
Urtica dioica Extract Abrogates Chlorpyrifos-Induced Toxicity in Zebrafish Larvae

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. DI-HRMS Analysis
2.2. Hatching and Survival Rates
2.3. Teratology Screening
2.4. The Effects of UDE on the Brain-Derived Neurotrophic Factor Expression in ‘Larvae’ Telencephalon
2.5. Locomotor Activity
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Extraction
4.2. Direct-Infusion High-Resolution Mass Spectrometry Analysis
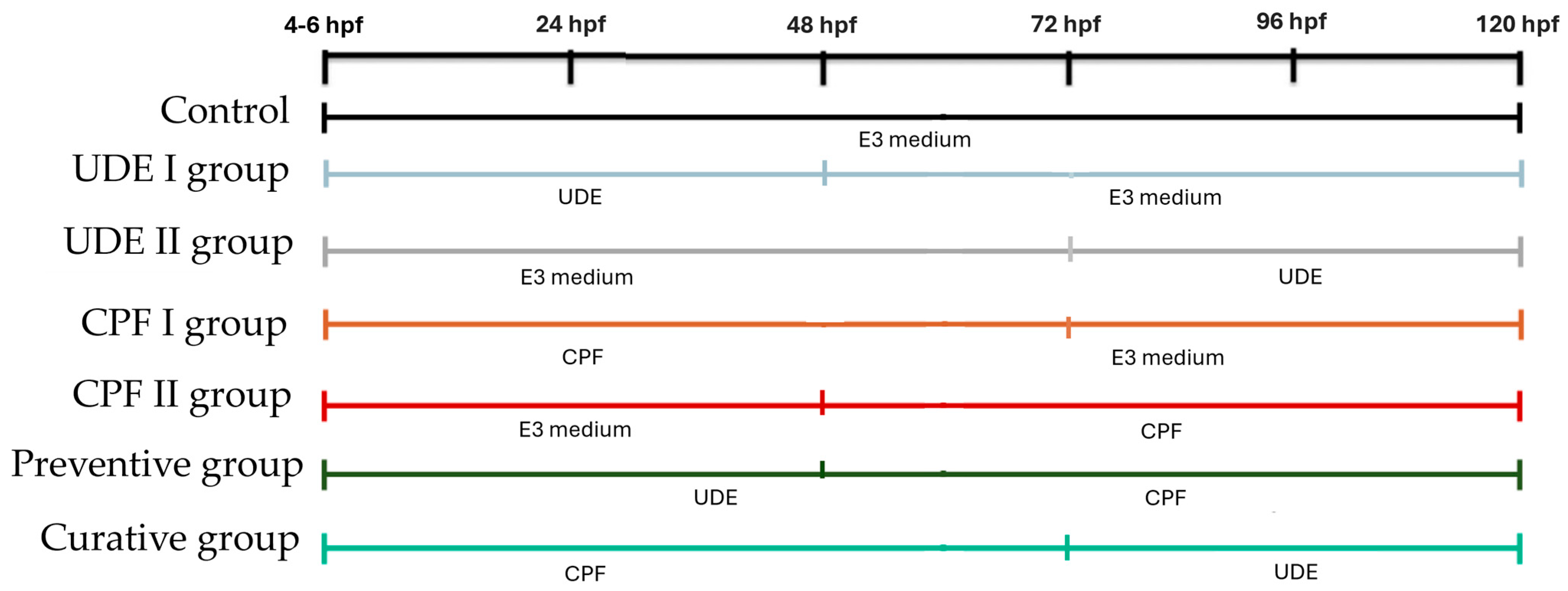
4.3. Embryos Maintenance and Treatments
- (1)
- The control group with no treatment.
- (2)
- The UDE I group, which was treated with UDE freshly prepared in E3 medium at a final concentration of 25 mg/L, starting at 4–6 hpf until 48 hpf was reached.
- (3)
- The UDE II group, which was treated with UDE freshly prepared in E3 medium at a final concentration of 25 mg/L, starting at 72 hpf until the end of the experiment was reached.
- (4)
- The CPF I group, which was treated with chlorpyrifos (campagnie, Sfax, Tunisia) from 4–6 hpf to 72 hpf, at a final concentration of 1 mg/L dissolved in E3 medium. This concentration was chosen based on LC50 = 1.520 mg/L, with a 95% confidence limit of 1.26–1.82 mg/L [29], which is moderately toxic to zebrafish larvae.
- (5)
- The CPF II group, which was treated with chlorpyrifos (campagnie, Sfax, Tunisia) from 48 hpf to 120 hpf at a final concentration of 1 mg/L dissolved in E3 medium.
- (6)
- The UDE+CPF preventive group in which UDE was maintained from 4–6 hpf to 48 hpf and CPF was administered from 48 hpf to 120 hpf.
- (7)
- The CPF+UDE curative group in which CPF was maintained from 4–6 hpf to 72 hpf and UDE was administered at 72 hpf and maintained until 120 hpf.
4.4. Teratology Assessment
4.5. Tissue Processing and Immunofluorescence
4.6. BDNF Fluorescence Intensity at Telencephalon
4.7. Larval Behavior
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oltramare, C.; Weiss, F.T.; Staudacher, P.; Kibirango, O.; Atuhaire, A.; Stamm, C. Pesticides Monitoring in Surface Water of a Subsistence Agricultural Catchment in Uganda Using Passive Samplers. Environ. Sci. Pollut. Res. 2023, 30, 10312–10328. [Google Scholar] [CrossRef]
- EPA Chlorpyrifos. Available online: https://www.epa.gov/ingredients-used-pesticide-products/chlorpyrifos (accessed on 18 July 2023).
- Kuca, K.; Juna, D.; Musilek, K. Structural Requirements of Acetylcholinesterase Reactivators. Mini Rev. Med. Chem. 2006, 6, 269–277. [Google Scholar] [CrossRef]
- Eaton, D.L.; Daroff, R.B.; Autrup, H.; Bridges, J.; Buffler, P.; Costa, L.G.; Coyle, J.; McKhann, G.; Mobley, W.C.; Nadel, L.; et al. Review of the Toxicology of Chlorpyrifos with an Emphasis on Human Exposure and Neurodevelopment. Crit. Rev. Toxicol. 2008, 38, 1–125. [Google Scholar] [CrossRef]
- Mhalhel, K.; Sicari, M.; Pansera, L.; Chen, J.; Levanti, M.; Diotel, N.; Rastegar, S.; Germanà, A.; Montalbano, G. Zebrafish: A Model Deciphering the Impact of Flavonoids on Neurodegenerative Disorders. Cells 2023, 12, 252. [Google Scholar] [CrossRef]
- Chira, A.; Rekik, I.; Rahmouni, F.; Ben Amor, I.; Gargouri, B.; Kallel, C.; Jamoussi, K.; Allouche, N.; El Feki, A.; Kadmi, Y.; et al. Phytochemical Composition of Urtica Dioica Essential Oil with Antioxidant and Anti-Inflammatory Properties: In Vitro and in Vivo Studies. Curr. Pharm. Biotechnol. 2022, 23, 1–11. [Google Scholar] [CrossRef]
- Semwal, P.; Rauf, A.; Olatunde, A.; Singh, P.; Zaky, M.Y.; Islam, M.M.; Khalil, A.A.; Aljohani, A.S.M.; Al Abdulmonem, W.; Ribaudo, G. The Medicinal Chemistry of Urtica dioica, L.: From Preliminary Evidence to Clinical Studies Supporting Its Neuroprotective Activity. Nat. Prod. Bioprospect. 2023, 13, 16. [Google Scholar] [CrossRef]
- Patel, S.S.; Udayabanu, M. Urtica Dioica Extract Attenuates Depressive like Behavior and Associative Memory Dysfunction in Dexamethasone Induced Diabetic Mice. Metab. Brain Dis. 2014, 29, 121–130. [Google Scholar] [CrossRef]
- Rahmati, M.; Keshvari, M.; Mirnasouri, R.; Chehelcheraghi, F. Exercise and Urtica Dioica Extract Ameliorate Hippocampal Insulin Signaling, Oxidative Stress, Neuroinflammation, and Cognitive Function in STZ-Induced Diabetic Rats. Biomed Pharmacother. 2021, 139, 111577. [Google Scholar] [CrossRef]
- Zotti, M.; Colaianna, M.; Morgese, M.G.; Tucci, P.; Schiavone, S.; Avato, P.; Trabace, L. Carvacrol: From Ancient Flavoring to Neuromodulatory Agent. Molecules 2013, 18, 6161–6172. [Google Scholar] [CrossRef]
- Guan, X.; Li, X.; Yang, X.; Yan, J.; Shi, P.; Ba, L.; Cao, Y.; Wang, P. The Neuroprotective Effects of Carvacrol on Ischemia/Reperfusion-Induced Hippocampal Neuronal Impairment by Ferroptosis Mitigation. Life Sci. 2019, 235, 116795. [Google Scholar] [CrossRef]
- Gül, S.; Demirci, B.; Başer, K.H.C.; Akpulat, H.A.; Aksu, P. Chemical Composition and In Vitro Cytotoxic, Genotoxic Effects of Essential Oil from Urtica dioica L. Bull. Environ. Contam. Toxicol. 2012, 88, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Murakami, S.; Ashikawa, Y.; Sasagawa, S.; Umemoto, N.; Shimada, Y.; Tanaka, T. Zebrafish as a Systems Toxicology Model for Developmental Neurotoxicity Testing. Congenit. Anom. 2015, 55, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mhalhel, K.; Briglia, M.; Aragona, M.; Porcino, C.; Abbate, F.; Guerrera, M.C.; Laurà, R.; Krichen, Y.; Guerbej, H.; Germanà, A.; et al. Nothobranchius as a Model for Anorexia of Aging Research: An Evolutionary, Anatomical, Histological, Immunohistochemical, and Molecular Study. Ann. Anat.-Anat. Anz. 2023, 250, 152116. [Google Scholar] [CrossRef] [PubMed]
- Mhalhel, K.; Germanà, A.; Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Montalbano, G. The Effect of Orally Supplemented Melatonin on Larval Performance and Skeletal Deformities in Farmed Gilthead Seabream (Sparus Aurata). Int. J. Mol. Sci. 2020, 21, 9597. [Google Scholar] [CrossRef] [PubMed]
- Mhalhel, K.; Levanti, M.; Abbate, F.; Laurà, R.; Guerrera, M.C.; Aragona, M.; Porcino, C.; Pansera, L.; Sicari, M.; Cometa, M.; et al. Skeletal Morphogenesis and Anomalies in Gilthead Seabream: A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 16030. [Google Scholar] [CrossRef] [PubMed]
- Porcino, C.; Mhalhel, K.; Briglia, M.; Cometa, M.; Guerrera, M.C.; Germanà, P.G.; Montalbano, G.; Levanti, M.; Laurà, R.; Abbate, F.; et al. Neurotrophins and Trk Neurotrophin Receptors in the Retina of Adult Killifish (Nothobranchius Guentheri). Int. J. Mol. Sci. 2024, 25, 2732. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Mhalhel, K.; Briglia, M.; Levanti, M.; Abbate, F.; Guerrera, M.C.; D’Alessandro, E.; Laurà, R.; Germanà, A. Zebrafish and Flavonoids: Adjuvants against Obesity. Molecules 2021, 26, 3014. [Google Scholar] [CrossRef] [PubMed]
- Porcino, C.; Briglia, M.; Aragona, M.; Mhalhel, K.; Laurà, R.; Levanti, M.; Abbate, F.; Montalbano, G.; Germanà, G.; Lauriano, E.R.; et al. Potential Neuroprotective Role of Calretinin-N18 and Calbindin-D28k in the Retina of Adult Zebrafish Exposed to Different Wavelength Lights. Int. J. Mol. Sci. 2023, 24, 1087. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.; Oliveri, A.; Levin, E.D. Zebrafish Model Systems for Developmental Neurobehavioral Toxicology. Birth Defects Res. Part C Embryo Toda Rev. 2013, 99, 14–23. [Google Scholar] [CrossRef]
- Schmitt, C.; McManus, M.; Kumar, N.; Awoyemi, O.; Crago, J. Comparative Analyses of the Neurobehavioral, Molecular, and Enzymatic Effects of Organophosphates on Embryo-Larval Zebrafish (Danio Rerio). Neurotoxicol. Teratol. 2019, 73, 67–75. [Google Scholar] [CrossRef]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish Disease Models in Drug Discovery: From Preclinical Modelling to Clinical Trials. Nat. Rev. Drug. Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef] [PubMed]
- DuTeaux, S.; Koshlukova, S.E. Chlorpyrifos. In Encyclopedia of Toxicology, 4th ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2024; pp. 1–13. ISBN 978-0-323-85434-4. [Google Scholar]
- Li, A.A.; Lowe, K.A.; McIntosh, L.J.; Mink, P.J. Evaluation of Epidemiology and Animal Data for Risk Assessment: Chlorpyrifos Developmental Neurobehavioral Outcomes. J. Toxicol. Environ. Health Part B 2012, 15, 109–184. [Google Scholar] [CrossRef] [PubMed]
- Sledge, D.; Yen, J.; Morton, T.; Dishaw, L.; Petro, A.; Donerly, S.; Linney, E.; Levin, E.D. Critical Duration of Exposure for Developmental Chlorpyrifos-Induced Neurobehavioral Toxicity. Neurotoxicol. Teratol. 2011, 33, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Perez-Fernandez, C.; Morales-Navas, M.; Guardia-Escote, L.; Colomina, M.T.; Giménez, E.; Sánchez-Santed, F. Postnatal Exposure to Low Doses of Chlorpyrifos Induces Long-Term Effects on 5C-SRTT Learning and Performance, Cholinergic and GABAergic Systems and BDNF Expression. Exp. Neurol. 2020, 330, 113356. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.D.; Todd, S.W.; Lumsden, E.; Mullins, R.J.; Mamczarz, J.; Fawcett, W.P.; Gullapalli, R.P.; Randall, W.R.; Pereira, E.F.R.; Albuquerque, E.X. Developmental Neurotoxicity of the Organophosphorus Insecticide Chlorpyrifos: From Clinical Findings to Preclinical Models and Potential Mechanisms. J. Neurochem. 2017, 142 (Suppl. 2), 162–177. [Google Scholar] [CrossRef] [PubMed]
- van Melis, L.V.J.; Heusinkveld, H.J.; Langendoen, C.; Peters, A.; Westerink, R.H.S. Organophosphate Insecticides Disturb Neuronal Network Development and Function via Non-AChE Mediated Mechanisms. NeuroToxicology 2023, 94, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; He, B.; Wei, Y.; Cui, J.; Zhang, Q.; Liu, X.; Liu, D.; Wang, P.; Zhou, Z. The Toxic Effects of Combined Exposure of Chlorpyrifos and p, p’-DDE to Zebrafish (Danio Rerio) and Tissue Bioaccumulation. Aquat. Toxicol. 2022, 248, 106194. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Canela, C.; Prats, E.; Piña, B.; Tauler, R. Assessment of Chlorpyrifos Toxic Effects in Zebrafish (Danio Rerio) Metabolism. Environ. Pollut. 2017, 220, 1231–1243. [Google Scholar] [CrossRef]
- Asif, M.; Shafaei, A.; Jafari, S.F.; Mohamed, S.K.; Ezzat, M.O.; Abdul Majid, A.S.; Oon, C.E.; Petersen, S.H.; Kono, K.; Abdul Majid, A.M.S. Isoledene from Mesua Ferrea Oleo-Gum Resin Induces Apoptosis in HCT 116 Cells through ROS-Mediated Modulation of Multiple Proteins in the Apoptotic Pathways: A Mechanistic Study. Toxicol. Lett. 2016, 257, 84–96. [Google Scholar] [CrossRef]
- Dawidar, A.M.; Mogib, M.A.; El-Ghorab, A.H.; Mahfouz, M.; Elsaid, F.G.; Hussien, K. Chemical Composition and Effect of Photo-Oxygenation on Biological Activities of Egyptian Commercial Anise and Fennel Essential Oils. J. Essent. Oil Bear. Plants 2008, 11, 124–136. [Google Scholar] [CrossRef]
- Chang, K.-M.; Kim, G.-H. Constituents of the Essential Oil from Eclipta prostrata L. J. Food Sci. Nutr. 2009, 14, 168–171. [Google Scholar] [CrossRef]
- Mukherjee, P.K. Chapter 7—Bioactive Phytocomponents and Their Analysis. In Quality Control and Evaluation of Herbal Drugs; Mukherjee, P.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 237–328. ISBN 978-0-12-813374-3. [Google Scholar]
- Micić, D.; Đurović, S.; Riabov, P.; Tomić, A.; Šovljanski, O.; Filip, S.; Tosti, T.; Dojčinović, B.; Božović, R.; Jovanović, D.; et al. Rosemary Essential Oils as a Promising Source of Bioactive Compounds: Chemical Composition, Thermal Properties, Biological Activity, and Gastronomical Perspectives. Foods 2021, 10, 2734. [Google Scholar] [CrossRef] [PubMed]
- Radulović, N.S.; Zlatković, D.B.; Randjelović, P.J.; Stojanović, N.M.; Novaković, S.B.; Akhlaghi, H. Chemistry of Spices: Bornyl 4-Methoxybenzoate from Ferula Ovina (Boiss.) Boiss. (Apiaceae) Induces Hyperalgesia in Mice. Food Funct. 2013, 4, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-I.; Choi, J.-H.; Kwon, T.-W.; Jo, H.-S.; Kim, D.-G.; Ko, S.-G.; Song, G.J.; Cho, I.-H. Neuroprotective Effects of Bornyl Acetate on Experimental Autoimmune Encephalomyelitis via Anti-Inflammatory Effects and Maintaining Blood-Brain-Barrier Integrity. Phytomedicine 2023, 112, 154569. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhong, X.; Zhou, Z.; Wang, N.; Deng, M. Neuroprotective Effects of Bornyl Acetate against Okadaic Acid–Induced Cytotoxicity in Pc12 Cells via Beclin-1-Dependent Autophagy Pathway. Pharmacogn. Mag. 2022, 18, 962. [Google Scholar]
- Huang, Q.; Chen, C.; Zhang, Z.; Xue, Q. Anti-Inflammatory Effects of Myristic Acid Mediated by the NF-κB Pathway in Lipopolysaccharide-Induced BV-2 Microglial Cells. Mol. Omics 2023, 19, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.-C.; Choi, H.S.; Kim, S.-L.; Lee, D.-S. 13-Oxo-9Z,11E-Octadecadienoic Acid Down-Regulates c-Myc and Attenuates Breast Cancer Cell Stemness. Vivo 2023, 37, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.-C.; Choi, H.S.; Kim, S.-L.; Yun, B.-S.; Lee, D.-S. Anti-Inflammatory Effects of (9Z,11E)-13-Oxooctadeca-9,11-Dienoic Acid (13-KODE) Derived from Salicornia herbacea L. on Lipopolysaccharide-Stimulated Murine Macrophage via NF-kB and MAPK Inhibition and Nrf2/HO-1 Signaling Activation. Antioxidants 2022, 11, 180. [Google Scholar] [CrossRef] [PubMed]
- Wójtowicz, S.; Strosznajder, A.K.; Jeżyna, M.; Strosznajder, J.B. The Novel Role of PPAR Alpha in the Brain: Promising Target in Therapy of Alzheimer’s Disease and Other Neurodegenerative Disorders. Neurochem. Res. 2020, 45, 972–988. [Google Scholar] [CrossRef]
- Kim, Y.; Hirai, S.; Goto, T.; Ohyane, C.; Takahashi, H.; Tsugane, T.; Konishi, C.; Fujii, T.; Inai, S.; Iijima, Y.; et al. Potent PPARα Activator Derived from Tomato Juice, 13-Oxo-9,11-Octadecadienoic Acid, Decreases Plasma and Hepatic Triglyceride in Obese Diabetic Mice. PLoS ONE 2012, 7, e31317. [Google Scholar] [CrossRef]
- Cai, T.; Cai, B. Pharmacological Activities of Esculin and Esculetin: A Review. Medicine 2023, 102, e35306. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, X.; Qin, S.; Zhou, S.; Li, J.; Gao, Y. Esculin Ameliorates Cognitive Impairment in Experimental Diabetic Nephropathy and Induces Anti-Oxidative Stress and Anti-Inflammatory Effects via the MAPK Pathway. Mol. Med. Rep. 2018, 17, 7395–7402. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.-L.; Zou, L.-B.; Lin, S.; Shi, J.-G.; Zhu, H.-B. Anti-Apoptotic Effect of Esculin on Dopamine-Induced Cytotoxicity in the Human Neuroblastoma SH-SY5Y Cell Line. Neuropharmacology 2007, 53, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Daroi, P.A.; Dhage, S.N.; Juvekar, A.R. P-Coumaric Acid Mitigates Lipopolysaccharide Induced Brain Damage via Alleviating Oxidative Stress, Inflammation and Apoptosis. J. Pharm. Pharmacol. 2022, 74, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-R.; Kim, M.-J.; Choi, E.-J.; Kim, Y.; Lee, H.-S.; Bae, D.; Choi, C. Protective Effects of P-Coumaric Acid Isolated from Vaccinium Bracteatum Thunb. Leaf Extract on Corticosterone-Induced Neurotoxicity in SH-SY5Y Cells and Primary Rat Cortical Neurons. Processes 2021, 9, 869. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Liu, K.; He, Q.; Sun, C.; Han, J.; Han, L.; Tian, Q. Xiaoaiping Induces Developmental Toxicity in Zebrafish Embryos Through Activation of ER Stress, Apoptosis and the Wnt Pathway. Front. Pharmacol. 2018, 9, 1250. [Google Scholar] [CrossRef] [PubMed]
- Waghmare, S.G.; Samarin, A.M.; Franěk, R.; Pšenička, M.; Policar, T.; Linhart, O.; Samarin, A.M. Oocyte Ageing in Zebrafish Danio Rerio (Hamilton, 1822) and Its Consequence on the Viability and Ploidy Anomalies in the Progeny. Animals 2021, 11, 912. [Google Scholar] [CrossRef]
- Yu, K.; Li, G.; Feng, W.; Liu, L.; Zhang, J.; Wu, W.; Xu, L.; Yan, Y. Chlorpyrifos Is Estrogenic and Alters Embryonic Hatching, Cell Proliferation and Apoptosis in Zebrafish. Chem.-Biol. Interact. 2015, 239, 26–33. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, Z.; Peng, T.; Fu, Z. The Toxicity of Chlorpyrifos on the Early Life Stage of Zebrafish: A Survey on the Endpoints at Development, Locomotor Behavior, Oxidative Stress and Immunotoxicity. Fish Shellfish. Immunol. 2015, 43, 405–414. [Google Scholar] [CrossRef]
- Levin, E.D.; Addy, N.; Nakajima, A.; Christopher, N.C.; Seidler, F.J.; Slotkin, T.A. Persistent Behavioral Consequences of Neonatal Chlorpyrifos Exposure in Rats. Dev. Brain Res. 2001, 130, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish Provide a Sensitive Model of Persisting Neurobehavioral Effects of Developmental Chlorpyrifos Exposure: Comparison with Nicotine and Pilocarpine Effects and Relationship to Dopamine Deficits. Neurotoxicol. Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.-J.; Lee, Y.-H.; Kim, M.-J.; Choi, S.-D.; Park, B.-J.; Lee, S.-E. Integrated Biomarkers Induced by Chlorpyrifos in Two Different Life Stages of Zebrafish (Danio Rerio) for Environmental Risk Assessment. Environ. Toxicol. Pharmacol. 2016, 43, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Zhang, W.; Li, L.; Jia, L.; Zhao, J.; Zhao, Z.; Peng, S.; Yuan, X.; Chen, Y. Individual and Synergistic Toxic Effects of Carbendazim and Chlorpyrifos on Zebrafish Embryonic Development. Chemosphere 2021, 280, 130769. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ren, F.; Li, Y.; Luo, J.; Pang, G. Chlorpyrifos Induces Metabolic Disruption by Altering Levels of Reproductive Hormones. J. Agric. Food Chem. 2019, 67, 10553–10562. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S.S.; Torell, F.; Donten, M.; Lundstedt-Enkel, K.; Bennett, K.; Rännar, S.; Trygg, J.; Lundstedt, T. Metabolic Profiling of Zebrafish Embryo Development from Blastula Period to Early Larval Stages. PLoS ONE 2019, 14, e0213661. [Google Scholar] [CrossRef] [PubMed]
- Macirella, R.; Curcio, V.; Brunelli, E. Morpho-Functional Alterations in the Gills of a Seawater Teleost, the Ornate Wrasse (Thalassoma pavo L.), after Short-Term Exposure to Chlorpyrifos. Toxics 2020, 8, 97. [Google Scholar] [CrossRef]
- Oğuz, A.R.; Kaval Oğuz, E.; Özok, N. Effects of Chlorpyrifos on Primary Gill Cell Culture of Lake Van Fish (Alburnus Tarichi Güldenstaadt 1814). Toxicol. Res. 2020, 9, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Sutradhar, L.; Sarker, T.R.; Saha, S.; Iqbal, M.M. Toxic Effects of Chlorpyrifos on the Growth, Hematology, and Different Organs Histopathology of Nile Tilapia, Oreochromis Niloticus. Saudi J. Biol. Sci. 2022, 29, 103316. [Google Scholar] [CrossRef]
- Zoupa, M.; Machera, K. Zebrafish as an Alternative Vertebrate Model for Investigating Developmental Toxicity—The Triadimefon Example. Int. J. Mol. Sci. 2017, 18, 817. [Google Scholar] [CrossRef]
- Jeon, H.-J.; Park, J.; Lee, S.-E. Developmental Toxicity of Chlorpyrifos-Methyl and Its Primary Metabolite, 3,5,6-Trichloro-2-Pyridinol to Early Life Stages of Zebrafish (Danio Rerio). Ecotoxicol. Environ. Saf. 2023, 249, 114352. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, J.; Avila-Barnard, S.; Nemarugommula, C.; Lyons, D.; Zhang, S.; Stapleton, H.M.; Volz, D.C. Triphenyl Phosphate-Induced Pericardial Edema in Zebrafish Embryos Is Dependent on the Ionic Strength of Exposure Media. Environ. Int. 2023, 172, 107757. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, J.; Xia, Y.; Tang, K.; Xu, J.; Wang, A.; Hu, S.; Wen, L.; Wang, B.; Yao, W.; et al. Toxic Effects of Isofenphos-Methyl on Zebrafish Embryonic Development. Ecotoxicol. Environ. Saf. 2023, 254, 114723. [Google Scholar] [CrossRef] [PubMed]
- Hedgpeth, B.M.; Redman, A.D.; Alyea, R.A.; Letinski, D.J.; Connelly, M.J.; Butler, J.D.; Zhou, H.; Lampi, M.A. Analysis of Sublethal Toxicity in Developing Zebrafish Embryos Exposed to a Range of Petroleum Substances. Environ. Toxicol. Chem. 2019, 38, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Chandra Sekaran, S.P.; Thotakura, B.; Jyothi, A.K.; Manickam, S.; Chanemougavally, J.; Prabhu, K.; Gopalan, D.H. Effect of Chlorpyrifos and Its Metabolites on Skeletal System Development of Chick Embryo. Birth Defects Res. 2023, 115, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Gao, X.; Yu, L.X.; Paria, N.; Henkelman, R.M.; Wise, C.A.; Ciruna, B. Ptk7 Mutant Zebrafish Models of Congenital and Idiopathic Scoliosis Implicate Dysregulated Wnt Signalling in Disease. Nat. Commun. 2014, 5, 4777. [Google Scholar] [CrossRef]
- Özdemir, S.; Altun, S.; Özkaraca, M.; Ghosi, A.; Toraman, E.; Arslan, H. Cypermethrin, Chlorpyrifos, Deltamethrin, and Imidacloprid Exposure up-Regulates the mRNA and Protein Levels of Bdnf and c-Fos in the Brain of Adult Zebrafish (Danio Rerio). Chemosphere 2018, 203, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-López, L.; Vega-Rivera, N.M.; Babu, H.; Ramírez-Rodríguez, G.B. Brain-Derived Neurotrophic Factor Induces Cell Survival and the Migration of Murine Adult Hippocampal Precursor Cells During Differentiation In Vitro. Neurotox. Res. 2017, 31, 122–135. [Google Scholar] [CrossRef]
- Park, H.; Poo, M. Neurotrophin Regulation of Neural Circuit Development and Function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Chen, H.; Dang, Y.; Liu, X.; Ren, J.; Wang, H. Exogenous Brain-Derived Neurotrophic Factor Attenuates Neuronal Apoptosis and Neurological Deficits after Subarachnoid Hemorrhage in Rats. Exp. Ther. Med. 2019, 18, 3837–3844. [Google Scholar] [CrossRef]
- Rahmati, M.; Keshvari, M.; Xie, W.; Yang, G.; Jin, H.; Li, H.; Chehelcheraghi, F.; Li, Y. Resistance Training and Urtica Dioica Increase Neurotrophin Levels and Improve Cognitive Function by Increasing Age in the Hippocampus of Rats. Biomed. Pharmacother. 2022, 153, 113306. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Ray, R.S.; Sharma, A.; Mehta, V.; Katyal, A.; Udayabanu, M. Antidepressant and Anxiolytic like Effects of Urtica Dioica Leaves in Streptozotocin Induced Diabetic Mice. Metab. Brain Dis. 2018, 33, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Khare, V.; Kushwaha, P.; Verma, S.; Gupta, A.; Srivastava, S.; Rawat, A.K.S. Pharmacognostic Evaluation and Antioxidant Activity of Urtica dioica L. Chin. Med. 2012, 3, 22709. [Google Scholar] [CrossRef]
- Namazi, F.; Bordbar, E.; Bakhshaei, F.; Nazifi, S. The Effect of Urtica Dioica Extract on Oxidative Stress, Heat Shock Proteins, and Brain Histopathology in Multiple Sclerosis Model. Physiol. Rep. 2022, 10, e15404. [Google Scholar] [CrossRef] [PubMed]
- Reiss, R.; Neal, B.; Lamb, J.C.; Juberg, D.R. Acetylcholinesterase Inhibition Dose–Response Modeling for Chlorpyrifos and Chlorpyrifos-Oxon. Regul. Toxicol. Pharmacol. 2012, 63, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Mit, C.; Tebby, C.; Gueganno, T.; Bado-Nilles, A.; Beaudouin, R. Modeling Acetylcholine Esterase Inhibition Resulting from Exposure to a Mixture of Atrazine and Chlorpyrifos Using a Physiologically-Based Kinetic Model in Fish. Sci. Total Environ. 2021, 773, 144734. [Google Scholar] [CrossRef] [PubMed]
- Biosca-Brull, J.; Guardia-Escote, L.; Blanco, J.; Basaure, P.; Cabré, M.; Sánchez-Santed, F.; Domingo, J.L.; Colomina, M.T. Prenatal, but Not Postnatal Exposure to Chlorpyrifos Affects Social Behavior of Mice and the Excitatory-Inhibitory Balance in a Sex-Dependent Manner. Food Chem. Toxicol. 2022, 169, 113423. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.G.; Boareto, A.C.; Schreiber, A.K.; Redivo, D.D.B.; Gambeta, E.; Vergara, F.; Morais, H.; Zanoveli, J.M.; Dalsenter, P.R. Chlorpyrifos Induces Anxiety-like Behavior in Offspring Rats Exposed during Pregnancy. Neurosci. Lett. 2017, 641, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Chrysanthis, E.; Yacisin, K.; Linney, E. Chlorpyrifos Exposure of Developing Zebrafish: Effects on Survival and Long-Term Effects on Response Latency and Spatial Discrimination. Neurotoxicol. Teratol. 2003, 25, 51–57. [Google Scholar] [CrossRef]
- Kavitha, P.; Rao, J.V. Toxic Effects of Chlorpyrifos on Antioxidant Enzymes and Target Enzyme Acetylcholinesterase Interaction in Mosquito Fish, Gambusia Affinis. Env. Toxicol. Pharmacol. 2008, 26, 192–198. [Google Scholar] [CrossRef]
- Halappa, R.; David, M. Behavioural Responses of the Freshwater Fish, Cyprinus Carpio (Linnaeus) Following Sublethal Exposure to Chlorpyrifos. Turk. J. Fish. Aquat. Sci. 2009, 9, 233–238. [Google Scholar] [CrossRef]
- Stalin, A.; Suganthi, P.; Mathivani, S.; Paray, B.A.; Al-Sadoon, M.K.; Gokula, V.; Musthafa, M.S. Impact of Chlorpyrifos on Behavior and Histopathological Indices in Different Tissues of Freshwater Fish Channa Punctatus (Bloch). Environ. Sci. Pollut. Res. 2019, 26, 17623–17631. [Google Scholar] [CrossRef] [PubMed]
- Izunwanne, D.; Mobisson, S.; Nwafor, C.; Ifedi, I.; Onwukaike, C.; Izunwanne, H. Excitatory Effect of Urtica Dioica on Locomotor Behaviour of Mice Using the Open Field Maze Task. JOCAMR 2024, 25, 34–42. [Google Scholar] [CrossRef]
- Aragona, M.; Porcino, C.; Guerrera, M.C.; Montalbano, G.; Laurà, R.; Cometa, M.; Levanti, M.; Abbate, F.; Cobo, T.; Capitelli, G.; et al. The BDNF/TrkB Neurotrophin System in the Sensory Organs of Zebrafish. Int. J. Mol. Sci. 2022, 23, 2621. [Google Scholar] [CrossRef]
- Germanà, A.; Guerrera, M.C.; Laurà, R.; Levanti, M.; Aragona, M.; Mhalhel, K.; Germanà, G.; Montalbano, G.; Abbate, F. Expression and Localization of BDNF/TrkB System in the Zebrafish Inner Ear. Int. J. Mol. Sci. 2020, 21, 5787. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tentative Assignment | Formula | [M-H] + Found | [M-H] − Found | RDB |
|---|---|---|---|---|
| Isoledene | C15H24 | 219.17525 | - | 4.5 |
| Hexadecanethiol | C16H34S | 256.13380 | - | 4 |
| Heptadecenoic acid | C17H34O2 | 278.07987 | - | 8 |
| Ethylene glycol | C18H38O4 | 318.30066 | - | 0 |
| Bornyl acetate | C12H20O2 | 153.13930 | - | 2.5 |
| Oxo-octadecadienoic acid | C18 H3003 | - | 293.17831 | 4.5 |
| Esculin | C15 H1609 | - | 339.19858 | 7.5 |
| p-Coumaric acid | C15 H1808 | - | 325.18317 | 7.5 |
| Antibody | Dilution | |
|---|---|---|
| Primary antibodies | Anti-brain-derived neurotrophic factor (Cat. # AB1534SP, Merck Millipore, Burlington, MA, USA) | [1:100] |
| Secondary antibodies | Goat anti-rabbit IgG (H + L) cross-adsorbed secondary antibody, Alexa Fluor™ 594 (Thermo Fisher Scientific, Chino, CA, USA, Cat. # A-11012) | [1:100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mhalhel, K.; Kadmi, Y.; Ben Chira, A.; Levanti, M.; Pansera, L.; Cometa, M.; Sicari, M.; Germanà, A.; Aragona, M.; Montalbano, G. Urtica dioica Extract Abrogates Chlorpyrifos-Induced Toxicity in Zebrafish Larvae. Int. J. Mol. Sci. 2024, 25, 6631. https://doi.org/10.3390/ijms25126631
Mhalhel K, Kadmi Y, Ben Chira A, Levanti M, Pansera L, Cometa M, Sicari M, Germanà A, Aragona M, Montalbano G. Urtica dioica Extract Abrogates Chlorpyrifos-Induced Toxicity in Zebrafish Larvae. International Journal of Molecular Sciences. 2024; 25(12):6631. https://doi.org/10.3390/ijms25126631
Chicago/Turabian StyleMhalhel, Kamel, Yassine Kadmi, Ahlem Ben Chira, Maria Levanti, Lidia Pansera, Marzio Cometa, Mirea Sicari, Antonino Germanà, Marialuisa Aragona, and Giuseppe Montalbano. 2024. "Urtica dioica Extract Abrogates Chlorpyrifos-Induced Toxicity in Zebrafish Larvae" International Journal of Molecular Sciences 25, no. 12: 6631. https://doi.org/10.3390/ijms25126631