Abstract
MicroRNAs (miRNAs) are small non-coding RNAs that play an important role by controlling gene expression in the cytoplasm in almost all biological pathways. Recently, scientists discovered that miRNAs are also found within mitochondria, the energy-producing organelles of cells. These mitochondrial miRNAs, known as mitomiRs, can originate from the nuclear or mitochondrial genome, and they are pivotal in controlling mitochondrial function and metabolism. New insights indicate that mitomiRs may influence key aspects of the onset and progression of cardiovascular disease, especially concerning mitochondrial function and metabolic regulation. While the importance of mitochondria in cardiovascular health and disease is well-established, our understanding of mitomiRs’ specific functions in crucial biological pathways, including energy metabolism, oxidative stress, inflammation, and cell death, is still in its early stages. Through this review, we aimed to delve into the mechanisms of mitomiR generation and their impacts on mitochondrial metabolic pathways within the context of vascular cell aging and atherosclerotic cardiovascular disease. The relatively unexplored field of mitomiR biology holds promise for future research investigations, with the potential to yield novel diagnostic tools and therapeutic interventions.
1. Introduction
Atherosclerotic cardiovascular disease (ACVD) is the principal cause of morbidity and mortality worldwide, with myocardial infarction (MI) being the most severe form [1]. Atherosclerosis, which is induced by the accumulation of lipids and fibrous elements in the arteries, serves as the primary factor for ACVD. It manifests as an inflammatory disease characterized by the formation of atherosclerotic plaque [2].
Atherosclerosis has a complex pathogenesis and numerous risk factors, such as diabetes, hypertension, low density lipoprotein cholesterol, and smoking. Aging is also a key element for atherosclerosis and perseveres as an independent contributor, even after controlling for other factors [3]. Age-related changes in the vasculature not only promote a proatherogenic environment but also impact the composition and vulnerability of atherosclerotic plaques [4].
Despite extensive research, many mechanisms driving the progression of atherosclerosis remain elusive. Thus, delving into the molecular mechanisms of ACVD is crucial, alongside identifying novel markers for early and effective disease prevention.
With the advent of personalized medicine and next-generation sequencing technology, microRNAs (miRNAs) have garnered significant attention. There is mounting evidence suggesting that miRNAs, being endogenous, stable, short, non-coding RNAs, could serve as diagnostic and prognostic markers and potential therapeutic targets for several CVDs [5].
miRNAs serve as crucial regulators of gene expression at the post-transcriptional level. They bind to complementary sequences within the 3′UTR of target genes, leading to the degradation or translation inhibition of mRNA [6]. miRNAs are expressed in the cardiovascular system and have arisen as important regulators of cardiovascular physiology and pathophysiology [5]. They exert regulatory functions in the gene and protein expression changes observed during atherogenesis, with miRNA-related molecular mechanisms driving the initiation and progression of atherosclerosis [7,8]. Furthermore, miRNAs were detected in circulation, either complexed with AGO2 or encapsulated within exosomes. Their abundance in disease contexts suggests promising potential as biomarkers for disease onset and progression [9].
Mature miRNAs are normally present in the cytoplasm of cells. Interestingly, several studies disclosed the presence of miRNAs, called mitochondrial miRNAs (mitomiRs), within the mitochondrion [10,11,12]. The mechanism underlying the translocation of these nuclear-encoded miRNAs into the mitochondria remains unclear. Most of them originate from nucleus, but some mitomiRs may have a mitochondrial origin, serving as regulators of gene expression within the mitochondria [13,14]. They are localized in mitochondria in different cells and have different thermodynamic properties compared with miRNAs [15,16].
mitomiRs could influence the translational activity at the mitochondrial genome, thereby playing critical roles in mitochondrial function and metabolic regulation [15]. Of importance, emerging evidence suggests that mitomiRs may be novel contributors to CVD, underscoring the potential of mitomiR signatures as molecular markers of disease [17,18,19].
This review aimed to provide a comprehensive overview of available data on mitomiR biosynthesis mechanisms and their relevance in vascular cell senescence and the pathogenesis of MI, while also highlight potential future research directions.
2. Canonical miRNA Biogenesis
miRNAs are frequently encoded singly or in clusters within larger host genes. They may reside within introns, in the 3′-untranslated regions (3′-UTR), or within coding regions. Another category of miRNAs exists known as intergenic miRNAs, which are transcribed under the regulation of their promoters [17].
The biogenesis of miRNAs is tightly regulated across multiple stages and involves several enzymatic steps occurring in both the nucleus and the cytoplasm [20,21]. The initiation of miRNA biogenesis starts in the nucleus with the transcription of a 100–1000-nucleotide (nt) primary miRNA (pri-miRNA) from the DNA by the RNA polymerase II. The pri-miRNA harbors a local stem–loop structure measuring around 35 base pairs in size.
The ribonuclease III (RNase III) enzyme Drosha cleaves the flanks of pri-miRNAs, releasing a ~70-nucleotide stem–loop structure known as precursor miRNA (pre-miRNA). Pre-miRNAs contain the ~22-nucleotide mature miRNA in either the 5′ or 3′ half of their stem, and the pair of cuts made by Drosha establishes either the 5′ or the 3′ end of the mature miRNA.
Another enzyme, Pasha, also referred to as the DiGeorge syndrome chromosomal region 8 enzyme (DGCR8), plays a crucial role in assisting Drosha in processing pre-miRNAs. Pasha also regulates Drosha activity within the nuclear fraction of cells. Pasha’s two double-stranded RNA-binding domains interact with the pri-miRNA, while the C-terminal domain recruits Drosha, forming a functional microprocessor complex [22].
Following the initial processing in the nucleus, pre-miRNA undergoes further maturation within the cytoplasm. It is associated with Exportin-5, which is a nucleocytoplasmic transporter factor, and RanGTP, which shields it from nuclear degradation and assists in its cytoplasmic translocation.
Once in the cytosol, pre-miRNA is subjected to cleavage by another enzyme called Dicer, leading to the formation of a small RNA duplex. This duplex is subsequently processed into a single-stranded and linear mature miRNA [19]. The small RNA duplex generated by Dicer binds to a protein called argonaute (Ago), initiating the assembly of a complex comprising multiple proteins, such as TRBP1 and TRBP2, as well as the targeted mRNA. This complex, known as the RNA-induced silencing complex (RISC), identifies target mRNA through complementarity in the 3′-UTR. This recognition results in the degradation of the mRNA or the inhibition of its translation to a protein sequence [23].
3. Mitochondrial miRNA Biogenesis
miRNAs carry out their functions in the cytoplasm; however, there have been instances of their presence in various subcellular locations. Some miRNAs can be retroactively transported to the nucleus [24,25]. Conversely, certain miRNAs, known as mitomiRs, are imported from the cytoplasm into mitochondria to regulate mitochondrial structure and function, notably biogenesis, bioenergetics, and dynamics [26].
Barrey and colleagues were the first to demonstrate the presence of pre-miRNAs in mitochondria. They suggested that some pre-miRNA sequences could be processed into mature miRNAs that become instantly active on mitochondrial transcripts or exported to the cytoplasm to disrupt mRNA [11]. In 2011, this research group screened 742 miRNAs and found that 243 miRNAs were significantly expressed in mitochondrial RNA isolated from human myotubes [11].
mitomiRs, mainly encoded by the nuclear genome, are transported into mitochondria to influence the expression of mRNAs that originate from the mitochondrial DNA. Although the exact mechanisms facilitating the miRNA import into mitochondria are not fully understood, various proteins were found to take part in this intricate transport process.
A key protein involved is Ago2, which is present in mitochondria across different cell types. Studies indicate that miRNA-Ago2 interactions play a crucial role in the mitochondrial translocation of mitomiRs [27,28,29,30]. For instance, it was shown that miRNA-1 upregulated during myogenesis can efficiently enter mitochondria in complex with Ago2 [31], thus increasing the translation of mitochondrial-encoded transcripts [32]. Another significant player in the import of miRNAs into mitochondria is polynucleotide phosphorylase (PNPase), which is situated at the inner mitochondrial membrane and projects into the intermembrane space. Disruption of the PNPase gene leads to disturbances in the mitochondrial morphology and function, partially due to hindered RNA imports crucial for transcription and translation processes [33]. Moreover, Shepherd et al. [34] observed that PNPase overexpression in HL-1 cardiomyocytes coincides with elevated levels of miRNA-378, highlighting the role of the transport of the miRNA [34].
GW bodies also play an important role in forming structures, where the protein GW182 associates with miRNA and Ago2 in a cap-like structure to form a stable RISC [35,36]. Furthermore, porins, which are conserved proteins in the outer mitochondrial membrane, may facilitate miRNA translocation from the cytoplasm to the mitochondria [15].
There is an emerging interest in identifying mitochondrial genome-encoded miRNAs and their potential impact on mitochondrial functions. Predictions of mitochondrial genome-encoded miRNAs surfaced through comprehensive analyses of the entire mitochondrial transcriptome [12,37]. Additionally, mitochondrial deep-sequencing and small-RNA-sequencing works identified mitosRNA in humans [38]. Reports suggest the direct transcription of miRNA-1974, miRNA-1977, and miRNA-1978 from mitochondrial DNA [11,39,40,41]. Notably, certain pre-miRNAs and mature miRNAs, like premiR-let7 and pre-miR-302a found in human mitochondria, exhibit alignment with the mitochondrial genome, suggesting intramitochondrial mitomiR biogenesis [11,42,43].
Intriguingly, a recent study found 13 dysregulated mitochondrial genome-encoded mitomiRs in cancer cell lines [44]. The authors found reduced mitomiR expression with mitochondrial DNA depletion and inhibition of mitochondrial transcription. These mitomiRs interacted physically with Ago2, regulating the expression of both nuclear and mitochondrial mRNAs. Notably, miRNA-5 targets the PPARGC1A gene, influencing the mtDNA content in breast cancer cells [44].
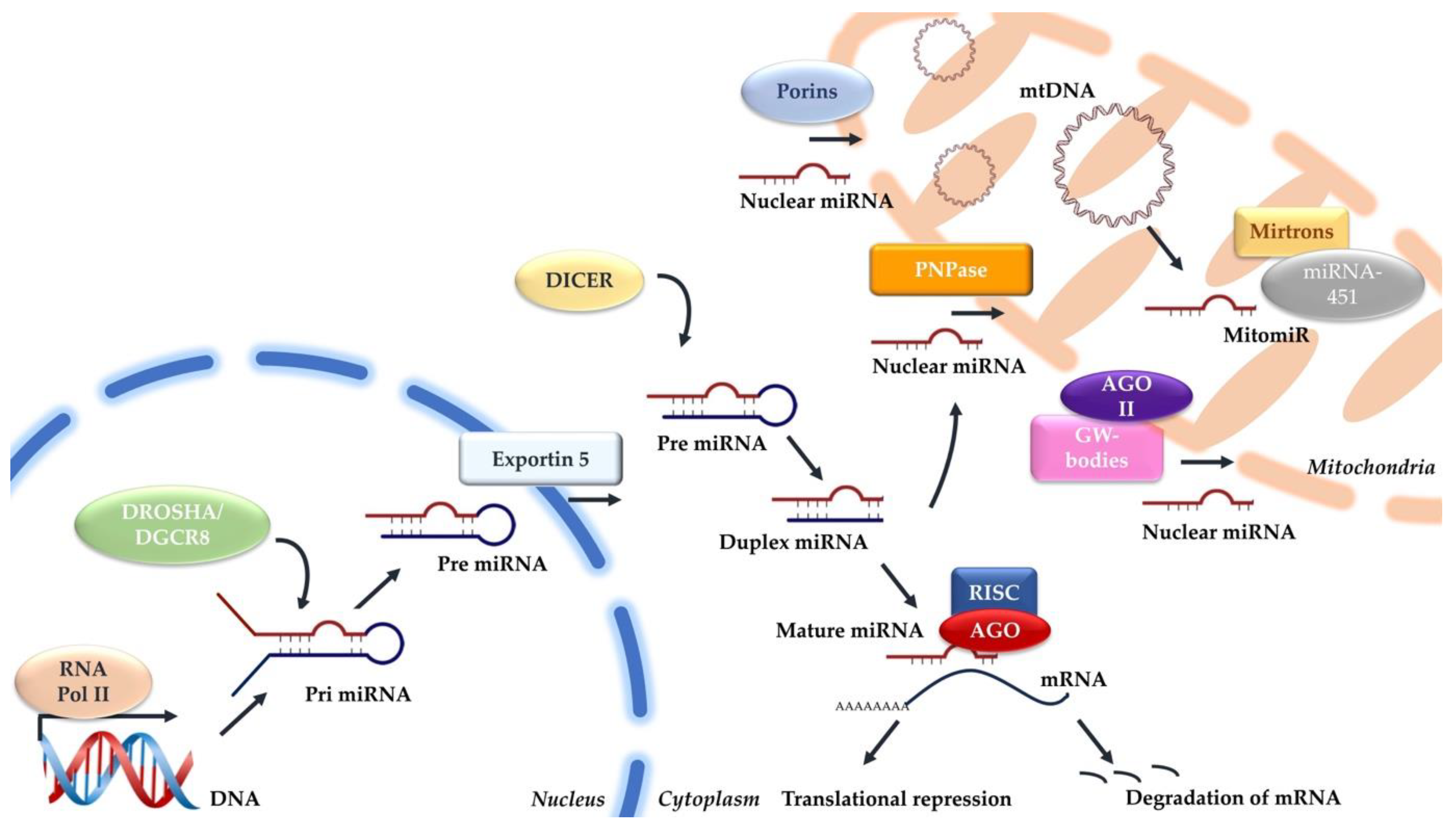
However, verifying these mitomiRs as transcribed from mitochondrial genome remains unknown. The absence of canonical miRNA processing enzymes—Drosha, DGCR8, and Dicer—within mitochondria poses a challenge in confirming the mitochondrial origin of these miRNAs [16]. To date, despite the exploration of non-canonical biogenesis pathways, such as Drosha-independent pathways like mirtrons and Dicer-independent pathways like miRNA-451, no established mechanisms have been proposed for mitochondrial mitomiR biogenesis [45]. Figure 1 shows the potential mechanisms involved in the production and import of mitomiRs.
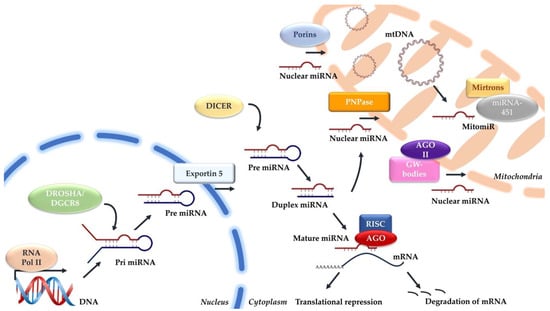
Figure 1.
Mechanisms involved in the canonical miRNA and mitochondrial miRNA biogenesis.
4. mitomiR Functions in Vascular Cell Senescence and Atherosclerosis
The process of aging encompasses a gradual decline in cellular function, marked by various indicators, such as genomic instability in both nuclear and mitochondrial DNA, epigenetic alterations, disrupted protein balance, cellular senescence, depletion of stem cells, and modified cellular interactions [46].
In recent years, scientists have increasingly recognized the significance of nuclear and mitochondrial DNA damage in vascular aging and atherogenesis [47,48]. There is substantial evidence indicating the presence of oxidative DNA damage, telomere shortening, and mitochondrial DNA damage in both experimental models and human atherosclerotic plaques [49,50,51,52], as well as in the peripheral cells of patients with atherosclerosis [53,54,55]. Moreover, it is becoming evident that genomic instability can directly impact vascular cell function, which activates signaling pathways that lead to a cascade of cellular and molecular changes, and ultimately, promotes inflammation, cell death, and cellular senescence, accompanied by the secretion of inflammatory factors known as the “senescence-associated secretory phenotype” (SASP) [56].
Nevertheless, the fine mechanisms linking DNA damage to vascular and cellular aging, as well as to the pathogenesis of atherosclerosis, are still awaiting elucidation. A growing body of evidence suggests that there exists a mutual crosstalk between telomere dysfunction and mitochondrial dysmetabolism during the process of vascular senescence [47]. This underscores the importance of comprehensively studying the molecular mediators, such as mitomiRs, involved in this complex and intricate connection [57,58,59].
Aside from their role as cellular powerhouses, mitochondria exert a key role in various cellular pathways, such as cell death, metabolic, and aging processes [60,61]. One crucial mechanism by which mitochondria contribute to aging is through reactive oxygen species (ROS) production. When the balance between ROS production and neutralization is disrupted, oxidative stress occurs, leading to premature aging and tissue damage [62,63]. Mitochondrial dysfunction is involved in cellular aging and atherogenesis [64] and is associated with the development of numerous diseases, including CVDs [18,65,66]. Changes in mitochondrial structure and function in endothelial and smooth muscle cells contribute to aging, driven by mitochondrial oxidative stress, DNA mutations, dynamics, and mitophagy [64,67].
The dysregulation of mitomiRs may disrupt mitochondrial homeostasis and function during cellular senescence [68]. For instance, miRNA-181c, which is encoded by nuclear DNA, was shown to regulate mitochondrial gene expression and function, affecting the cellular physiology [13,69]. Cardiomyocytes, which have a high density of mitochondria, rely on oxidative metabolism for continuous energy production and were instrumental in studying mitomiRs’ role in mitochondrial homeostasis [13].
Recent studies highlighted the role of miRNA-181b in regulating vascular endothelial aging through the MAP3K3-MAPK signaling process, demonstrating its importance in endothelial cell function and aging [70]. Additionally, mitomiRs, such as miRNA-146a and miRNA-34a, were observed to increase in endothelial cells undergoing replicative senescence, influencing ROS production and apoptosis sensitivity [71]. These mitomiRs play a significant role in mediating the biochemical and morphological changes associated with aging cells, affecting cellular outcomes based on the senescence status of the cell [72].
miRNA-146a is widely studied in cellular senescence and the SASP [73]. Its synthesis is closely linked to inflammatory processes, and its effects on cellular activities are highly dependent on the specific stimulus. Enriched in the mitochondrial fraction of cardiomyocytes, miRNA-146a plays a cardioprotective role by inhibiting the mitochondria-dependent apoptotic pathway and mitigating the loss of mitochondrial membrane potential [73].
miR-34a modulates endothelial senescence by the suppression of SIRT1. Its expression increases in senescent human umbilical cord vein endothelial cells (HUVECs) and in the hearts of old mice. Overexpressing miRNA-34a inhibits the expression of SIRT1, which is a key epigenetic regulator of p66Shc, which is a mitochondrial adaptor involved in ROS accumulation and mitochondrial dysfunction [74,75]. Under physiological conditions, SIRT1-induced histone deacetylation suppresses p66Shc gene expression by reducing transcription factor access to chromatin. The loss of SIRT1 balance activates harmful pathways linked to vascular aging [75].
miRNA-21, which was initially identified for its role in cancer, has emerged as a key player in inflammatory and senescence processes, influencing mitochondrial function and cellular metabolism [76]. The expression of miRNA-21 was detected to increase in both replicative and induced senescence [77,78]. miR-21 is highly enriched in the myocardium in various cardiovascular disease models, where it inhibits cardiomyocyte apoptosis by targeting the PDCD4 mRNA [79]. Additionally, the overexpression of miRNA-21 in rat cardiomyocytes leads to a decrease in mitochondrial fatty acid oxidation and mitochondrial respiration, suggesting a role in shifting cellular metabolism toward the glycolytic pathway [80]. Recent evidence confirms that miRNA-21 can translocate into mitochondria and enhance the translation of mt-Cyb in a rat model, further supporting its role in mitochondrial function regulation [81].
The disruption of certain mitomiR-mediated pathways contributes to atherosclerosis by altering mitochondrial homeostasis. 7-ketocholesterol, which is found in oxidized LDL, decreases isocitrate dehydrogenase levels via miRNA-144 overexpression, leading to reduced NADPH, increased ROS, and decreased NO, which impacts endothelial health and promotes atherosclerosis [82]. Additionally, miRNA-34a, which is induced by oxidized LDL, enhances apoptosis and ROS levels, but reducing its levels enhances mitochondrial function and cell survival, making it a potential treatment target and biomarker for atherosclerosis [83].
5. mitomiRs in Myocardial Infarction
5.1. In Vitro and In Vivo Functional Studies
Multiple miRNAs play a role in remodeling the myocardium following ischemia/reperfusion injury in an MI. Changes in miRNA expression are associated with stress-signaling pathway activation [84,85], which impacts ACVD by modulating key mitochondrial components involved in cell survival and death [86].
Elevated levels of miRNA-532, miRNA-690, miRNA-345-3p, and miRNA-696 were detected in cardiomyocyte mitochondria within damaged cardiac areas, correlating with energy metabolism and oxidative stress pathways [87]. Among these, miRNA-696 targets PGC-1α, diminishing mitochondrial biosynthesis and fatty acid oxidation [88], while miRNA-532-3p directly targets apoptosis repressor, influencing mitochondrial fission and cardiomyocyte apoptosis [89].
mitomiRs are important for their role in the regulation of the mitochondrial fragmentation. miR-19b has been identified to modulate Socs6, which is a protein involved in mitochondrial fragmentation [90]. Additionally, the reduction in the miR-26b leads to an upregulation of its target Mfn1, inhibiting excessive mitochondrial fragmentation and reducing heart damage caused by ischemia/reperfusion (I/R) injury [91]. miR-140 was shown to regulate Mfn1 levels in cardiomyocytes during I/R injury [92]. Conversely, the upregulation of miR-421 results in reduced levels of its target Pink-1, which promotes mitochondrial fragmentation and exacerbates infarction damage [93].
Some mitomiRs also exert their effects by regulating the expression of mitochondrial genes. In myocardial infarction, the expression of mitomiRs miRNA-762 and miRNA-210 is increased, whereas miRNA-1 is downregulated. miRNA-1 modulates the mitochondrial electron transport chain (ETC) by targeting mitochondrial genes, such as mt-COX1 and mt-ND1 [31]. miRNA-762 upregulation directly inhibits ND2 translation, reducing mitochondrial complex I enzyme activity and intracellular ATP levels, and increasing ROS levels and apoptosis in cardiomyocytes [94]. Additionally, the knockdown of miRNA-762 was found to mitigate myocardial ischemia/reperfusion injury in mice [95].
In cardiac tissue, miR-210, which is a hypoxia-induced miRNA, suppresses the expression of iron-sulfur cluster homolog (ISCU1/2), impairing mitochondrial respiration and ROS generation [96]. miRNA-210 also suppresses mitochondrial function by targeting the 3′UTR of mitochondrial proteins, including mitochondrial ISCU, COX10, succinate dehydrogenase complex subunit D, and complex III [94]. However, the overexpression of this mitomiR was found to reduce cell death and improved cardiac function and angiogenesis after acute myocardial infarction in vivo [97]. Therefore, the correct function of miRNA-210 in ischemic heart disease remains to be elucidated.
Certain miRNAs regulate epigenetic modulators, influencing mitochondrial function. Upregulated miRNA-195 in an ischemic heart directly binds to Sirtuin 3, downregulating its expression, leading to compromised mitochondrial respiratory activity [98]. The upregulation of the miR-15/16 family also suppresses ATP levels [99].
Conversely, miRNAs that are upregulated following cardiac stress, such as miRNA-499 and miRNA-214, show protective functions. miRNA-499 downregulates calcineurin and Drp-1, which are both involved in mitochondrial fission, thereby reducing cardiomyocyte apoptosis and infarct size. Its inhibition produces the opposite effect [100]. Myocardial infarction also increases the miRNA-214 level, which enhances protection during ischemia by reducing calcium overload and promoting cardiomyocyte survival through the inhibition of the mitochondrial permeability transition pore and pro-apoptotic proteins [101]. Other signaling pathways regulating mitochondrial fission include miRNA-761/Mff [102] and miRNA-484/Fis1 [103]. Table 1 summarizes the mitomiRs and their impacts on the mitochondrial processes in a myocardial infarction and related conditions.

Table 1.
List of mitomiRs and their impact on mitochondrial processes in myocardial infarction.
5.2. mitomiRs as Circulating Biomarkers
Changes in the circulating levels of miRNA-1, miRNA-21, and miRNA-499 have been reported in patients with an acute myocardial infarction, highlighting the role of mitomiRs as diagnostic markers of disease [5]. miRNA-1 was correlated with the myocardial infarct size [104]. miRNA-1 was also associated with the absolute change in infarct volume, LV ejection fraction, and AMI mortality [105].
Similarly, patients with acute myocardial infarction demonstrated elevated levels of plasma miRNA-21 compared with their healthy counterparts. miRNA-21 emerged as a novel marker with predictive potential for LV remodeling post-AMI [106]. Additionally, miRNA-21 correlated with conventional markers of an AMI, including creatine kinase-MB (CK-MB), creatine kinase (CK), and cardiac troponin I (cTnI), exhibiting a comparable diagnostic accuracy [107].
In comparison with miRNA-1, miRNA-499 exhibited superior predictive value over the most consistent AMI biomarkers, such as cTnI and CK-MB. Alterations in the levels of circulating miRNA-499 were found to be related to unstable angina and non-ST elevation MI [108]. Moreover, miRNA-210 was recently found to differentiate between different clinical presentations of coronary artery disease [109]. Interestingly, it discriminated between individuals in the non-CAD group and those with unstable angina and myocardial infarction [109].
5.3. mitomiRs as Therapeutic Targets
Despite years of promising basic research, pre-clinical, and early clinical trial findings, to date, miRNAs have not been approved for the treatment of any ACVD [5]. Many miRNAs influence multiple signaling pathways and regulate numerous physiological processes, making them attractive but challenging candidates for therapy.
Only a few studies explored the potential of mitomiRs as therapeutic targets in ACVD. For instance, miRNA-144 demonstrated protective effects against adverse post-MI remodeling in both ischemia/reperfusion and non-reperfused MI mouse models [110]. Administering miRNA-144 through repeated intravenous injections reduces the infarct size and improves cardiac function in a mouse model of MI induced by LAD coronary artery ligation. miRNA-144 provides acute cardioprotection and mitigates chronic MI-induced remodeling. Furthermore, intracardiac injection of either miR-19a or miR-19b mimics a mouse model of MI with permanently ligated LAD coronary artery reduced infarct size, preserved cardiac function from 5 days to 9 weeks post-MI, and increased survival [111].
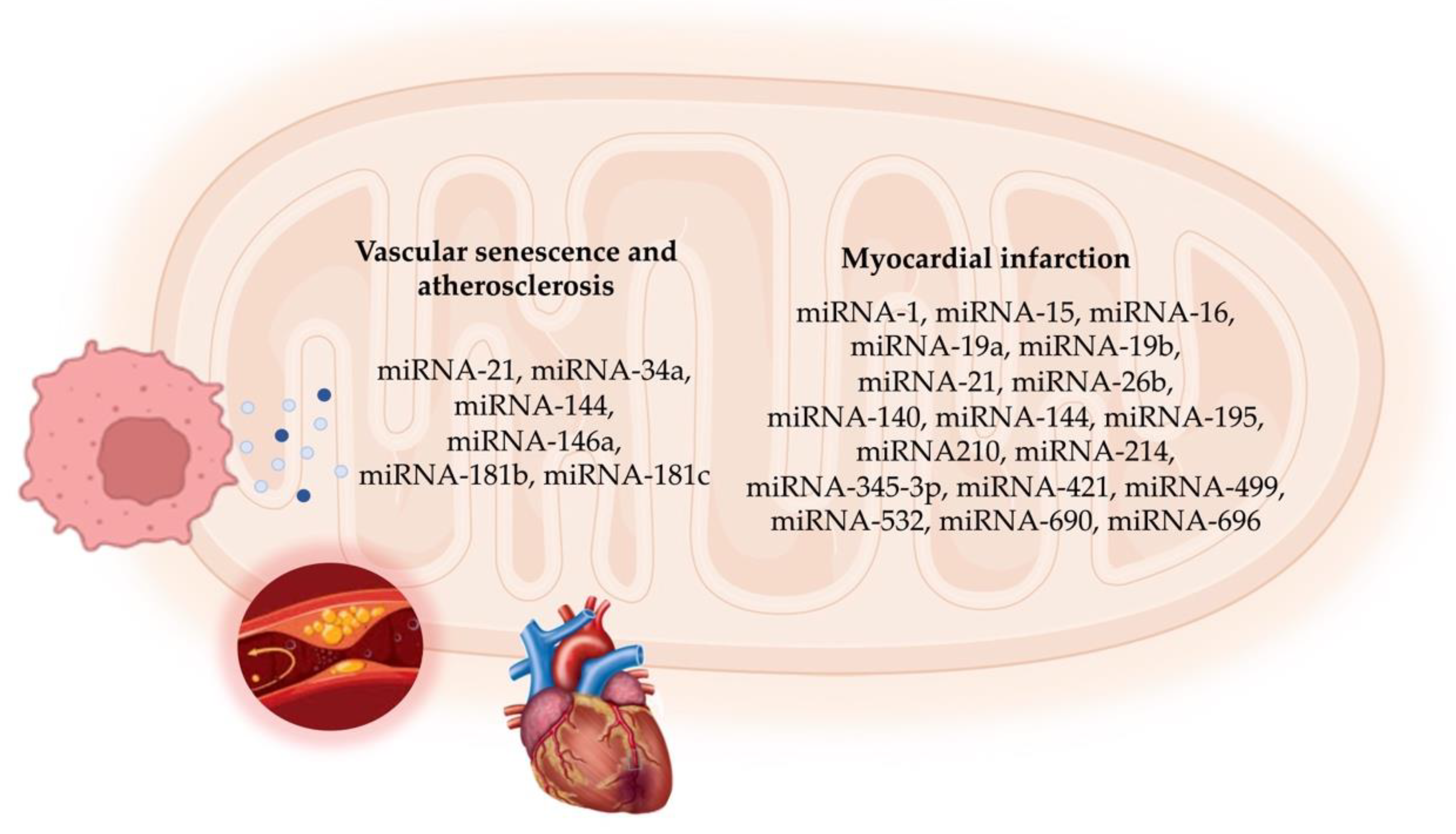
In MI, overcoming microvascular obstruction is crucial for delivering therapeutic agents to the infarct site. Hong et al. [112] discovered an anti-coagulative nanocomplex to deliver a miRNA-1 inhibitor loaded with dendrigraft poly-L-lysine. This complex effectively reduced thrombus formation in microvessels and inhibited coagulation factor Xa, thus overcoming microvascular obstruction in the infarct area [112]. Additionally, the miRNA-1 inhibitor reduced cardiomyocyte apoptosis and fibrosis, ultimately improving cardiac function. A parallel strategy by Bejerano et al. [113] involved nanoparticle-based delivery of a miRNA-21 mimic to cardiac macrophages at the infarct site in a mouse model. Treatment with the miRNA-21 mimic shifted cardiac macrophages from a pro-inflammatory to an anti-inflammatory state, leading to increased angiogenesis, decreased apoptosis, and reduced pathological remodeling in the infarct area [106]. Figure 2 summarizes the mitomiRs involved in cellular senescence and ACVD.
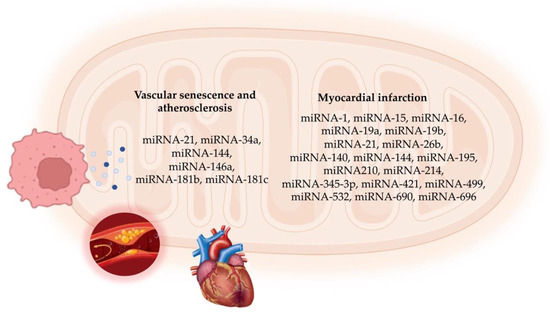
Figure 2.
Schematic representation of mitomiRs involved in vascular cell senescence and atherosclerotic cardiovascular disease.
6. Concluding Remarks and Future Perspectives
miRNAs are small molecules that play crucial roles in regulating cellular processes. They are easily transcribed, synthesized, transported, and degraded, making them ideal for regulating functions within cells, including shuttling between the nucleus and mitochondria. While much research has focused on miRNA functions in the nucleus and cytoplasm, there is a growing interest in understanding mitomiRs, which are a subset of miRNAs that specifically regulate mitochondrial homeostasis.
mitomiRs tightly regulate various aspects of mitochondrial function, including metabolism, redox status, apoptosis, dynamics, and DNA maintenance.
Emerging knowledge suggests that mitomiRs may play a major role in regulating several hallmarks of vascular senescence and ACVD related to mitochondria, such as mitochondrial function and metabolic regulation. These findings open a new field to explore novel molecular mechanisms controlling mitochondrial homeostasis in cardiovascular disease, highlighting these miRNAs as promising therapeutic targets. Novel strategies to reduce myocardial cell death, inhibit adverse remodeling, and/or stimulate heart regeneration are necessary.
However, research on mitomiRs is still in its early stages. Many questions remain unanswered, such as how they are transported to mitochondria and how they are transcribed from the mitochondrial genome. Additionally, while in vitro and animal studies show the potential of mitomiRs as diagnostic and prognostic markers, translating findings to humans requires further investigation. Retrospective and prospective non-interventional cohort studies involving patients with ACVD have the potential to significantly advance our understanding of how mitomiRs influence the clinical presentation of ischemic heart disease and atherosclerotic plaque phenotypes. Of importance, exploring the interplay between telomere dysfunction and mitochondrial dysmetabolism, mediated by mitomiRs, could provide insights into ACVD.
In conclusion, our review highlights the significance of mitochondrial-related miRNAs in vascular aging and ACVD, with the aim of developing novel diagnostic and therapeutic strategies in the future.
Author Contributions
Conceptualization, A.B.; writing—original draft preparation, P.C. and A.B.; writing—review and editing, P.C. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest. This research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- World Health Organization. Cardiovascular Diseases (CVDs). Available online: https://www.who.int/en/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 6 May 2024).
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgözoğlu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C.; Bennett, M. Aging and atherosclerosis: Mechanisms, functional consequences, and potential therapeutics for cellular senescence. Circ. Res. 2012, 111, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.C.Y.; Climie, R.E.; Shu, M.; Grieve, S.M.; Kozor, R.; Figtree, G.A. Vascular aging and cardiovascular disease: Pathophysiology and measurement in the coronary arteries. Front. Cardiovasc. Med. 2023, 10, 1206156. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.S.; Jin, J.P.; Wang, J.Q.; Zhang, Z.G.; Freedman, J.H.; Zheng, Y.; Cai, L. miRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta. Pharm. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Thavarajah, T.; Gu, W.; Cai, J.; Xu, Q. Impact of miRNA in Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Churov, A.; Summerhill, V.; Grechko, A.; Orekhova, V.; Orekhov, A. MicroRNAs as Potential Biomarkers in Atherosclerosis. Int. J. Mol. Sci. 2019, 20, 5547. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.N. MicroRNAs as therapeutic targets and biomarkers of cardiovascular disease. Sci. Transl. Med. 2014, 6, 239ps3. [Google Scholar] [CrossRef]
- Bian, Z.; Li, L.M.; Tang, R.; Hou, D.X.; Chen, X.; Zhang, C.Y.; Zen, K. Identification of mouse liver mitochondria-associated miRNAs and their potential biological functions. Cell Res. 2010, 20, 1076–1078. [Google Scholar] [CrossRef]
- Barrey, E.; Saint-Auret, G.; Bonnamy, B.; Damas, D.; Boyer, O.; Gidrol, X. Pre-microRNA and mature microRNA in human mitochondria. PLoS ONE 2011, 6, 20220. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Neph, S.; Dinger, M.E.; Crawford, J.; Smith, M.A.; Shearwood, A.M.; Haugen, E.; Bracken, C.P.; Rackham, O.; Stamatoyannopoulos, J.A.; et al. The human mitochondrial transcriptome. Cell 2011, 146, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ferlito, M.; Kent, O.A.; Fox-Talbot, K.; Wang, R.; Liu, D.; Raghavachari, N.; Yang, Y.; Wheelan, S.J.; Murphy, E.; et al. Nuclear miRNA regulates the mitochondrial genome in the heart. Circ. Res. 2012, 110, 1596–1603. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Tian, T.; Chen, W.; Lv, X.; Lei, X.; Zhang, H.; Sun, S.; Cai, L.; Pan, G.; He, L.; et al. Mitochondrial miRNA Determines Chemoresistance by Reprogramming Metabolism and Regulating Mitochondrial Transcription. Cancer Res. 2019, 79, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Matégot, R.; Girard, M.; Demongeot, J.; Henrion-Caude, A. MitomiRs delineating the intracellular localization of microRNAs at mitochondria. Free Radic. Biol. Med. 2013, 64, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Macgregor-Das, A.M.; Das, S. A microRNA’s journey to the center of the mitochondria. Am. J. Physiol. Circ. Physiol. 2018, 315, 206–215. [Google Scholar] [CrossRef]
- Roiz-Valle, D.; Caravia, X.M.; López-Otín, C. Mechanisms of mitochondrial microRNA regulation in cardiovascular diseases. Mech. Ageing Dev. 2023, 212, 111822. [Google Scholar] [CrossRef]
- Song, R.; Hu, X.Q.; Zhang, L. Mitochondrial MiRNA in Cardiovascular Function and Disease. Cells 2019, 8, 1475. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, H.; Das, S. Mitochondrial miRNA (MitomiR): A new player in cardiovascular health. Can. J. Physiol. Pharmacol. 2015, 93, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2015, 15, 509–524. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.H.; Nam, J.W.; Heo, I.; Rhee, J.K.; Sohn, S.Y.; Cho, Y.; Zhang, B.T.; Kim, V.N. Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Hwang, H.W.; Wentzel, E.A.; Mendell, J.T. A hexanucleotide element directs microRNA nuclear import. Science 2007, 315, 97–100. [Google Scholar] [CrossRef]
- Meister, G.; Landthaler, M.; Patkaniowska, A.; Dorsett, Y.; Teng, G.; Tuschl, T. Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell. 2004, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Jiao, J.; Gao, G.; Prabhakar, B.S. Control of mitochondrial activity by miRNAs. J. Cell Biochem. 2012, 113, 1104–1110. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [PubMed]
- Latronico, M.V.; Condorelli, G. The might of microRNA in mitochondria. Circ. Res. 2012, 110, 1540–1542. [Google Scholar] [CrossRef] [PubMed]
- Meloni, M.; Marchetti, M.; Garner, K.; Littlejohns, B.; Sala-Newby, G.; Xenophontos, N.; Floris, I.; Suleiman, M.S.; Madeddu, P.; Caporali, A.; et al. Local inhibition of microRNA-24 improves reparative angiogenesis and left ventricle remodeling and function in mice with myocardial infarction. Mol. Ther. 2013, 21, 1390–1402. [Google Scholar] [CrossRef]
- Wang, W.X.; Visavadiya, N.P.; Pandya, J.D.; Nelson, P.T.; Sullivan, P.G.; Springer, J.E. Mitochondria-associated microRNAs in rat hippocampus following traumatic brain injury. Exp. Neurol. 2015, 265, 84–93. [Google Scholar] [CrossRef]
- Zhang, X.; Zuo, X.; Yang, B.; Li, Z.; Xue, Y.; Zhou, Y.; Huang, J.; Zhao, X.; Zhou, J.; Yan, Y.; et al. MicroRNA directly enhances mitochondrial translation during muscle differentiation. Cell 2014, 158, 607–619. [Google Scholar] [CrossRef]
- Gohel, D.; Sripada, L.; Prajapati, P.; Currim, F.; Roy, M.; Singh, K.; Shinde, A.; Mane, M.; Kotadia, D.; Tassone, F.; et al. Expression of expanded FMR1-CGG repeats alters mitochondrial miRNAs and modulates mitochondrial functions and cell death in cellular model of FXTAS. Free Radic. Biol. Med. 2021, 165, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, H.W.; Oktay, Y.; Zhang, J.; Allen, E.L.; Smith, G.M.; Fan, K.C.; Hong, J.S.; French, S.W.; McCaffery, J.M.; et al. PNPASE regulates RNA import into mitochondria. Cell 2010, 142, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, D.L.; Hathaway, Q.A.; Pinti, M.V.; Nichols, C.E.; Durr, A.J.; Sreekumar, S.; Hughes, K.M.; Stine, S.M.; Martinez, I.; Hollander, J.M. Exploring the mitochondrial microRNA import pathway through Polynucleotide Phosphorylase (PNPase). J. Mol. Cell Cardiol. 2017, 110, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Gibbings, D.J.; Ciaudo, C.; Erhardt, M.; Voinnet, O. Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity. Nat. Cell Biol. 2009, 11, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Jusic, A.; Devaux, Y. Mitochondrial noncoding RNA-regulatory network in cardiovascular disease. Basic. Res. Cardiol. 2020, 115, 23. [Google Scholar] [CrossRef] [PubMed]
- Shinde, S.; Bhadra, U. A complex genome-microRNA interplay in human mitochondria. Biomed. Res. Int. 2015, 2015, 206382. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Shearwood, A.M.; Mercer, T.R.; Davies, S.M.; Mattick, J.S.; Filipovska, A. Long noncoding RNAs are generated from the mitochondrial genome and regulated by nuclear-encoded proteins. RNA 2011, 17, 2085–2093. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Rüberg, S.; Girard, M.; Cagnard, N.; Hanein, S.; Chrétien, D.; Munnich, A.; Lyonnet, S.; Henrion-Caude, A. Nuclear outsourcing of RNA interference components to human mitochondria. PLoS ONE 2011, 6, 20746. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Ji, H.; Cho, E.; Park, N.H.; Hwang, K.; Park, W.; Lee, K.S.; Park, D.; Jung, E. nc886, a Non-Coding RNA, Is a New Biomarker and Epigenetic Mediator of Cellular Senescence in Fibroblasts. Int. J. Mol. Sci. 2021, 22, 13673. [Google Scholar] [CrossRef]
- Sripada, L.; Tomar, D.; Prajapati, P.; Singh, R.; Singh, A.K.; Singh, R. Systematic analysis of small RNAs associated with human mitochondria by deep sequencing: Detailed analysis of mitochondrial associated miRNA. PLoS ONE 2012, 9, 44873. [Google Scholar] [CrossRef]
- Murri, M.; El Azzouzi, H. MicroRNAs as regulators of mitochondrial dysfunction and obesity. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Kussainova, A.; Bulgakova, O.; Aripova, A.; Khalid, Z.; Bersimbaev, R.; Izzotti, A. The role of mitochondrial miRNAs in the development of radon-induced lung cancer. Biomedicines 2022, 2, 428. [Google Scholar] [CrossRef] [PubMed]
- Kuthethur, R.; Shukla, V.; Mallya, S.; Adiga, D.; Kabekkodu, S.P.; Ramachandra, L.; Saxena, P.U.P.; Satyamoorthy, K.; Chakrabarty, S. Expression analysis and function of mitochondrial genome-encoded microRNAs. J. Cell Sci. 2022, 135, 258937. [Google Scholar] [CrossRef] [PubMed]
- Santovito, D.; Weber, C. Non-canonical features of microRNAs: Paradigms emerging from cardiovascular disease. Nat. Rev. Cardiol. 2022, 19, 620–638. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Smith, F.; Kumar, S.; Vijayan, M.; Reddy, P.H. Are microRNAs true sensors of ageing and cellular senescence? Ageing Res. Rev. 2017, 35, 350–363. [Google Scholar] [CrossRef]
- Vecoli, C.; Borghini, A.; Andreassi, M.G. The molecular biomarkers of vascular aging and atherosclerosis: Telomere length and mitochondrial DNA4977 common deletion. Mutat. Res. Rev. Mutat. Res. 2020, 784, 108309. [Google Scholar] [CrossRef]
- Borghini, A.; Cervelli, T.; Galli, A.; Andreassi, M.G. DNA modifications in atherosclerosis: From the past to the future. Atherosclerosis 2013, 230, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Miyauchi, H.; Yoshida, T.; Ishida, Y.; Yoshida, H.; Komuro, I. Endothelial cell senescence in human atherosclerosis: Role of telomere in endothelial dysfunction. Circulation 2002, 105, 1541–1544. [Google Scholar] [CrossRef]
- Matthews, C.; Gorenne, I.; Scott, S.; Figg, N.; Kirkpatrick, P.; Ritchie, A.; Goddard, M.; Bennett, M. Vascular smooth muscle cells undergo telomere-based senescence in human atherosclerosis: Effects of telomerase and oxidative stress. Circ. Res. 2006, 99, 156–164. [Google Scholar] [CrossRef]
- Yu, E.; Calvert, P.A.; Mercer, J.R.; Harrison, J.; Baker, L.; Figg, N.L.; Kumar, S.; Wang, J.C.; Hurst, L.A.; Obaid, D.R.; et al. Mitochondrial DNA damage can promote atherosclerosis independently of reactive oxygen species through effects on smooth muscle cells and monocytes and correlates with higher-risk plaques in humans. Circulation 2013, 128, 702–712. [Google Scholar] [CrossRef]
- Ataei Ataabadi, E.; Golshiri, K.; Jüttner, A.A.; de Vries, R.; Van den Berg-Garrelds, I.; Nagtzaam, N.M.A. Vascular Ageing Features Caused by Selective DNA Damage in Smooth Muscle Cell. Oxid. Med. Cell Longev. 2021, 2021, 2308317. [Google Scholar] [CrossRef] [PubMed]
- Vecoli, C.; Borghini, A.; Pulignani, S.; Mercuri, A.; Turchi, S.; Carpeggiani, C.; Picano, E.; Andreassi, M.G. Prognostic value of mitochondrial DNA4977 deletion and mitochondrial DNA copy number in patients with stable coronary artery disease. Atherosclerosis 2018, 276, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Vecoli, C.; Borghini, A.; Pulignani, S.; Mercuri, A.; Turchi, S.; Picano, E.; Andreassi, M.G. Independent and Combined Effects of Telomere Shortening and mtDNA4977 Deletion on Long-term Outcomes of Patients with Coronary Artery Disease. Int. J. Mol. Sci. 2019, 20, 5508. [Google Scholar] [CrossRef] [PubMed]
- Andreassi, M.G.; Borghini, A.; Vecoli, C. Micronucleus assay for predicting coronary artery disease: A systematic review and meta-analysis. Mutat. Res. Rev. Mutat. Res. 2021, 787, 108348. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Sorond, F.; Merkely, B.; Csiszar, A. Mechanisms of Vascular Aging, A Geroscience Perspective: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; DePinho, R.A. Axis of ageing: Telomeres, p53 and mitochondria. Nat. Rev. Mol. Cell Biol. 2012, 13, 397–404. [Google Scholar] [CrossRef]
- Sahi, E.; Colla, S.; Liesa, M.; Moslehi, J.; Müller, F.L.; Guo, M.; Cooper, M.; Kotton, D.; Fabian, A.J.; Walkey, C.; et al. Telomere dysfunction induces metabolic and mitochondrial compromise. Nature 2011, 470, 359–365. [Google Scholar]
- Fang, E.F.; Scheibye-Knudsen, M.; Chua, K.F.; Mattson, M.P.; Croteau, D.L.; Bohr, V.A. Nuclear DNA damage signalling to mitochondria in aging. Nat. Rev. Mol. Cell Biol. 2016, 17, 308–321. [Google Scholar] [CrossRef]
- Weber, T.A.; Reichert, A.S. Impaired quality control of mitochondria: Aging from a new perspective. Exp. Gerontol. 2010, 45, 503–511. [Google Scholar] [CrossRef]
- Pan, S.; Ryu, S.Y.; Sheu, S.S. Distinctive characteristics and functions of multiple mitochondrial Ca2+ influx mechanisms. Sci. China Life Sci. 2011, 54, 763–769. [Google Scholar] [CrossRef]
- Marí, M.; de Gregorio, E.; de Dios, C.; Roca-Agujetas, V.; Cucarull, B.; Tutusaus, A.; Morales, A.; Colell, A. Mitochondrial Glutathione: Recent Insights and Role in Disease. Antioxidants 2020, 9, 909. [Google Scholar] [CrossRef] [PubMed]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in skin health, aging, and disease. Cell Death Dis. 2020, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.J.; Goldstein, D.R. Ageing and atherosclerosis: Vascular intrinsic and extrinsic factors and potential role of IL-6. Nat. Rev. Cardiol. 2021, 18, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; Perry, J.B.; Allen, M.E.; Sabbah, H.N.; Stauffer, B.L.; Shaikh, S.R.; Cleland, J.G.; Colucci, W.S.; Butler, J.; Voors, A.A.; et al. Expert consensus document: Mitochondrial function as a therapeutic target in heart failure. Nat. Rev. Cardiol. 2017, 14, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Shkurat, T.P.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. The role of mitochondrial dysfunction in cardiovascular disease: A brief review. Ann. Med. 2018, 50, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Jin, X.; Li, D.; Lu, J.; Zhang, X.N.; Yang, S.J.; Zhao, Y.X.; Wu, M. New insights into vascular aging: Emerging role of mitochondria function. Biomed. Pharmacother. 2022, 156, 113954. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; An, X.; Xiao, Y.; Sun, X.; Li, S.; Wang, Y.; Sun, W.; Yu, D. Mitochondrial-related microRNAs and their roles in cellular senescence. Front. Physiol. 2024, 14, 1279548. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Kohr, M.; Dunkerly-Eyring, B.; Lee, D.I.; Bedja, D.; Kent, O.A.; Leung, A.K.; Henao-Mejia, J.; Flavell, R.A.; Steenbergen, C. Divergent Effects of miR-181 Family Members on Myocardial Function Through Protective Cytosolic and Detrimental Mitochondrial microRNA Targets. J. Am. Heart Assoc. 2017, 6, 004694. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, D.; Cheng, H.S.; McCoy, M.G.; Pérez-Cremades, D.; Haemmig, S.; Wong, D.; Chen, L.; Feinberg, M.W. miR-181b regulates vascular endothelial aging by modulating an MAP3K3 signaling pathway. FASEB J. 2022, 36, 22353. [Google Scholar] [CrossRef]
- Giuliani, A.; Cirilli, I.; Prattichizzo, F.; Mensà, E.; Fulgenzi, G.; Sabbatinelli, J.; Graciotti, L.; Olivieri, F.; Procopio, A.D.; Tiano, L.; et al. The mitomiR/Bcl-2 axis affects mitochondrial function and autophagic vacuole formation in senescent endothelial cells. Aging 2018, 10, 2855–2873. [Google Scholar] [CrossRef]
- Giuliani, A.; Prattichizzo, F.; Micolucci, L.; Ceriello, A.; Procopio, A.D.; Rippo, M.R. Mitochondrial (dys) function in inflammaging: Do MitomiRs influence the energetic, oxidative, and inflammatory status of senescent cells? Mediat. Inflamm. 2017, 2017, 2309034. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Lazzarini, R.; Recchioni, R.; Marcheselli, F.; Rippo, M.R.; Di Nuzzo, S.; Albertini, M.C.; Graciotti, L.; Babini, L.; Mariotti, S.; et al. MiR-146a as marker of senescence-associated pro-inflammatory status in cells involved in vascular remodelling. Age 2013, 35, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J. miR-34a repression of SIRT1 regulates apoptosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13421–13426. [Google Scholar] [CrossRef] [PubMed]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, metabolism and cardiovascular disease. J. Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Prattichizzo, F.; Giuliani, A.; Matacchione, G.; Rippo, M.R.; Sabbatinelli, J.; Bonafè, M. miR-21 and miR-146a: The microRNAs of inflammaging and age-related diseases. Ageing Res. Rev. 2021, 70, 101374. [Google Scholar] [CrossRef] [PubMed]
- Dellago, H.; Preschitz-Kammerhofer, B.; Terlecki-Zaniewicz, L.; Schreiner, C.; Fortschegger, K.; Chang, M.W.; Hackl, M.; Monteforte, R.; Kühnel, H.; Schosserer, M.; et al. High levels of oncomiR-21 contribute to the senescence-induced growth arrest in normal human cells and its knock-down increases the replicative lifespan. Aging Cell 2013, 12, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Mensà, E.; Guescini, M.; Giuliani, A.; Bacalini, M.G.; Ramini, D.; Corleone, G.; Ferracin, M.; Fulgenzi, G.; Graciotti, L.; Prattichizzo, F.; et al. Small extracellular vesicles deliver miR-21 and miR-217 as pro-senescence effectors to endothelial cells. J. Extracell. Vesicles 2020, 9, 1725285. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Yu, Y.; Dong, H.; Bian, X.; Guo, X.; Dong, S. MicroRNA 21 inhibits left ventricular remodeling in the early phase of rat model with ischemia-reperfusion injury by suppressing cell apoptosis. Int. J. Med. Sci. 2012, 9, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Nasci, V.L.; Chuppa, S.; Griswold, L.; Goodreau, K.A.; Dash, R.K.; Kriegel, A.J. Mir-21-5p regulates mitochondrial respiration and lipid content in H9C2 cells. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, 710–721. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Wang, F.; Zhou, L.; Yin, Z.; Fan, J.; Nie, X.; Wang, P.; Fu, X.D.; Chen, C.; et al. MicroRNA-21 Lowers Blood Pressure in Spontaneous Hypertensive Rats by Upregulating Mitochondrial Translation. Circulation 2016, 134, 734–751. [Google Scholar] [CrossRef]
- Fu, X.; Huang, X.; Li, P.; Chen, W.; Xia, M. 7-Ketocholesterol inhibits isocitrate dehydrogenase 2 expression and impairs endothelial function via microRNA-144. Free Radic. Biol. Med. 2014, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Li, P.; Li, J.; He, R.; Cheng, G.; Li, Y. Downregulation of microRNA-34a inhibits oxidized low-density lipoprotein-induced apoptosis and oxidative stress in human umbilical vein endothelial cells. Int. J. Mol. Med. 2018, 42, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Mendell, J.T.; Olson, E.N. MicroRNAs in stress signaling and human disease. Cell 2012, 148, 1172–1187. [Google Scholar] [CrossRef] [PubMed]
- Engedal, N.; Zerovnik, E.; Rudov, A.; Galli, F.; Olivieri, F.; Procopio, A.D.; Rippo, M.R.; Monsurro, V.; Betti, M.; Albertini, M.C. From Oxidative Stress Damage to Pathways, Networks, and Autophagy via MicroRNAs. Oxid. Med. Cell Longev. 2018, 2018, 4968321. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, J.; Thum, T. MicroRNAs in myocardial infarction. Arter. Thromb. Vasc. Biol. 2013, 33, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, C.; Zhou, X.; Han, X.; Li, J.; Wang, Z.; Shang, H.; Liu, Y.; Cao, H. Mitochondria Associated MicroRNA Expression Profiling of Heart Failure. Biomed. Res. Int. 2017, 2017, 4042509. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Mizushima, K.; Takanami, Y.; Kawai, Y.; Ichikawa, H.; Yoshikawa, T. The microRNA miR-696 regulates PGC-1{alpha} in mouse skeletal muscle in response to physical activity. Am. J. Physiol. Metab. 2010, 298, 799–806. [Google Scholar]
- Wang, J.X.; Zhang, X.J.; Feng, C.; Sun, T.; Wang, K.; Wang, Y.; Zhou, L.Y.; Li, P.F. MicroRNA-532-3p regulates mitochondrial fission through targeting apoptosis repressor with caspase recruitment domain in doxorubicin cardiotoxicity. Cell Death Dis. 2015, 6, 1677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Guan, P.; Ye, X.; Lu, Y.; Hang, Y.; Su, Y.; Hu, W. SOCS6 promotes mitochondrial fission and cardiomyocyte apoptosis and is negatively regulated by quaking-mediated miR-19b. Oxid. Med. Cell. Longev. 2022, 2022, 1121323. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lee, D.S.; Choong, O.K.; Chang, S.K.; Hsu, T.; Nicholson, M.W.; Liu, L.W.; Lin, P.J.; Ruan, S.C.; Lin, S.W.; et al. Cardiac-specific microRNA-125b deficiency induces perinatal death and cardiac hypertrophy. Sci. Rep. 2021, 11, 2377. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Jiao, J.; Wang, J.; Li, Y.; Qin, D.; Li, P. Mitofusin 1 is negatively regulated by microRNA 140 in cardiomyocyte apoptosis. Mol. Cell Biol. 2014, 34, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, L.Y.; Wang, J.X.; Wang, Y.; Sun, T.; Zhao, B.; Yang, Y.J.; An, T.; Long, B.; Li, N.; et al. E2F1-dependent miR-421 regulates mitochondrial fragmentation and myocardial infarction by targeting Pink1. Nat. Commun. 2015, 6, 7619. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Zhang, H.; Huang, P.; Luthra, R. Hypoxia-regulated microRNA-210 modulates mitochondrial function and decreases ISCU and COX10 expression. Oncogene 2010, 29, 4362–4368. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; An, T.; Zhai, M.; Huang, Y.; Wang, Q.; Wang, Y.; Zhang, R.; Wang, T.; Liu, J.; Zhang, Y.; et al. Mitochondrial miR-762 regulates apoptosis and myocardial infarction by impairing ND2. Cell Death Dis. 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Zhang, Y.Y.; Hemann, C.; Mahoney, C.E.; Zweier, J.L.; Loscalzo, J. MicroRNA-210 controls mitochondrial metabolism during hypoxia by repressing the iron-sulfur cluster assembly proteins ISCU1/2. Cell Metab. 2009, 10, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Huang, M.; Li, Z.; Jia, F.; Ghosh, Z.; Lijkwan, M.A.; Fasanaro, P.; Sun, N.; Wang, X.; Martelli, F.; et al. MicroRNA-210 as a novel therapy for treatment of ischemic heart disease. Circulation 2010, 122, 124–131. [Google Scholar] [CrossRef]
- Zhang, X.; Ji, R.; Liao, X.; Castillero, E.; Kennel, P.J.; Brunjes, D.L.; Franz, M.; Möbius-Winkler, S.; Drosatos, K.; George, I.; et al. MicroRNA-195 regulates metabolism in failing myocardium via alterations in Sirtuin 3 expression and mitochondrial protein acetylation. Circulation 2018, 137, 2052–2067. [Google Scholar] [CrossRef] [PubMed]
- Hullinger, T.G.; Montgomery, R.L.; Seto, A.G.; Dickinson, B.A.; Semus, H.M.; Lynch, J.M.; Dalby, C.M.; Robinson, K.; Stack, C.; Latimer, P.A.; et al. Inhibition of miR-15 protects against cardiac ischemic injury. Circ. Res. 2012, 110, 71–81. [Google Scholar] [CrossRef]
- Wang, J.X.; Jiao, J.Q.; Li, Q.; Long, B.; Wang, K.; Liu, J.P.; Li, Y.R.; Li, P.F. miR-499 regulates mitochondrial dynamics by targeting calcineurin and dynamin-related protein-1. Nat. Med. 2011, 17, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Lv, G.; Shao, S.; Dong, H.; Bian, X.; Yang, X.; Dong, S. MicroRNA-214 protects cardiac myocytes against H2O2-induced injury. J. Cell Biochem. 2014, 115, 93–101. [Google Scholar] [CrossRef]
- Long, B.; Wang, K.; Li, N.; Murtaza, I.; Xiao, J.Y.; Fan, Y.Y.; Liu, C.Y.; Li, W.H.; Cheng, Z.; Li, P. miR-761 regulates the mitochondrial network by targeting mitochondrial fission factor. Free Radic. Biol. Med. 2013, 65, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Long, B.; Jiao, J.Q.; Wang, J.X.; Liu, J.P.; Li, Q.; Li, P.F. miR-484 regulates mitochondrial network through targeting Fis1. Nat. Commun. 2012, 3, 781. [Google Scholar] [CrossRef] [PubMed]
- Townley-Tilson, W.H.; Callis, T.E.; Wang, D. MicroRNAs 1, 133, and 206: Critical factors of skeletal and cardiac muscle development, function, and disease. Int. J. Biochem. Cell Biol. 2010, 42, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tan, N.; Yang, J.; Liu, X.; Cao, X.; He, P.; Dong, X.; Qin, S.; Zhang, C. A translational study of circulating cell-free microRNA-1 in acute myocardial infarction. Clin. Sci. 2010, 119, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.J.; Liu, T.; Zhang, H.; Yang, S.J. Plasma microRNA-21 is a potential diagnostic biomarker of acute myocardial infarction. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 323–329. [Google Scholar] [PubMed]
- Liu, X.; Dong, Y.; Chen, S.; Zhang, G.; Zhang, M.; Gong, Y.; Li, X. Circulating microRNA-146a and microRNA-21 predict left ventricular remodeling after ST-elevation myocardial infarction. Cardiology 2015, 132, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, S.M.; El-Shal, A.S.; Shoukry, A.; Khedr, M.H.; Abdelraheim, N. Serum miRNA-499 and miRNA-210: A potential role in early diagnosis of acute coronary syndrome. IUBMB Life 2016, 68, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Ozuynuk-Ertugrul, A.S.; Ekici, B.; Erkan, A.F.; Coban, N. Alteration of circulating miRNAs during myocardial infarction and association with lipid levels. Lab. Med. 2023, 55, 361–372. [Google Scholar] [CrossRef]
- Li, J.; Cai, S.X.; He, Q.; Zhang, H.; Friedberg, D.; Wang, F.; Redington, A.N. Intravenous miR-144 reduces left ventricular remodeling after myocardial infarction. Basic. Res. Cardiol. 2018, 113, 36. [Google Scholar] [CrossRef]
- Gao, F.; Kataoka, M.; Liu, N.; Liang, T.; Huang, Z.P.; Gu, F.; Ding, J.; Liu, J.; Zhang, F.; Ma, Q.; et al. Therapeutic role of miR-19a/19b in cardiac regeneration and protection from myocardial infarction. Nat. Commun. 2019, 10, 1802. [Google Scholar] [CrossRef]
- Hong, T.; Wei, Y.; Xue, X.; Li, Y.; Dong, H.; Guo, X.; Shi, X.; He, B. A novel anti-coagulative nanocomplex in delivering miRNA-1 inhibitor against microvascular obstruction of myocardial infarction. Adv. Healthc. Mater. 2020, 9, 1901783. [Google Scholar] [CrossRef] [PubMed]
- Bejerano, T.; Etzion, S.; Elyagon, S.; Etzion, Y.; Cohen, S. Nanoparticle delivery of miRNA-21 mimic to cardiac macrophages improves myocardial remodeling after myocardial infarction. Nano Lett. 2018, 18, 5885–5891. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).