
Lysophosphatidylcholines Promote Influenza Virus Reproduction through the MAPK/JNK Pathway in PMA-Differentiated THP-1 Macrophages
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. LPCs Decrease the Survival of Cells Infected with IAV
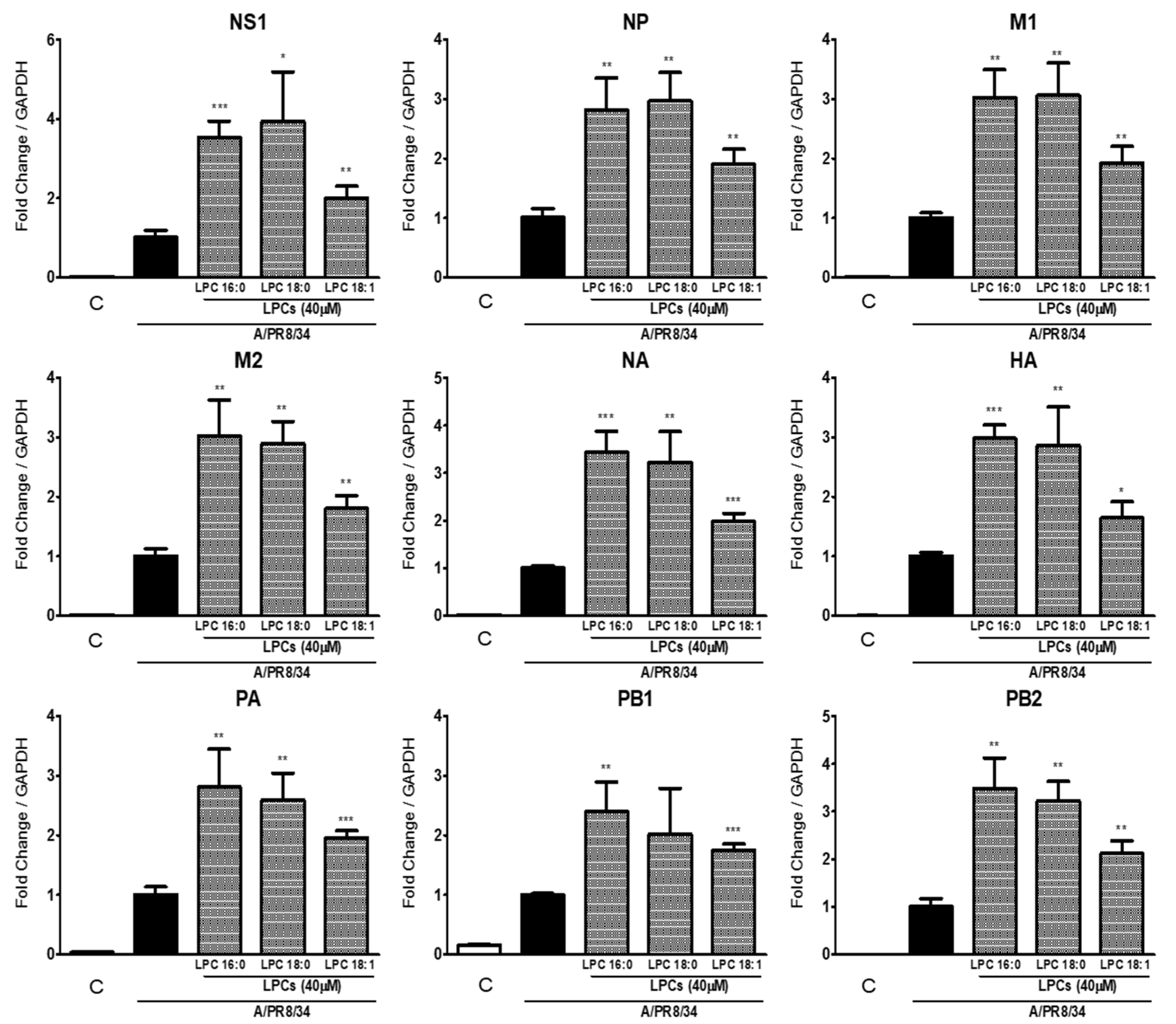
2.2. LPCs Increase Viral RNA and Protein Expression
2.3. Saturated LPCs Further Increase IAV RNA Levels
2.4. LPCs Alter Host Gene Expression
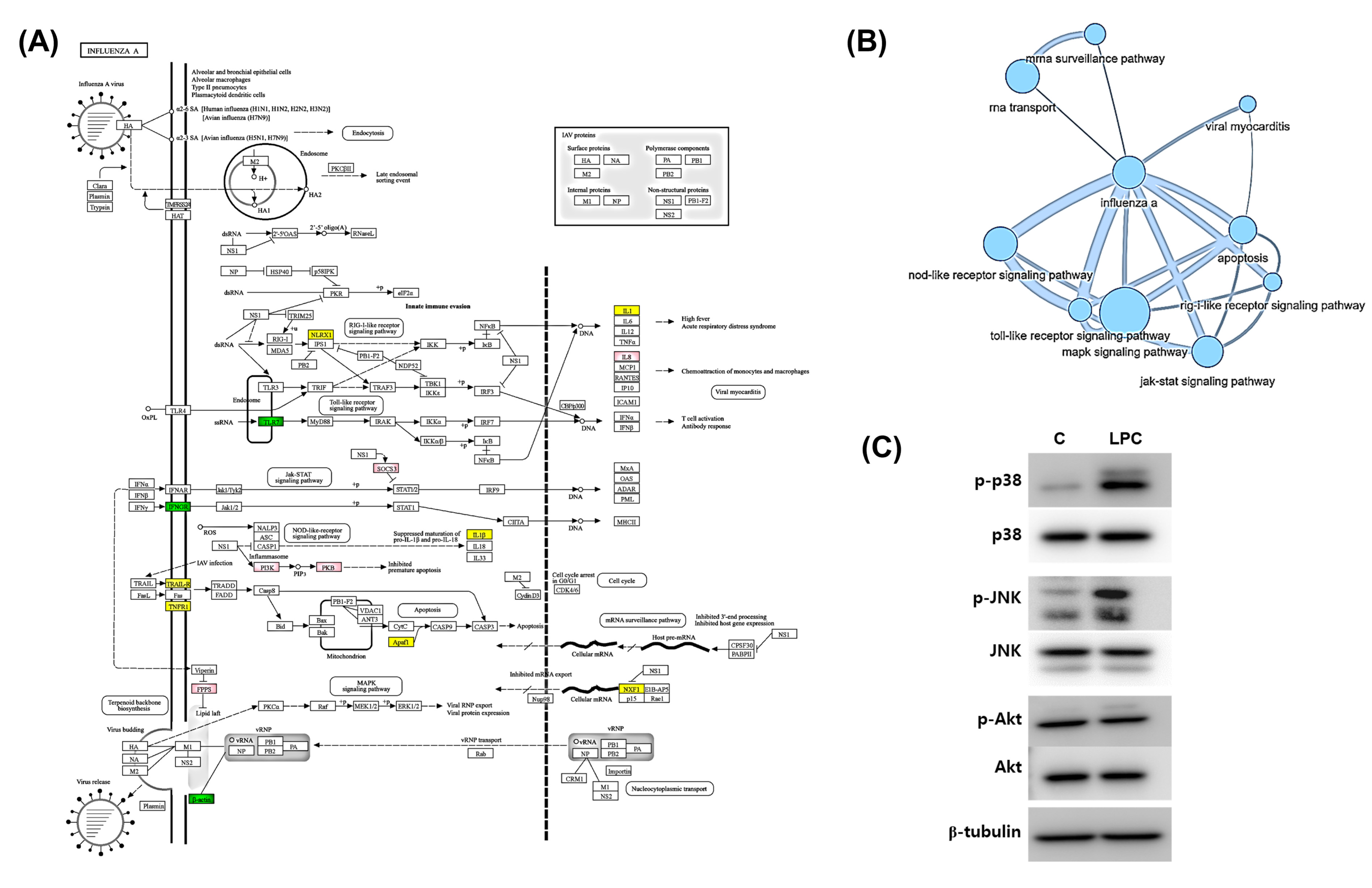
2.5. LPCs Affect Influenza A Pathway and Their Related Pathways
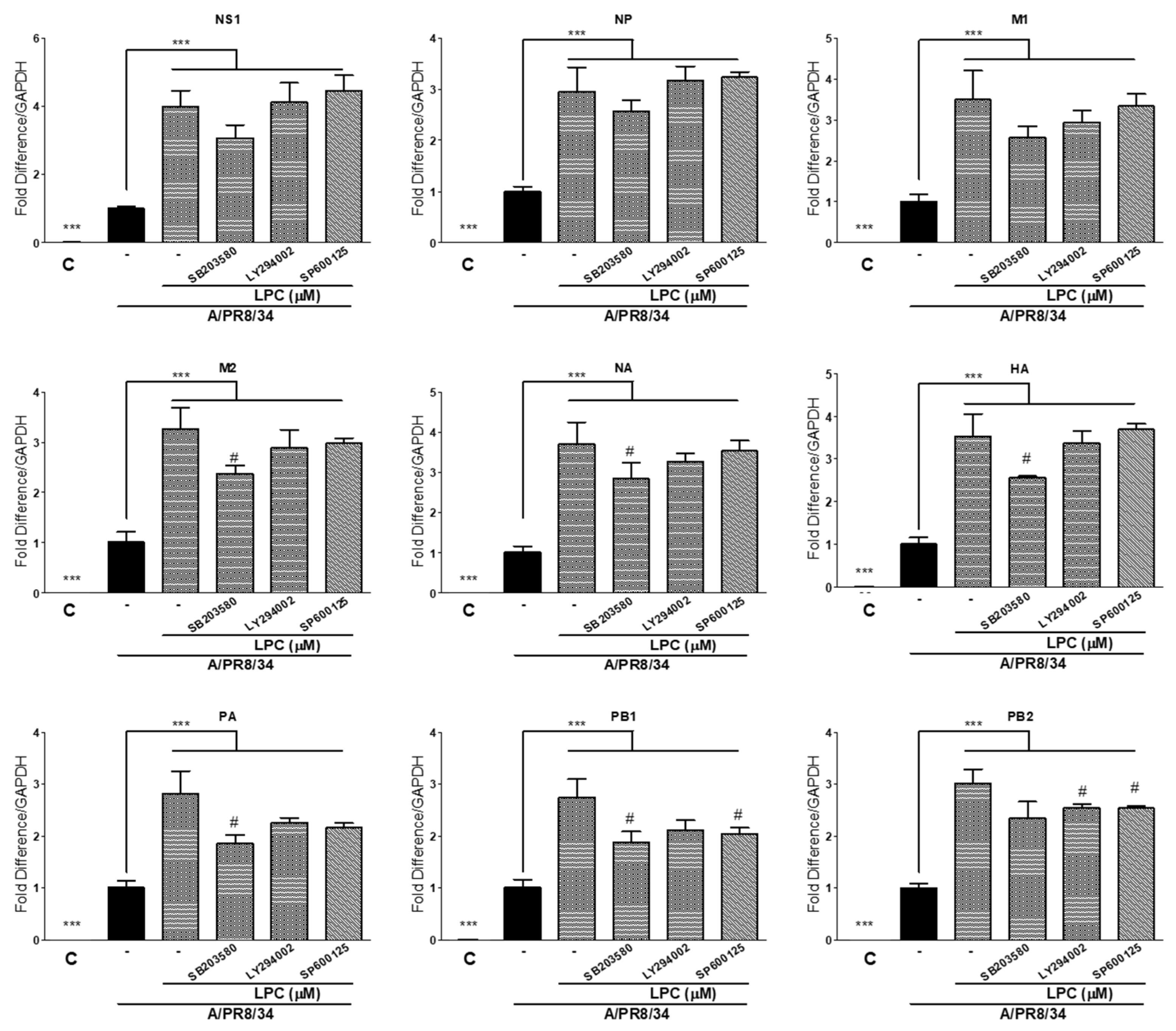
2.6. MAPK and PI3K Inhibitors Decreased Viral RNA Levels
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Virus Infection
4.3. Cytotoxicity Test
4.4. Virus Titration from Cell Supernatant
4.5. Quantitative PCR (qPCR)
4.6. Western Blot
4.7. Prediction of Putative Transcription Factor Binding Sites
4.8. RNA Profile Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gelpi, M.; Afzal, S.; Lundgren, J.; Ronit, A.; Roen, A.; Mocroft, A.; Gerstoft, J.; Lebech, A.M.; Lindegaard, B.; Kofoed, K.F.; et al. Higher Risk of Abdominal Obesity, Elevated Low-Density Lipoprotein Cholesterol, and Hypertriglyceridemia, but not of Hypertension, in People Living with Human Immunodeficiency Virus (HIV): Results From the Copenhagen Comorbidity in HIV Infection Study. Clin. Infect. Dis. 2018, 67, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Gradisteanu Pircalabioru, G.; Liaw, J.; Gundogdu, O.; Corcionivoschi, N.; Ilie, I.; Oprea, L.; Musat, M.; Chifiriuc, M.C. Effects of the Lipid Profile, Type 2 Diabetes and Medication on the Metabolic Syndrome-Associated Gut Microbiome. Int. J. Mol. Sci. 2022, 23, 7509. [Google Scholar] [CrossRef] [PubMed]
- Matveeva, S.A. Relationships between serum lipid composition and acetylator status as markers of energy metabolism in women with coronary heart disease, metabolic syndrome, and type 2 diabetes mellitus. Ter. Arkh. 2013, 85, 14–20. [Google Scholar] [PubMed]
- Zanetti, M.; Bosutti, A.; Ferreira, C.; Vinci, P.; Biolo, G.; Fonda, M.; Valente, M.; Cattin, L.; Guarnieri, G.; Barazzoni, R. Circulating pentraxin 3 levels are higher in metabolic syndrome with subclinical atherosclerosis: Evidence for association with atherogenic lipid profile. Clin. Exp. Med. 2009, 9, 243–248. [Google Scholar] [CrossRef]
- Abumweis, S.; Alrefai, W.; Alzoughool, F. Association of obesity with COVID-19 diseases severity and mortality: A meta-analysis of studies. Obes. Med. 2022, 33, 100431. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Wu, D.; Li, L.; Ding, S.; Meydani, S.N. Obesity, rather than high fat diet, exacerbates the outcome of influenza virus infection in influenza-sensitized mice. Front. Nutr. 2022, 9, 1018831. [Google Scholar] [CrossRef] [PubMed]
- Dessie, Z.G.; Zewotir, T. Mortality-related risk factors of COVID-19: A systematic review and meta-analysis of 42 studies and 423,117 patients. BMC Infect Dis. 2021, 21, 855. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.J.; Rebeles, J.; Dhungana, S.; Stewart, D.A.; Sumner, S.C.; Meyers, M.H.; Mancuso, P.; Beck, M.A. Obesity Increases Mortality and Modulates the Lung Metabolome during Pandemic H1N1 Influenza Virus Infection in Mice. J. Immunol. 2015, 194, 4846–4859. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Gu, H. The anti-obesity effects of rhein on improving insulin resistance (IR) and blood lipid levels are involved in endoplasmic reticulum stress (ERs), inflammation, and oxidative stress in vivo and vitro. Bioengineered 2021, 12, 5797–5813. [Google Scholar] [CrossRef]
- Salazar, M.R.; Carbajal, H.A.; Espeche, W.G.; Aizpurua, M.; Marillet, A.G.; Leiva Sisnieguez, C.E.; Leiva Sisnieguez, B.C.; Stavile, R.N.; March, C.E.; Reaven, G.M. Use of the triglyceride/high-density lipoprotein cholesterol ratio to identify cardiometabolic risk: Impact of obesity? J. Investig. Med. 2017, 65, 323–327. [Google Scholar] [CrossRef]
- Zhou, F.; Sun, X.; Liu, J.; Li, L.; Li, L.; Li, P. Triglyceride to high-density lipoprotein cholesterol ratio in adolescence as a predictive marker of metabolic syndrome and obesity in early adulthood in China. Endocrine 2022, 76, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Wang, X.; Li, K.; Yu, K.; Yang, L. Lipidomic profiling reveals distinct differences in plasma lipid composition in overweight or obese adolescent students. BMC Endocr. Disord. 2021, 21, 201. [Google Scholar] [CrossRef]
- Bellot, P.; Moia, M.N.; Reis, B.Z.; Pedrosa, L.F.C.; Tasic, L.; Barbosa, F., Jr.; Sena-Evangelista, K.C.M. Are Phosphatidylcholine and Lysophosphatidylcholine Body Levels Potentially Reliable Biomarkers in Obesity? A Review of Human Studies. Mol. Nutr. Food Res. 2023, 67, e2200568. [Google Scholar] [CrossRef]
- Heimerl, S.; Fischer, M.; Baessler, A.; Liebisch, G.; Sigruener, A.; Wallner, S.; Schmitz, G. Alterations of plasma lysophosphatidylcholine species in obesity and weight loss. PLoS ONE 2014, 9, e111348. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Horimoto, T.; Kawaoka, Y. Classification and genome structure of influenza virus. Nihon Rinsho 2003, 61, 1886–1891. [Google Scholar] [PubMed]
- British Infection Society; British Thoracic Society; Health Protection Agency in collaboration with the Department of Health. Pandemic flu. Clinical management of patients with an influenza-like illness during an influenza pandemic. J. Infect. 2006, 53 (Suppl. S1), S1–S58. [CrossRef]
- Wei, K.; Chen, Y.; Xie, D. Genome-scale evolution and phylodynamics of H5N1 influenza virus in China during 1996–2012. Vet. Microbiol. 2013, 167, 383–393. [Google Scholar] [CrossRef]
- Renzette, N.; Caffrey, D.R.; Zeldovich, K.B.; Liu, P.; Gallagher, G.R.; Aiello, D.; Porter, A.J.; Kurt-Jones, E.A.; Bolon, D.N.; Poh, Y.P.; et al. Evolution of the influenza A virus genome during development of oseltamivir resistance in vitro. J. Virol. 2014, 88, 272–281. [Google Scholar] [CrossRef]
- Nencioni, L.; Sgarbanti, R.; De Chiara, G.; Garaci, E.; Palamara, A.T. Influenza virus and redox mediated cell signaling: A complex network of virus/host interaction. New Microbiol. 2007, 30, 367–375. [Google Scholar]
- Overeem, N.J.; van der Vries, E.; Huskens, J. A Dynamic, Supramolecular View on the Multivalent Interaction between Influenza Virus and Host Cell. Small 2021, 17, e2007214. [Google Scholar] [CrossRef]
- Chlanda, P.; Zimmerberg, J. Protein-lipid interactions critical to replication of the influenza A virus. FEBS Lett. 2016, 590, 1940–1954. [Google Scholar] [CrossRef]
- Limsuwat, N.; Boonarkart, C.; Phakaratsakul, S.; Suptawiwat, O.; Auewarakul, P. Influence of cellular lipid content on influenza A virus replication. Arch. Virol. 2020, 165, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Almabrouk, T.A.; Ewart, M.A.; Salt, I.P.; Kennedy, S. Perivascular fat, AMP-activated protein kinase and vascular diseases. Br. J. Pharmacol. 2014, 171, 595–617. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Gang, X.; He, G.; Li, Z.; Lv, Y.; Han, Q.; Wang, G. Obesity Increases the Severity and Mortality of Influenza and COVID-19: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2020, 11, 595109. [Google Scholar] [CrossRef] [PubMed]
- Neidich, S.D.; Green, W.D.; Rebeles, J.; Karlsson, E.A.; Schultz-Cherry, S.; Noah, T.L.; Chakladar, S.; Hudgens, M.G.; Weir, S.S.; Beck, M.A. Increased risk of influenza among vaccinated adults who are obese. Int. J. Obes. 2017, 41, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.J.; Lee, D.K.; Lee, S.Y.; Sohn, S.H.; Park, H.L.; Park, Y.W.; Kim, H.; Nam, J.H. Diet-induced obesity reduces the production of influenza vaccine-induced antibodies via impaired macrophage function. Acta Virol. 2016, 60, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Gaur, S.N.; Arora, N. Lysophosphatidylcholine plays critical role in allergic airway disease manifestation. Sci. Rep. 2016, 6, 27430. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Liao, L.; Zeng, Q.; Liao, Z.; Liu, Y.; Jin, C.; Zhu, G.; Chen, C.; Yang, M.; Ai, Z.; et al. Study on the new anti-atherosclerosis activity of different Herba patriniae through down-regulating lysophosphatidylcholine of the glycerophospholipid metabolism pathway. Phytomedicine 2022, 94, 153833. [Google Scholar] [CrossRef] [PubMed]
- Yoder, M.; Zhuge, Y.; Yuan, Y.; Holian, O.; Kuo, S.; van Breemen, R.; Thomas, L.L.; Lum, H. Bioactive lysophosphatidylcholine 16:0 and 18:0 are elevated in lungs of asthmatic subjects. Allergy Asthma Immunol. Res. 2014, 6, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.H.; Lee, S.M.; Jung, J. Lysophosphatidylcholine induces expression of genes involved in cholesterol biosynthesis in THP-1 derived macrophages. Steroids 2018, 139, 28–34. [Google Scholar] [CrossRef]
- Shin, Y.K.; Li, Y.; Liu, Q.; Anderson, D.H.; Babiuk, L.A.; Zhou, Y. SH3 binding motif 1 in influenza A virus NS1 protein is essential for PI3K/Akt signaling pathway activation. J. Virol. 2007, 81, 12730–12739. [Google Scholar] [CrossRef]
- Jaworska, J.; Coulombe, F.; Downey, J.; Tzelepis, F.; Shalaby, K.; Tattoli, I.; Berube, J.; Rousseau, S.; Martin, J.G.; Girardin, S.E.; et al. NLRX1 prevents mitochondrial induced apoptosis and enhances macrophage antiviral immunity by interacting with influenza virus PB1-F2 protein. Proc. Natl. Acad. Sci. USA 2014, 111, E2110–E2119. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, M.; Cheng, A.; Jia, R.; Zhu, D.; Liu, M.; Chen, S.; Zhao, X.; Yang, Q.; Wu, Y.; et al. The role of SOCS proteins in the development of virus-induced hepatocellular carcinoma. Virol. J. 2021, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Jing, Q.; Xin, S.M.; Zhang, W.B.; Wang, P.; Qin, Y.W.; Pei, G. Lysophosphatidylcholine activates p38 and p42/44 mitogen-activated protein kinases in monocytic THP-1 cells, but only p38 activation is involved in its stimulated chemotaxis. Circ. Res. 2000, 87, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Heo, J.; Yi, C.M.; Ban, J.; Lee, N.J.; Lee, N.R.; Kim, S.W.; Kim, N.J.; Inn, K.S. A novel p38 mitogen activated protein kinase (MAPK) specific inhibitor suppresses respiratory syncytial virus and influenza A virus replication by inhibiting virus-induced p38 MAPK activation. Biochem. Biophys. Res. Commun. 2016, 477, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Hao, F.; Xu, X.; Chisolm, G.M.; Cui, M.Z. Lysophosphatidylcholine activates a novel PKD2-mediated signaling pathway that controls monocyte migration. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Smyrlaki, I.; Ekman, M.; Lentini, A.; Rufino de Sousa, N.; Papanicolaou, N.; Vondracek, M.; Aarum, J.; Safari, H.; Muradrasoli, S.; Rothfuchs, A.G.; et al. Massive and rapid COVID-19 testing is feasible by extraction-free SARS-CoV-2 RT-PCR. Nat. Commun. 2020, 11, 4812. [Google Scholar] [CrossRef]
- Minadakis, G.; Christodoulou, K.; Tsouloupas, G.; Spyrou, G.M. PathIN: An integrated tool for the visualization of pathway interaction networks. Comput. Struct. Biotechnol. J. 2023, 21, 378–387. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, M.-H.; Choi, H.-J.; Ma, J.-Y. Lysophosphatidylcholines Promote Influenza Virus Reproduction through the MAPK/JNK Pathway in PMA-Differentiated THP-1 Macrophages. Int. J. Mol. Sci. 2024, 25, 6538. https://doi.org/10.3390/ijms25126538
Cha M-H, Choi H-J, Ma J-Y. Lysophosphatidylcholines Promote Influenza Virus Reproduction through the MAPK/JNK Pathway in PMA-Differentiated THP-1 Macrophages. International Journal of Molecular Sciences. 2024; 25(12):6538. https://doi.org/10.3390/ijms25126538
Chicago/Turabian StyleCha, Min-Ho, Hee-Jeong Choi, and Jin-Yeul Ma. 2024. "Lysophosphatidylcholines Promote Influenza Virus Reproduction through the MAPK/JNK Pathway in PMA-Differentiated THP-1 Macrophages" International Journal of Molecular Sciences 25, no. 12: 6538. https://doi.org/10.3390/ijms25126538
APA StyleCha, M.-H., Choi, H.-J., & Ma, J.-Y. (2024). Lysophosphatidylcholines Promote Influenza Virus Reproduction through the MAPK/JNK Pathway in PMA-Differentiated THP-1 Macrophages. International Journal of Molecular Sciences, 25(12), 6538. https://doi.org/10.3390/ijms25126538