
Versatile Self-Assembly of Triblock Peptides into Stable Collagen Mimetic Heterotrimers

 , and
, and
Abstract
1. Introduction
2. Results
2.1. Rational Design of Triblock Peptides
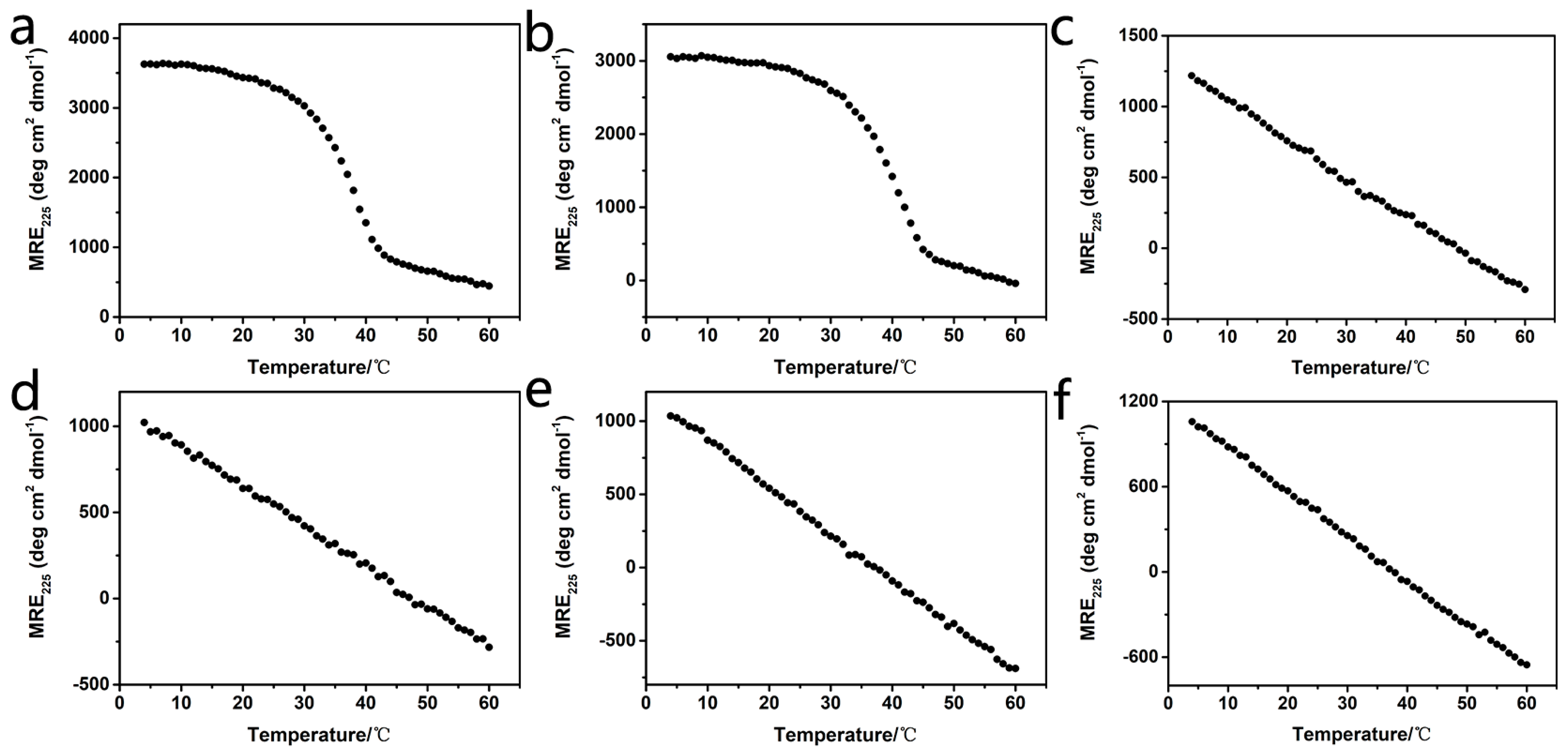
2.2. Stability of Homotrimers
2.3. Stability of Heterotrimers
2.4. Mimicking the Natural Composition of Type I Collagen
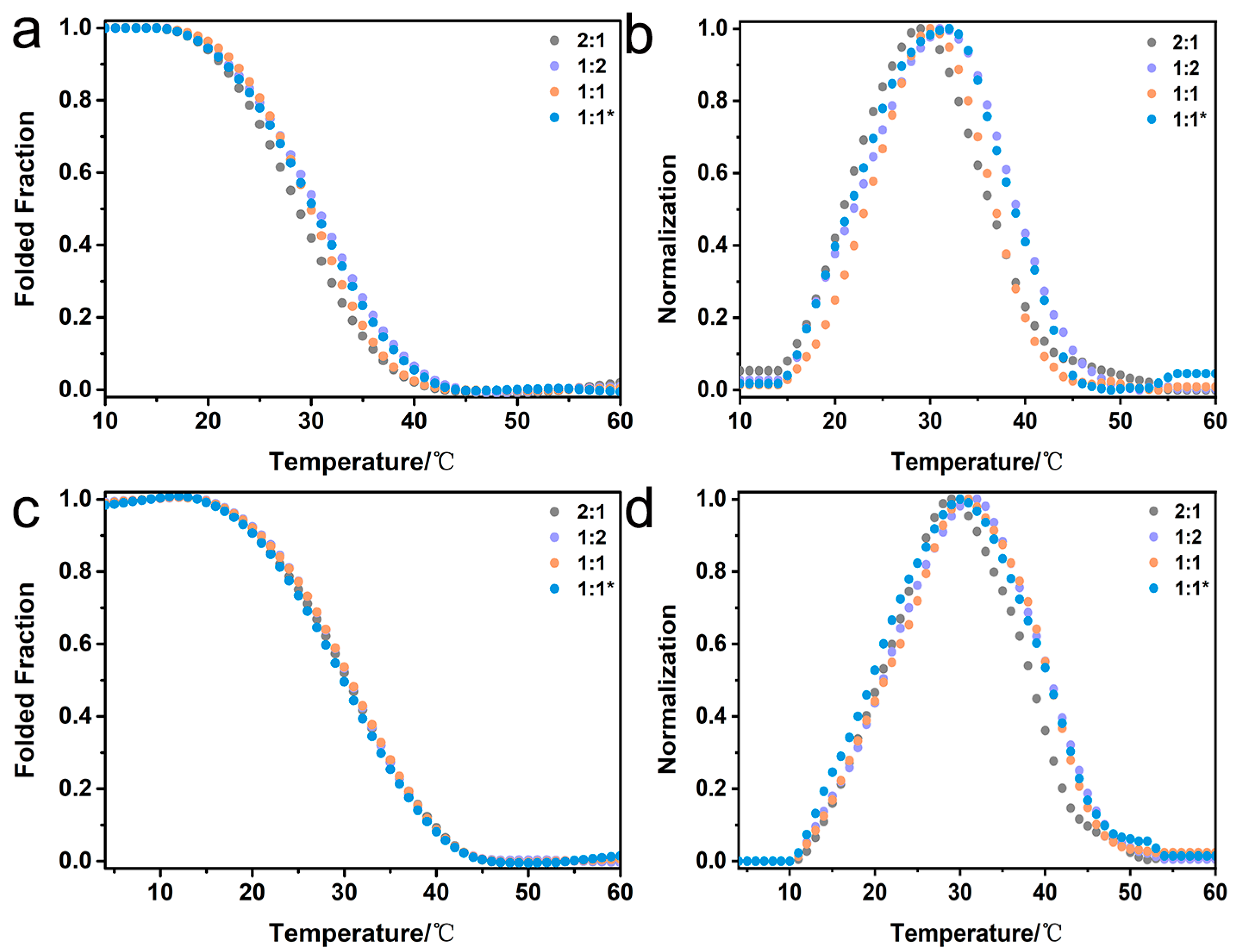
2.5. Composition Analysis of Collagen Mimetic Heterotrimers
2.6. Structural Features of Collagen Mimetic Heterotrimers
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Circular Dichroism Spectroscopy
4.3. Solution-State Nuclear Magnetic Resonance Spectroscopy
4.4. Molecular Dynamics Simulation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farhadi, S.A.; Restuccia, A.; Sorrentino, A.; Cruz-Sanchez, A.; Hudalla, G.A. Heterogeneous protein co-assemblies with tunable functional domain stoichiometry. Mol. Syst. Des. Eng. 2022, 7, 44–57. [Google Scholar] [CrossRef]
- Rudashevskaya, E.L.; Sacco, R.; Kratochwill, K.; Huber, M.L.; Gstaiger, M.; Superti-Furga, G.; Bennett, K.L. A method to resolve the composition of heterogeneous affinity-purified protein complexes assembled around a common protein by chemical cross-linking, gel electrophoresis and mass spectrometry. Nat. Protoc. 2013, 8, 75–97. [Google Scholar] [CrossRef] [PubMed]
- El Andaloussi, S.; Mager, I.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Ottl, J.; Gabriel, D.; Murphy, G.; Knäuper, V.; Tominaga, Y.; Nagase, H.; Kröger, M.; Tschesche, H.; Bode, W.; Moroder, L. Recognition and catabolism of synthetic heterotrimeric collagen peptides by matrix metalloproteinases. Chem. Biol. 2000, 7, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Renner, C.; Sacca, B.; Moroder, L. Synthetic heterotrimeric collagen peptides as mimics of cell adhesion sites of the basement membrane. Biopolymers 2004, 76, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Sacca, B.; Fiori, S.; Moroder, L. Studies of the local conformational properties of the cell-adhesion domain of collagen type IV in synthetic heterotrimeric peptides. Biochemistry 2003, 42, 3429–3436. [Google Scholar] [CrossRef] [PubMed]
- Sacca, B.; Moroder, L. Synthesis of heterotrimeric collagen peptides containing the alpha1beta1 integrin recognition site of collagen type IV. J. Pept. Sci. 2002, 8, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Sacca, B.; Renner, C.; Moroder, L. The chain register in heterotrimeric collagen peptides affects triple helix stability and folding kinetics. J. Mol. Biol. 2002, 324, 309–318. [Google Scholar] [CrossRef]
- Bella, J. Collagen structure: New tricks from a very old dog. Biochem. J. 2016, 473, 1001–1025. [Google Scholar] [CrossRef]
- Yang, L.; van der Werf, K.O.; Fitie, C.F.; Bennink, M.L.; Dijkstra, P.J.; Feijen, J. Mechanical properties of native and cross-linked type I collagen fibrils. Biophys. J. 2008, 94, 2204–2211. [Google Scholar] [CrossRef] [PubMed]
- Keene, D.R.; San Antonio, J.D.; Mayne, R.; McQuillan, D.J.; Sarris, G.; Santoro, S.A.; Iozzo, R.V. Decorin binds near the C terminus of type I collagen. J. Biol. Chem. 2000, 275, 21801–21804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.M.; Kapyla, J.; Puranen, J.S.; Knight, C.G.; Tiger, C.F.; Pentikainen, O.T.; Johnson, M.S.; Farndale, R.W.; Heino, J.; Gullberg, D. alpha 11beta 1 integrin recognizes the GFOGER sequence in interstitial collagens. J. Biol. Chem. 2003, 278, 7270–7277. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, X.; Niyibizi, C. Distribution of single-cell expanded marrow derived progenitors in a developing mouse model of osteogenesis imperfecta following systemic transplantation. Stem. Cells 2007, 25, 3183–3193. [Google Scholar] [CrossRef]
- Castillo-Briceno, P.; Bihan, D.; Nilges, M.; Hamaia, S.; Meseguer, J.; Garcia-Ayala, A.; Farndale, R.W.; Mulero, V. A role for specific collagen motifs during wound healing and inflammatory response of fibroblasts in the teleost fish gilthead seabream. Mol. Immunol. 2011, 48, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development. Adv. Mater. 2019, 31, e1801651. [Google Scholar] [CrossRef] [PubMed]
- Bella, J.; Hulmes, D.J. Fibrillar Collagens. Subcell. Biochem. 2017, 82, 457–490. [Google Scholar] [PubMed]
- Fiori, S.; Saccà, B.; Moroder, L. Structural properties of a collagenous heterotrimer that mimics the collagenase cleavage site of collagen type I. J. Mol. Biol. 2002, 319, 1235–1242. [Google Scholar] [CrossRef]
- Jalan, A.A.; Sammon, D.; Hartgerink, J.D.; Brear, P.; Stott, K.; Hamaia, S.W.; Hunter, E.J.; Walker, D.R.; Leitinger, B.; Farndale, R.W. Chain alignment of collagen I deciphered using computationally designed heterotrimers. Nat. Chem. Biol. 2020, 16, 423–429. [Google Scholar] [CrossRef]
- Luo, T.; Kiick, K.L. Collagen-Like Peptide Bioconjugates. Bioconjug. Chem. 2017, 28, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Strauss, K.; Chmielewski, J. Advances in the design and higher-order assembly of collagen mimetic peptides for regenerative medicine. Curr. Opin. Biotechnol. 2017, 46, 34–41. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, L.E.; Fallas, J.A.; Hartgerink, J.D. Positive and negative design leads to compositional control in AAB collagen heterotrimers. J. Am. Chem. Soc. 2011, 133, 5432–5443. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Lu, C.; Lan, J.; Fan, S.; Nanda, V.; Xu, F. How electrostatic networks modulate specificity and stability of collagen. Proc. Natl. Acad. Sci. USA 2018, 115, 6207–6212. [Google Scholar] [CrossRef] [PubMed]
- Hodges, J.A.; Raines, R.T. Stereoelectronic and steric effects in the collagen triple helix: Toward a code for strand association. J. Am. Chem. Soc. 2005, 127, 15923–15932. [Google Scholar] [CrossRef] [PubMed]
- Hentzen, N.B.; Islami, V.; Kohler, M.; Zenobi, R.; Wennemers, H. A Lateral Salt Bridge for the Specific Assembly of an ABC-Type Collagen Heterotrimer. J. Am. Chem. Soc. 2020, 142, 2208–2212. [Google Scholar] [CrossRef] [PubMed]
- Gauba, V.; Hartgerink, J.D. Self-assembled heterotrimeric collagen triple helices directed through electrostatic interactions. J. Am. Chem. Soc. 2007, 129, 2683–2690. [Google Scholar] [CrossRef] [PubMed]
- Jalan, A.A.; Demeler, B.; Hartgerink, J.D. Hydroxyproline-Free Single Composition ABC Collagen Heterotrimer. J. Am. Chem. Soc. 2013, 135, 6014–6017. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.G.; Morton, L.F.; Onley, D.J.; Peachey, A.R.; Messent, A.J.; Smethurst, P.A.; Tuckwell, D.S.; Farndale, R.W.; Barnes, M.J. Identification in collagen type I of an integrin alpha2 beta1-binding site containing an essential GER sequence. J. Biol. Chem. 1998, 273, 33287–33294. [Google Scholar] [CrossRef]
- Malcor, J.D.; Bax, D.; Hamaia, S.W.; Davidenko, N.; Best, S.M.; Cameron, R.E.; Farndale, R.W.; Bihan, D. The synthesis and coupling of photoreactive collagen-based peptides to restore integrin reactivity to an inert substrate, chemically-crosslinked collagen. Biomaterials 2016, 85, 65–77. [Google Scholar] [CrossRef]
- Siljander, P.R.; Hamaia, S.; Peachey, A.R.; Slatter, D.A.; Smethurst, P.A.; Ouwehand, W.H.; Knight, C.G.; Farndale, R.W. Integrin activation state determines selectivity for novel recognition sites in fibrillar collagens. J. Biol. Chem. 2004, 279, 47763–47772. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, W.; Yu, J.; Luo, L.; Wang, J.; Xiao, J. Ln3+-Triggered self-assembly of a heterotrimer collagen mimetic peptide into luminescent nanofibers. Chem. Commun. 2020, 56, 15141–15144. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.R.; Soliakov, A.; Lewis, R.J. In vitro fibrillogenesis of collagen type I in varying ionic and pH conditions. Micron 2013, 49, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; He, M.; Wang, L.; Luo, L.; Wang, J.; Xiao, J. Luminescent Biofunctional Collagen Mimetic Nanofibers. ACS Omega 2019, 4, 16270–16279. [Google Scholar] [CrossRef]
- Rele, S.; Song, Y.; Apkarian, R.P.; Qu, Z.; Conticello, V.P.; Chaikof, E.L. D-periodic collagen-mimetic microfibers. J. Am. Chem. Soc. 2007, 129, 14780–14787. [Google Scholar] [CrossRef]
- O’Leary, L.E.; Fallas, J.A.; Bakota, E.L.; Kang, M.K.; Hartgerink, J.D. Multi-hierarchical self-assembly of a collagen mimetic peptide from triple helix to nanofibre and hydrogel. Nat. Chem. 2011, 3, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Rainey, J.K.; Goh, M.C. An interactive triple-helical collagen builder. Bioinformatics 2004, 20, 2458–2459. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle Mesh Ewald: An N·Log(N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Name | Peptide Sequence | Tm/°C |
|---|---|---|
| α1 | (GPO)3GFOGER(GPO)3 | 38.0 |
| α2 | (GPO)3GPOGES(GPO)3 | 41.0 |
| K-α1-D | K5(GPO)3GFOGER(GPO)3D5 | <4.0 |
| D-α1-K | D5(GPO)3GFOGER(GPO)3K5 | <4.0 |
| K-α2-D | K5(GPO)3GPOGES(GPO)3D5 | <4.0 |
| D-α2-K | D5(GPO)3GPOGES(GPO)3K5 | <4.0 |
| K-α1*-D | K5(GPO)3GFOG*ER(GPO)3D5 | - |
| D-α2*-K | D5(GPO)3GPOG*ES(GPO)3K5 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, L.; Ling, B.; Zhao, S.; Yu, F.; Liu, H.; Wang, S.; Xiao, J. Versatile Self-Assembly of Triblock Peptides into Stable Collagen Mimetic Heterotrimers. Int. J. Mol. Sci. 2024, 25, 6550. https://doi.org/10.3390/ijms25126550
Yao L, Ling B, Zhao S, Yu F, Liu H, Wang S, Xiao J. Versatile Self-Assembly of Triblock Peptides into Stable Collagen Mimetic Heterotrimers. International Journal of Molecular Sciences. 2024; 25(12):6550. https://doi.org/10.3390/ijms25126550
Chicago/Turabian StyleYao, Linyan, Biyang Ling, Sha Zhao, Fansen Yu, Huanxiang Liu, Shenlin Wang, and Jianxi Xiao. 2024. "Versatile Self-Assembly of Triblock Peptides into Stable Collagen Mimetic Heterotrimers" International Journal of Molecular Sciences 25, no. 12: 6550. https://doi.org/10.3390/ijms25126550
APA StyleYao, L., Ling, B., Zhao, S., Yu, F., Liu, H., Wang, S., & Xiao, J. (2024). Versatile Self-Assembly of Triblock Peptides into Stable Collagen Mimetic Heterotrimers. International Journal of Molecular Sciences, 25(12), 6550. https://doi.org/10.3390/ijms25126550