Meunier, F.A.; et al. SARS-CoV-2 infection and viral fusogens cause neuronal and glial fusion that compromises neuronal activity. Sci. Adv. 2023, 9, eadg2248. [Google Scholar] [CrossRef] [PubMed]
- Aguado, J.; Amarilla, A.A.; Taherian Fard, A.; Albornoz, E.A.; Tyshkovskiy, A.; Schwabenland, M.; Chaggar, H.K.; Modhiran, N.; Gómez-Inclán, C.; Javed, I.; et al. Senolytic therapy alleviates physiological human brain aging and COVID-19 neuropathology. Nat. Aging. 2023, 3, 1561–1575. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Kishimoto, Y.; Grammatikakis, I.; Gottimukkala, K.; Cutler, R.G.; Zhang, S.; Abdelmohsen, K.; Bohr, V.A.; Misra Sen, J.; Gorospe, M.; et al. Senolytic therapy alleviates Aβ-associated oligodendrocyte progenitor cell senescence and cognitive deficits in an Alzheimer’s disease model. Nat. Neurosci. 2019, 22, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sievert, D.; Clark, A.E.; Lee, S.; Federman, H.; Gastfriend, B.D.; Shusta, E.V.; Palecek, S.P.; Carlin, A.F.; Gleeson, J.G. A human three-dimensional neural-perivascular ‘assembloid’ promotes astrocytic development and enables modeling of SARS-CoV-2 neuropathology. Nat. Med. 2021, 27, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Fares, M.; Cochet-Bernoin, M.; Gonzalez, G.; Montero-Menei, C.N.; Blanchet, O.; Benchoua, A.; Boissart, C.; Lecollinet, S.; Richardson, J.; Haddad, N.; et al. Pathological modeling of TBEV infection reveals differential innate immune responses in human neurons and astrocytes that correlate with their susceptibility to infection. J. Neuroinflamm. 2020, 17, 76. [Google Scholar] [CrossRef] [PubMed]
- dos Reis, R.S.; Sant, S.; Keeney, H.; Wagner, M.C.E.; Ayyavoo, V. Modeling HIV-1 neuropathogenesis using three-dimensional human brain organoids (hBORGs) with HIV-1 infected microglia. Sci. Rep. 2020, 10, 15209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, H.; Dong, L.; Xia, Q. Antiviral and Neuroprotective Abilities of Influenza Virus Infection in Tractable Brain Organoids. bioRxiv 2022, 482634. [Google Scholar] [CrossRef]
- Cairns, D.M.; Rouleau, N.; Parker, R.N.; Walsh, K.G.; Gehrke, L.; Kaplan, D.L. A 3D human brain–like tissue model of herpes-induced Alzheimer’s disease. Sci. Adv. 2020, 6, eaay8828. [Google Scholar] [CrossRef] [PubMed]
- Barreras, P.; Pamies, D.; Monaco, M.C.; Muñoz, L.S.; Zhong, X.; Major, E.O.; Hogberg, H.T.; Hartung, T.; Pardo, C.A. A human-derived 3D brain organoid model to study JC virus infection. J. Neurovirol. 2022, 28, 17–26. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, L.; Naciri, J.; Radio, N.; Tekur, S.; Clayton, D.; Apodaca, G.; Di Maio, R.; Zhi, Y.; Dimitrion, P.; Piazza, P.; et al. Generation of three-dimensional human neuronal cultures: Application to modeling CNS viral infections. Stem Cell. Res. Ther. 2018, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Koutsouras, G.W.; Ramos, R.L.; Martinez, L.R. Role of microglia in fungal infections of the central nervous system. Virulence 2017, 8, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kyung Tae, L.; Jong Seung, L.; Shin, J.; Cui, B.; Yang, K.; Yi Sun, C.; Choi, N.; Soo Hyun, L.; Jae Hyun, L.; et al. Fungal brain infection modelled in a human-neurovascular-unit-on-a-chip with a functional blood–brain barrier. Nat. Biomed. Eng. 2021, 5, 830–846. [Google Scholar] [CrossRef] [PubMed]
- Al-Obaidi, M.M.J.; Desa, M.N.M. Mechanisms of Blood Brain Barrier Disruption by Different Types of Bacteria, and Bacterial–Host Interactions Facilitate the Bacterial Pathogen Invading the Brain. Cell. Mol. Neurobiol. 2018, 38, 1349–1368. [Google Scholar] [CrossRef] [PubMed]
- Benmimoun, B.; Papastefanaki, F.; Périchon, B.; Segklia, K.; Roby, N.; Miriagou, V.; Schmitt, C.; Dramsi, S.; Matsas, R.; Spéder, P. An original infection model identifies host lipoprotein import as a route for blood-brain barrier crossing. Nat. Commun. 2020, 11, 6106. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, A.; Rochfort, K.D.; McDonnell, C.J.; Kerrigan, S.W.; Cummins, P.M. Staphylococcus aureus-mediated blood-brain barrier injury: An in vitro human brain microvascular endothelial cell model. Cell. Microbiol. 2017, 19, e12664. [Google Scholar] [CrossRef] [PubMed]
- Endres, L.M.; Jungblut, M.; Divyapicigil, M.; Sauer, M.; Stigloher, C.; Christodoulides, M.; Kim, B.J.; Schubert-Unkmeir, A. Development of a multicellular in vitro model of the meningeal blood-CSF barrier to study Neisseria meningitidis infection. Fluids Barriers CNS 2022, 19, 81. [Google Scholar] [CrossRef] [PubMed]
- Halonen, S.K. Use of in vitro derived human neuronal models to study host-parasite interactions of Toxoplasma gondii in neurons and neuropathogenesis of chronic toxoplasmosis. Front. Cell. Infect. Microbiol. 2023, 13, 1129451. [Google Scholar] [CrossRef] [PubMed]
- Halonen, S.K. Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons. Cells 2017, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.-H.; Han, H.-W.; Lee, S.-E.; Hong, S.-H.; Cho, S.-H.; Kim, S.C.; Koo, S.K.; Kim, J.-H. Modelling Toxoplasma gondii infection in human cerebral organoids. Emerg. Microbes Infect. 2020, 9, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Correa Leite, P.E.; de Araujo Portes, J.; Pereira, M.R.; Russo, F.B.; Martins-Duarte, E.S.; Almeida dos Santos, N.; Attias, M.; Barrantes, F.J.; Baleeiro Beltrão-Braga, P.C.; de Souza, W. Morphological and biochemical repercussions of Toxoplasma gondii infection in a 3D human brain neurospheres model. Brain Behav. Immun. Health 2021, 11, 100190. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Velazquez, D.; Betanzos, A.; Serrano-Luna, J.; Shibayama, M. An In Vitro Model of the Blood-Brain Barrier: Naegleria fowleri Affects the Tight Junction Proteins and Activates the Microvascular Endothelial Cells. J. Eukaryot. Microbiol. 2018, 65, 804–819. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-C.E.; Jodat, Y.A.; Samanipour, R.; Zorzi, G.; Zhu, K.; Hirano, M.; Chang, K.; Arnaout, A.; Hassan, S.; Matharu, N.; et al. Toward a neurospheroid niche model: Optimizing embedded 3D bioprinting for fabrication of neurospheroid brain-like co-culture constructs. Biofabrication 2020, 13, 015014. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Rich, J.N.; Chen, S. Biomaterials and 3D Bioprinting Strategies to Model Glioblastoma and the Blood–Brain Barrier. Adv. Mater. 2021, 33, 2004776. [Google Scholar] [CrossRef] [PubMed]
- Esworthy, T.J.; Miao, S.; Lee, S.-J.; Zhou, X.; Cui, H.; Zuo, Y.Y.; Zhang, L.G. Advanced 4D-bioprinting technologies for brain tissue modeling and study. Int. J. Smart Nano Mater. 2019, 10, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Chen, X.; Dowbaj, A.M.; Sljukic, A.; Bratlie, K.; Lin, L.; Fong, E.L.S.; Balachander, G.M.; Chen, Z.; Soragni, A.; et al. Organoids. Nat. Rev. Methods Primers 2022, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Qiao, H.; Xu, F.; Zhao, W.; Wang, J.; Gu, L.; Chen, P.; Peng, M. Bioengineering of a human physiologically relevant microfluidic blood-cerebrospinal fluid barrier model. Lab Chip 2023, 23, 32–315. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Tang, B.; Romero, J.C.; Yang, Y.; Elsayed, S.K.; Pahapale, G.; Lee, T.-J.; Morales Pantoja, I.E.; Han, F.; Berlinicke, C.; et al. Shell microelectrode arrays (MEAs) for brain organoids. Sci. Adv. 2022, 8, eabq5031. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Forró, C.; Li, T.L.; Miura, Y.; Zaluska, T.J.; Tsai, C.-T.; Kanton, S.; McQueen, J.P.; Chen, X.; Mollo, V.; et al. Kirigami electronics for long-term electrophysiological recording of human neural organoids and assembloids. Nat. Biotechnol. 2024, 1, 1038. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Koo, B.-K.; Knoblich, J.A. Human organoids: Model systems for human biology and medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Tebon, P.J.; Wang, B.; Markowitz, A.L.; Davarifar, A.; Tsai, B.L.; Krawczuk, P.; Gonzalez, A.E.; Sartini, S.; Murray, G.F.; Nguyen, H.T.L.; et al. Drug screening at single-organoid resolution via bioprinting and interferometry. Nat. Commun. 2023, 14, 3168. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-Y.; Wu, S.; Wang, D.; Chu, C.; Hong, Y.; Tao, M.; Hu, H.; Xu, M.; Guo, X.; Liu, Y. Human organoids in basic research and clinical applications. Signal Transduct. Target. Ther. 2022, 7, 168. [Google Scholar] [CrossRef] [PubMed]
- Qing, L.-S.; Wang, T.-T.; Luo, H.-Y.; Du, J.-L.; Wang, R.-Y.; Luo, P. Microfluidic strategies for natural products in drug discovery: Current status and future perspectives. TrAC Trends Anal. Chem. 2023, 158, 116832. [Google Scholar] [CrossRef]
- Schuster, B.; Junkin, M.; Kashaf, S.S.; Romero-Calvo, I.; Kirby, K.; Matthews, J.; Weber, C.R.; Rzhetsky, A.; White, K.P.; Tay, S. Automated microfluidic platform for dynamic and combinatorial drug screening of tumor organoids. Nat. Commun. 2020, 11, 5271. [Google Scholar] [CrossRef] [PubMed]
- Menche, C.; Farin, H.F. Strategies for genetic manipulation of adult stem cell-derived organoids. Exp. Mol. Med. 2021, 53, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, G.; Babu, P.E.; Singh, R.; Trehanpati, N. Application of CRISPR-Cas9 based gene editing to study the pathogenesis of colon and liver cancer using organoids. Hepatol. Int. 2021, 15, 1309–1317. [Google Scholar] [CrossRef]
- Lange, J.; Zhou, H.; McTague, A. Cerebral Organoids and Antisense Oligonucleotide Therapeutics: Challenges and Opportunities. Front. Mol. Neurosci. 2022, 15, 941528. [Google Scholar] [CrossRef] [PubMed]
- Quadrato, G.; Brown, J.; Arlotta, P. The promises and challenges of human brain organoids as models of neuropsychiatric disease. Nat. Med. 2016, 22, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Morrocchi, E.; van Haren, S.; Palma, P.; Levy, O. Modeling human immune responses to vaccination in vitro. Trends Immunol. 2024, 45, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, L.C.; Costa, P.A.C.; Scalzo Júnior, S.R.A.; Ferreira, H.A.S.; Braga, A.C.S.; de Oliveira, L.C.; Figueiredo, M.M.; Shepherd, S.; Hamilton, A.; Queiroz-Junior, C.M.; et al. Nanoparticle-based DNA vaccine protects against SARS-CoV-2 variants in female preclinical models. Nat. Commun. 2024, 15, 590. [Google Scholar] [CrossRef] [PubMed]
- Hammel, J.H.; Zatorski, J.M.; Cook, S.R.; Pompano, R.R.; Munson, J.M. Engineering in vitro immune-competent tissue models for testing and evaluation of therapeutics. Adv. Drug Del. Rev. 2022, 182, 114111. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, S.; Lin, X.; Chen, G.; Kang, J.; Ma, Z.; Wang, Y.; Li, Z.; Xiao, X.; He, A.; et al. Application of Organoids in Carcinogenesis Modeling and Tumor Vaccination. Front. Oncol. 2022, 12, 855996. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Pak, J.; Doss-Gollin, S.; Ryff, K.; Beijnen, E.; Pedersen, G.K.; Christensen, D.; Levy, O.; van Haren, S.D. Human In vitro Modeling Identifies Adjuvant Combinations that Unlock Antigen Cross-presentation and Promote T-helper 1 Development in Newborns, Adults and Elders. J. Mol. Biol. 2024, 436, 168446. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Cho, A.-N. Human Brain In Vitro Model for Pathogen Infection-Related Neurodegeneration Study. Int. J. Mol. Sci. 2024, 25, 6522. https://doi.org/10.3390/ijms25126522
Yan Y, Cho A-N. Human Brain In Vitro Model for Pathogen Infection-Related Neurodegeneration Study. International Journal of Molecular Sciences. 2024; 25(12):6522. https://doi.org/10.3390/ijms25126522
Chicago/Turabian StyleYan, Yuwei, and Ann-Na Cho. 2024. "Human Brain In Vitro Model for Pathogen Infection-Related Neurodegeneration Study" International Journal of Molecular Sciences 25, no. 12: 6522. https://doi.org/10.3390/ijms25126522
APA StyleYan, Y., & Cho, A.-N. (2024). Human Brain In Vitro Model for Pathogen Infection-Related Neurodegeneration Study. International Journal of Molecular Sciences, 25(12), 6522. https://doi.org/10.3390/ijms25126522