Abstract
Phytochemicals and tryptophan (Trp) metabolites have been found to modulate gut function and health. However, whether these metabolites modulate gut ion transport and serotonin (5-HT) metabolism and signaling requires further investigation. The aim of this study was to investigate the effects of selected phytochemicals and Trp metabolites on the ion transport and 5-HT metabolism and signaling in the ileum of mice in vitro using the Ussing chamber technique. During the in vitro incubation, vanillylmandelic acid (VMA) reduced (p < 0.05) the short-circuit current, and 100 μM chlorogenic acid (CGA) (p = 0.12) and perillic acid (PA) (p = 0.14) had a tendency to reduce the short-circuit current of the ileum. Compared with the control, PA and N-acetylserotonin treatment upregulated the expression of tryptophan hydroxylase 1 (Tph1), while 100 μM cinnamic acid, indolelactic acid (ILA), and 10 μM CGA or indoleacetaldehyde (IAld) treatments downregulated (p < 0.05) the mRNA levels of Tph1. In addition, 10 μM IAld or 100 μM ILA upregulated (p < 0.05) the expression of monoamine oxidase A (Maoa). However, 10 μM CGA or 100 μM PA downregulated (p < 0.05) Maoa expression. All selected phytochemicals and Trp metabolites upregulated (p < 0.05) the expression of Htr4 and Htr7 compared to that of the control group. VMA and CGA reduced (p < 0.05) the ratios of Htr1a/Htr7 and Htr4/Htr7. These findings may help to elucidate the effects of phytochemicals and Trp metabolites on the regulation of gut ion transport and 5-HT signaling-related gut homeostasis in health and disease.
1. Introduction
The gut microbiota, together with its metabolites, has been proven to modulate host metabolism and improve the gut barrier and function [1,2]. Robust gut function is essential for maintaining whole-body nutrition and health. The small intestine is crucial for nutrient absorption and immunity; however, these processes are affected by the gut microbiota, which has attracted much attention in recent years [3]. It has been shown that tryptophan (Trp) metabolites regulate the gut barrier and immune response [4,5]. Indoleacetic acid and indole-3-aldehyde alleviate gut inflammation by decreasing lipopolysaccharide-induced production of the proinflammatory cytokines tumor necrosis factor alpha (TNF-α) and interleukin (IL)-1β in macrophages and promoting the production of IL-22 and the expression of tight junction proteins in the colon of mice [6,7,8]. In addition, a study with aryl hydrocarbon receptor (AHR)−/− mice suggested that indole-3-ethanol and indole-3-pyruvate regulated gut barrier function in an AHR-signaling dependent manner [9]. However, other mechanisms underlying the regulatory role of Trp metabolites in small intestine function require further investigation.
Moreover, phytochemicals, which are naturally bioactive compounds in plants, have attracted great attention because of their health-promoting benefits, including antioxidant, immunomodulatory, and antibacterial activities and the ability to increase the efficiency of nutrient digestion and absorption [10,11,12]. Perillic acid, a major metabolite of D-limonene, regulates the body immune response by increasing the total numbers of white blood cells and promotes the production of the antioxidative compound glutathione in the small intestine of mice [13,14,15]. Curcumin protected against oxidative injury and inflammation of the gut through inhibiting the nuclear factor κ-B (NF-κB)/IκB signaling pathway, decreasing m6A RNA methylation and boosting intestinal autophagy in duckling and piglet models [16,17,18]. In addition, the phenolic compounds cinnamic acid and chlorogenic acid have been proposed to have antioxidant and anti-inflammatory properties [19]. Cinnamic acid can alleviate oxidative stress in the liver by inhibiting macrophage infiltration and downregulating the expression of proinflammatory cytokines such as TNF-α and IL-6 [20,21]. In addition, chlorogenic acid suppressed gut inflammation and reduced hepatotoxicity by downregulating the expression of the proinflammatory cytokines IL-1β, IL-6, and interferon-γ (IFN-γ) and interfering with mitochondrial function and Nrf2/HO-1 signaling [22,23]. The above findings suggested that the antioxidant and immune-modulatory capacity of the abovementioned phytochemicals can improve gut function and health through multiple mechanisms [24]. However, further studies are warranted to uncover the detailed mechanisms that regulate gut function.
Serotonin (5-HT) is an important hormone and signaling molecule that regulates gut functions, including gut motility, ion transport, fluid secretion, and immune function [25,26,27]. More than 90% of 5-HT in the body is produced in the enterochromaffin cells (EC) of the gut through the metabolism of Trp by tryptophan hydroxylase 1 (Tph1). Extracellular 5-HT can be taken up by the serotonin reuptake transporter and then degraded to 5-hydroxyindoleacetic acid by monoamine oxidase A (Maoa) [28]. A study on the link between 5-HT and gut immune function showed that 5-HT altered colitis susceptibility in Tph1−/− mice [29]. In addition, we have shown that Trp and Limosilactobacillus mucosae can alleviate dextran sulfate sodium (DSS)-induced colonic inflammation by upregulating the expression of 5-HT receptors (Htr), including Htr1a and Htr4, and reducing Htr2a and Htr7 expression [30,31]. Moreover, a study in weaned piglets revealed that there was a positive correlation between Htr7 expression and the levels of proinflammatory cytokines, including IL-1β, IL-8, and TNF-α, but a negative correlation between Htr4 expression and levels of IFN-γ in the jejunum [32]. 5-HT was found to regulate intestinal motility via the activation of Htr3 and Htr4 [33]. In addition, microbial tryptamine stimulates gut transit by stimulating epithelial HTR4, which is accompanied by increased colonic fluid secretion in mice [34]. Therefore, the combination of 5-HT and Htr has a substantial effect on gut function [35]. However, the regulation of 5-HT metabolism and signaling by dietary bioactive compounds in the small intestine and the underlying mechanisms require further investigation.
This study aimed to identify the differences in the mode of action of different phytochemicals and Trp metabolites in the regulation of 5-HT signaling and ion transport in the mouse ileum using the Ussing chamber technique. The findings of this study will aid in the development of potential dietary interventions for improving gut health.
2. Results
2.1. Effects of Different Phytochemicals and Trp Metabolites on the Short-Circuit Current of the Mouse Ileum
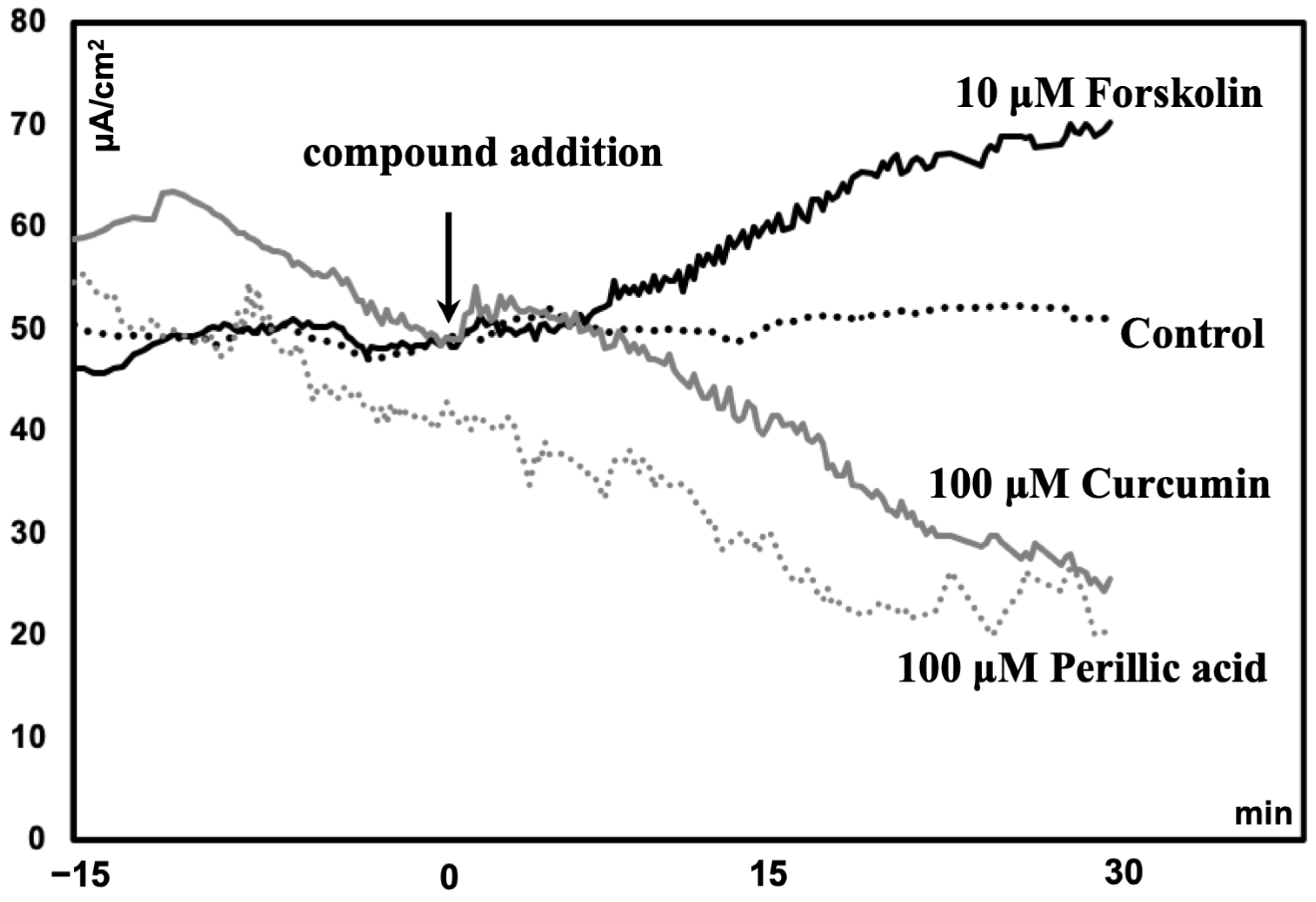
Compared to those in the control group, the ileum of mice treated with forskolin (10 μM) had greater Isc (p < 0.05). However, the VMA (100 μM) treatment group had a lower Isc (p = 0.05) than the control group (Table 1). Compared with the control, PA (100 μM) or curcumin (100 μM) tended to reduce Isc (p = 0.1) (Table 1). A representative diagram comparing the short-circuit current (Isc) following the administration of phytochemicals is shown in Figure 1.

Table 1.
Effects of different phytochemicals and Trp metabolites on the short-circuit current (Isc) across the mouse ileum 1.
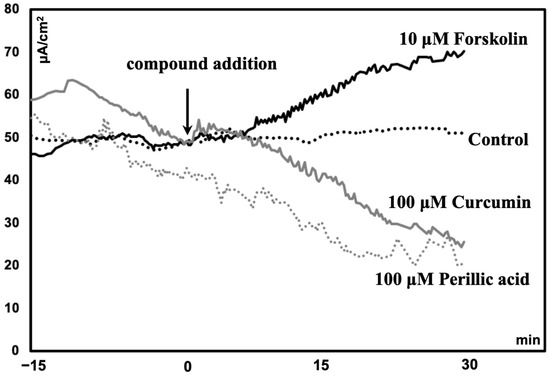
Figure 1.
Representative diagram comparing the short-circuit current (Isc) of the ileum of mice following the administration of phytochemicals.
2.2. Effects of Different Phytochemicals and Trp Metabolites on the Gene Expression and Protein Abundance of Key Enzymes Involved in 5-HT Metabolism in the Mouse Ileum
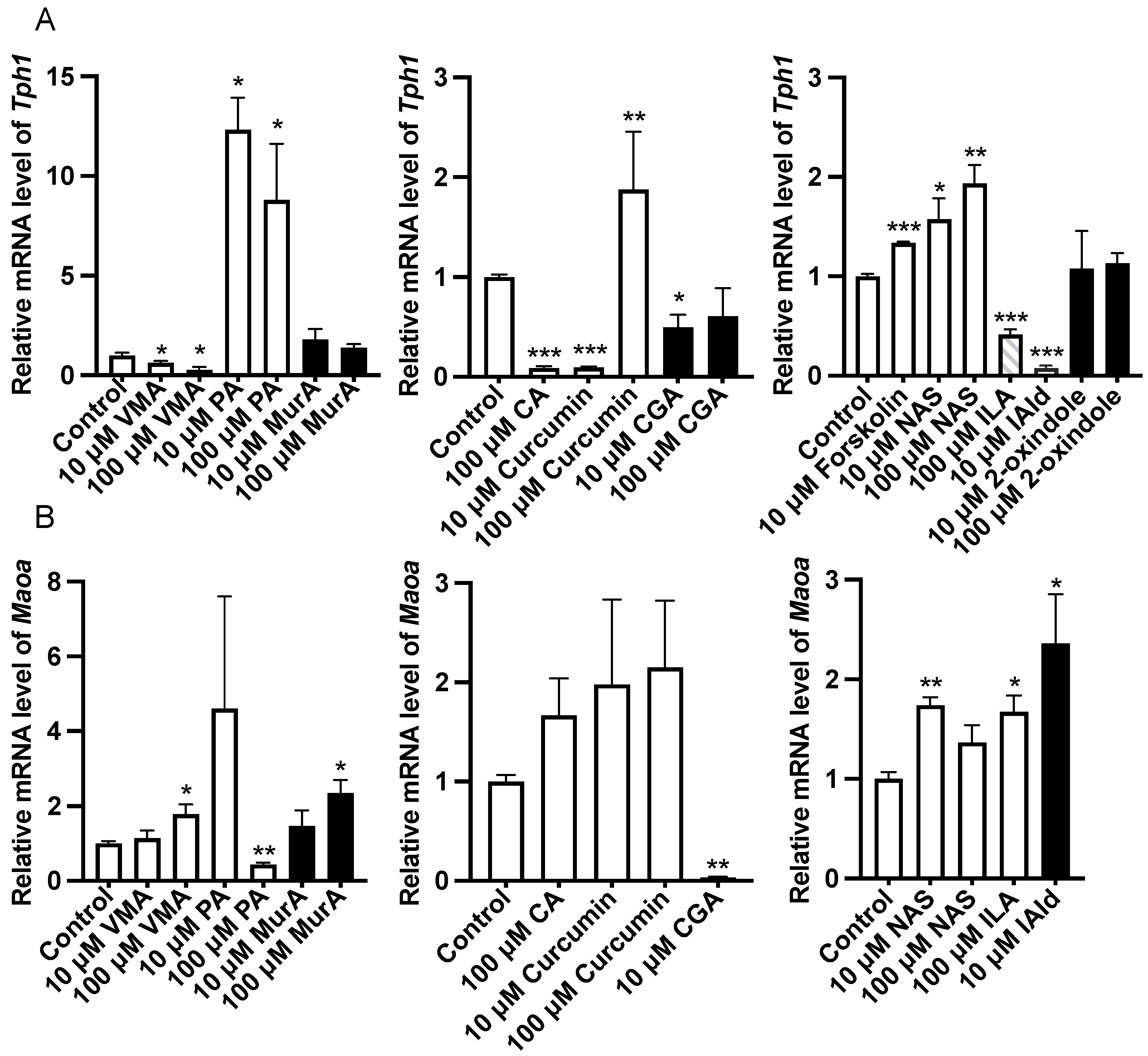
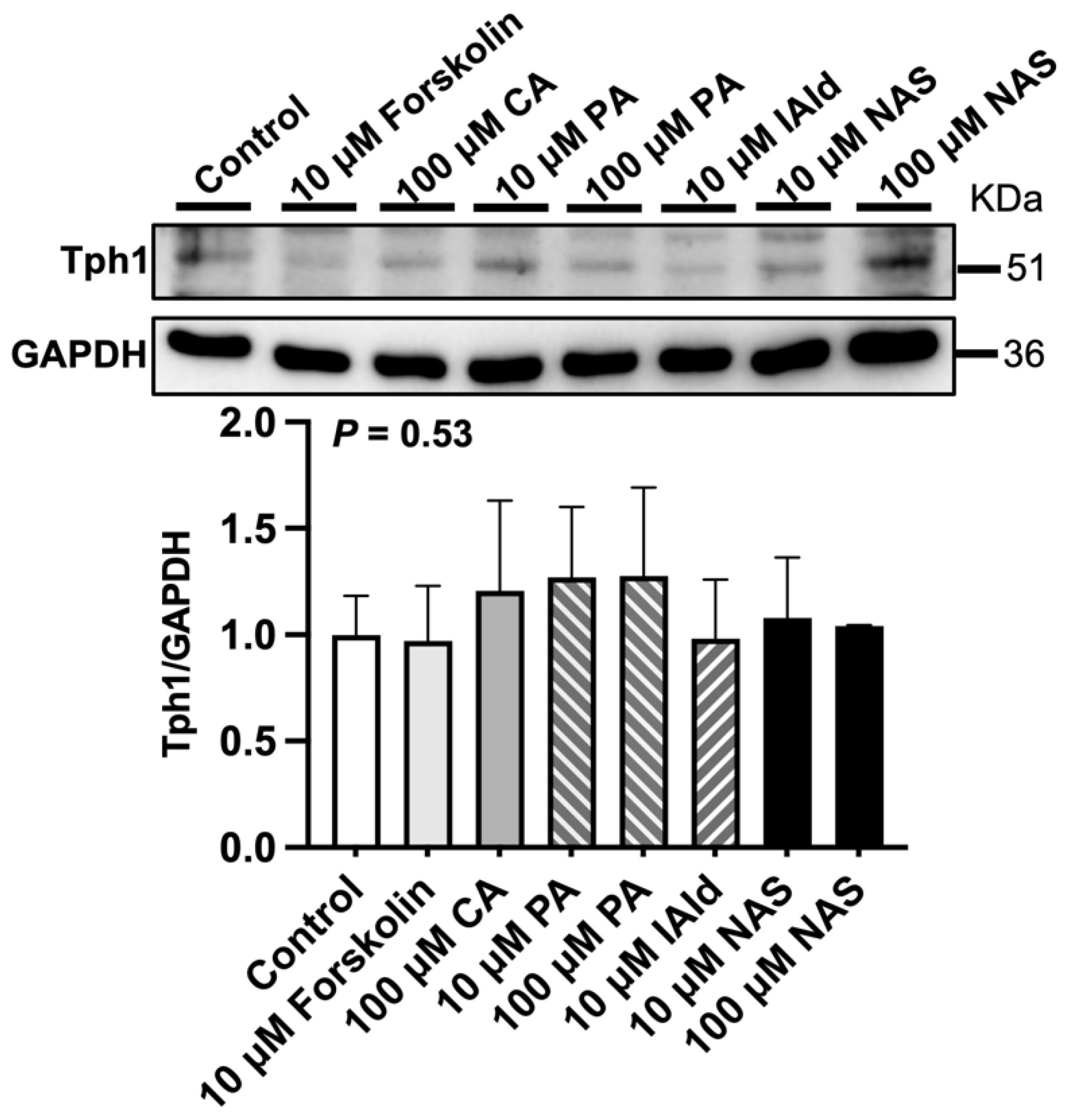
Analysis of the gene expression of 5-HT metabolism-related enzymes in the ileum revealed that forskolin (10 μM), PA or NAS treatment upregulated (p < 0.05) the expression of Tph1 compared with that in the control group (Figure 2). In addition, VMA, CA (100 μM), CGA (10 μM), ILA (100 μM), and IAld (10 μM) downregulated (p < 0.05) the expression of Tph1 (Figure 2). The protein abundance of Tph1 in the ileum was not affected by the abovementioned phytochemicals and Trp metabolites (Figure 3). In addition, 100 μM VMA, MurA, ILA or 10 μM IAld upregulated (p < 0.05) the expression of Maoa. However, PA (100 μM) and CGA (10 μM) downregulated (p < 0.05) Maoa expression (Figure 2).
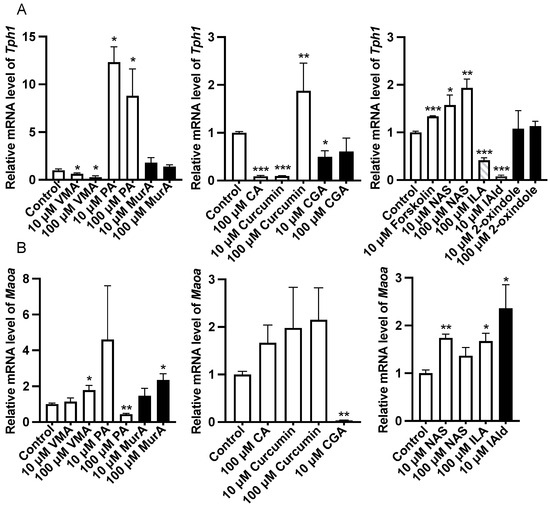
Figure 2.
Effects of different phytochemicals and Trp metabolites on the gene expression of 5-HT metabolism-related enzymes, including Tph1 (A) and Maoa (B) in the ileum of mice. The data in the charts are the means ± SEMs. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control group. Tph1, tryptophan hydroxylase 1; Maoa, monoamine oxidase A; VMA, vanillylmandelic acid; PA, perillic acid; MurA, muramic acid; CA, cinnamic acid; CGA, chlorogenic acid; NAS, N-acetylserotonin; ILA, indolelactic acid; IAld, indoleacetaldehyde. The number of animals is shown in Table 1.
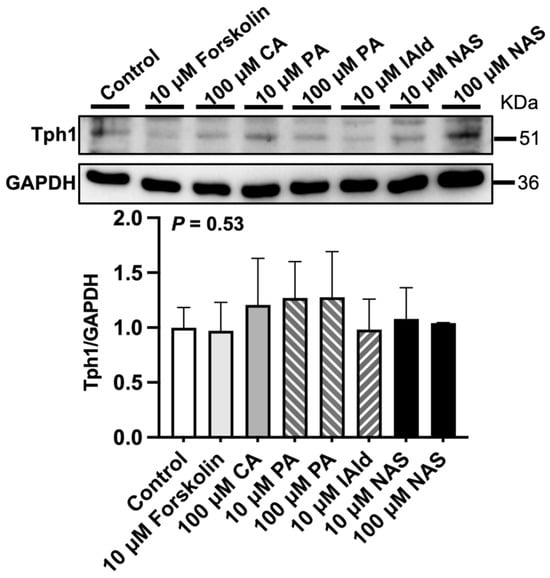
Figure 3.
Effects of different phytochemicals and Trp metabolites on the protein abundance of tryptophan hydroxylase 1 (Tph1) in the ileum of mice. The data in the charts are the means ± SEMs, n = 3. CA, cinnamic acid; PA, perillic acid; IAld, indoleacetaldehyde; NAS, N-acetylserotonin.
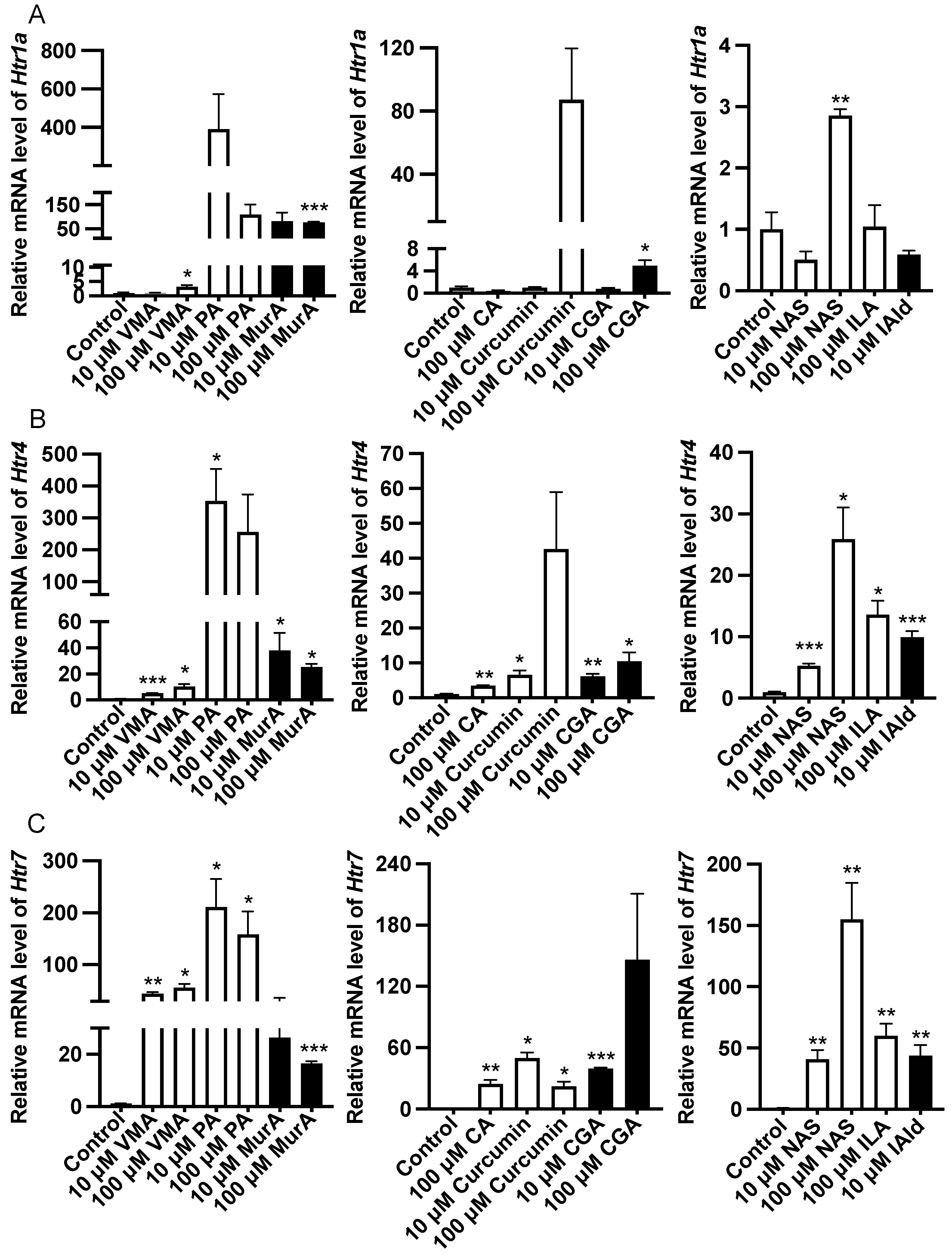
2.3. Effects of Different Phytochemicals and Trp Metabolites on the Gene Expression of Htr and Their Ratios in the Mouse Ileum
Analysis of the gene expression of the Htr revealed that 100 μM VMA, GA, and NAS or 10 μM MurA upregulated (p < 0.05) the expression of Htr1a (Figure 4). Treatment with 10 μM VMA, PA, MurA, curcumin, CGA, NAS or IAld or 100 μM CA or ILA upregulated (p < 0.05) the expression of Htr4 and Htr7 (Figure 4).
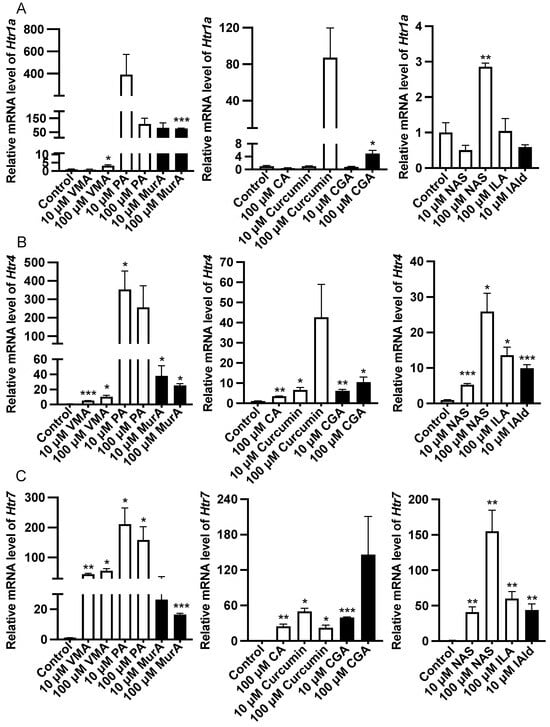
Figure 4.
Effects of different phytochemicals and Trp metabolites on the expression of serotonin receptor genes, including Htr1a (A), Htr4 (B), and Htr7 (C) in the ileum of mice. The data in the charts are the means ± SEMs. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control group. The number of animals is shown in Table 1. VMA, vanillylmandelic acid; PA, perillic acid; MurA, muramic acid; CA, cinnamic acid; CGA, chlorogenic acid; NAS, N-acetylserotonin; ILA, indolelactic acid; IAld, indoleacetaldehyde.
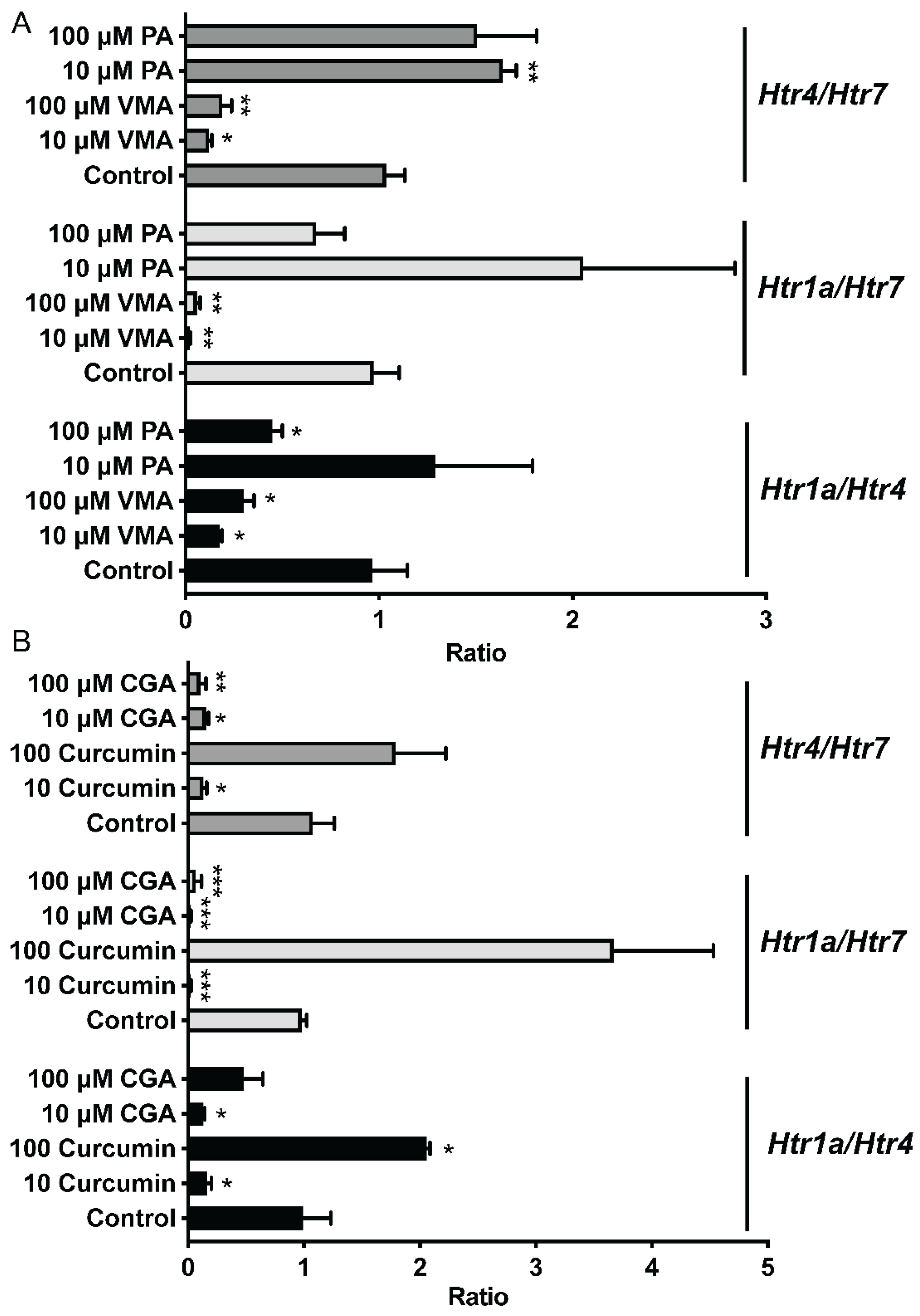
To further compare the effects of selected phytochemicals and Trp metabolites on the expression of Htr, the expression ratios of Htr1a/Htr7, Htr4/Htr7, and Htr4/Htr7 were calculated and analyzed based on their relative expression to that of Gapdh (Figure 5). Compared to those in the control group, the Htr1a/Htr4 and Htr4/Htr7 ratios in the curcumin (100 μM) and PA (10 μM) groups were greater (p < 0.05). However, the ileum treated with 10 μM or 100 μM VMA or CGA and 10 μM curcumin had lower (p < 0.05) Htr1a/Htr7, Htr4/Htr7, and Htr1a/Htr4 ratios (Figure 5).
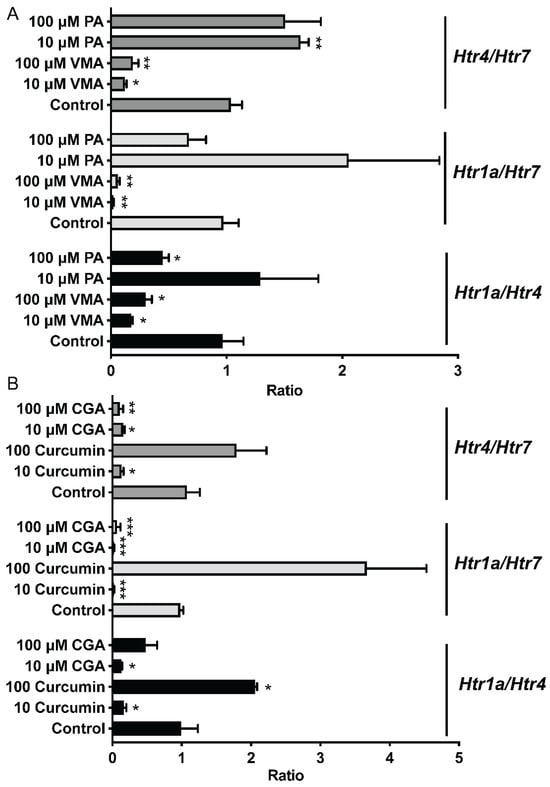
Figure 5.
Different phytochemicals, including PA and VMA (A), and CGA and Curcumin (B) on the serotonin receptor gene ratios in the ileum of mice. The data in the charts are the means ± SEMs. * p < 0.05, ** p < 0.01, and *** p < 0.001 compared to the control group. The number of animals is shown in Table 1. CGA, chlorogenic acid; PA, perillic acid; VMA, vanillylmandelic acid.
3. Discussion
Phytochemicals and Trp metabolites have been shown to improve gut function and homeostasis, and these health-promoting effects may be partially due to their regulation of gut 5-HT metabolism and signaling [35,36]. Gut barrier function is important not only for defense against luminal pathogens but also for nutrient absorption and electrolyte balance [37]. Our current study suggested that VMA reduced the short-circuit current, and 100 μM CGA and PA had a tendency to reduce the short-circuit current of the ileum. PA and NAS upregulated Tph1 expression, while CA, CGA, ILA, or IAld downregulated Tph1 expression. Moreover, PA or CGA downregulated Maoa expression. However, ILA and IAld upregulated Maoa expression. All selected phytochemicals and Trp metabolites upregulated the expression of Htr4 and Htr7; however, PA increased the Htr4/Htr7 ratio, while VMA and CGA reduced the Htr1a/Htr7 and Htr4/Htr7 ratios. These findings may help to gain further insight into the regulatory role of phytochemicals and Trp metabolites on gut 5-HT homeostasis-mediated gut function in health and disease.
The findings of our present study revealed that different Trp metabolites regulate gut 5-HT homeostasis differently. Mounting evidence has shown that the gut microbiota and its metabolites are crucial for the production of 5-HT by regulating Tph1 expression in the intestines of rodents [38]. In particular, microbial-derived Trp metabolites exert diverse effects on the host physiology and gut function, including gut immune modulation [4,39]. Microbial indole derivatives such as ILA inhibit the proliferation of macrophages by reducing glycolysis, NF-κB, and hypoxia-inducible factor pathways and decreasing CCL2/7 in epithelial cells and mice [40]. In rat and mouse models, NAS can alleviate intestinal ischemic injury and DSS-induced chronic colitis [41,42,43]. In TNF-α-stimulated epithelial cells, the addition of ILA did not alter the expression of Tph1 [44]. Additionally, microbial 5-hydroxyindole has been shown to promote 5-HT release from RIN14B cells, which regulates gut motility by activating Htr3 and Htr4 [39]. In our study, the Trp metabolite NAS upregulated the expression of Tph1, while ILA and IAld downregulated the expression of Tph1, suggesting that the regulation of 5-HT production differs among different Trp metabolites. Notably, the abovementioned regulatory role of different Trp metabolites may also be affected by different physiological conditions of the gut (i.e., normal conditions vs. inflammation) [44]. Further studies are required to determine the differential effects of various Trp metabolites on 5-HT signaling to identify new targets for dietary strategies to improve gut function.
Our current study also reinforces the importance of gut 5-HT metabolism and signaling in the anti-inflammatory effects of phytochemicals. The limonene derivative PA has been widely used in food, medicine, and cosmetics [13]. PA was proposed to be a potential anticancer, immunomodulatory, and anti-obesogenic bioactive compound [45]. However, there are very few studies on the regulatory role of PA on gut 5-HT. In mice, PA alleviated radiation-induced small intestinal histopathological damage and reduced the production of the proinflammatory cytokines IL-1β and TNF-α [15]. In addition, supplementation with 0.1% D-limonene hampered diet-induced obesity in mice with increased levels of PA and perillic acid-8,9-diol in the urine [46]. Our current results demonstrated that PA stimulated 5-HT synthesis, inhibited 5-HT degradation, and upregulated Htr4 and Htr7 expression. We therefore deduced that PA might exert its beneficial effects by affecting 5-HT metabolism and Htr, but this needs to be further confirmed in vivo. Additionally, the present findings confirmed that CGA downregulated the expression of Tph1 and Maoa, which is in line with a previous finding that CGA suppressed Maoa activity and prevented 5-HT from being deaminated [47]. In addition, studies have shown that curcumin can modulate 5-HT levels in the brain and exert antidepressant and anticonvulsive effects [48,49,50]. Additionally, curcumin has been widely reported to promote intestinal health by affecting cross-talk among cell signaling pathways, immune function, and the gut microbiota [24,51]. However, a recent study suggested that higher concentrations (20–80 μM) of curcumin exerted cytotoxic effects on human small intestine epithelial cells, which might be a compensatory protective mechanism under conditions of impaired cell vitality [52]. Notably, our current results revealed that curcumin at low and high concentrations had opposite effects on Tph1 expression in the ileum, indicating that its effect on 5-HT signaling may be dose dependent. Moreover, the levels of 5-HT were decreased in the colon of rats with irritable bowel syndrome by curcumin supplementation, but an Htr1a antagonist reversed this effect, indicating that the beneficial effect of curcumin is Htr1a dependent [53]. This finding is in line with our current finding that the Htr1a/Htr4 ratio was increased by 100 μM curcumin. Taken together, the intricate role of 5-HT and its receptors in the regulation of gut function modulated by the abovementioned compounds requires further investigation.
The expression of different subtypes of Htr in the gut plays vital roles in gut functions, including gut secretion, motility, and immune modulation [33]. Despite a great clinical interest and an increasing number of studies devoted to Htr, the mechanism modulating Htr effect on physiological and pathological functions required further validation. A previous study showed that the Htr1 and Htr7 receptors can form homo- and heterodimers [54], thus impairing the ability of Htr1 to activate the Gi-protein system and further downstream signaling pathway. Especially, a recent report found that overexpression of Htr7 led to the decreased abundance of Htr1a in the membrane protein fraction from the midbrain samples of C57BL/6 mice [55], thus exerting an antidepressive effect, which suggests the interaction and proportion of Htr played important roles in the physiology and pathology of the body. However, changes in the proportion of Htr in the gut and the effects and mechanisms of Htr interaction on gut function in gut health and disease remain poorly understood. A recent study in weaned piglets indicated that there is a correlation between the expression of Htr, including Htr4 and Htr7, and the production of inflammatory factors in the jejunum [32]. In addition, Trp upregulated Htr1a and Htr4 expression, thus mitigating DSS-induced colitis in mice [30]. Interestingly, although the inhibition of Htr1a or Htr4 by their antagonists exacerbated DSS-induced colitis in mice, the modulation of one Htr via its antagonist did not change the expression of other Htr [30]. In our current study, CGA stimulated the expression of Htr1a, Htr4, and Htr7, but the Htr1a/Htr4, Htr1a/Htr7, and Htr4/Htr7 ratios were lower than those in the control group. Hence, we speculated that the relative proportion of each Htr in the intestine may be vital for the regulation of gut homeostasis; however, more studies are warranted to test this hypothesis.
The Ussing chamber system has been extensively used for evaluating the transport of ions, nutrients, and drugs across various epithelial tissues, as well as intestinal permeability [56]. Short-circuit current (Isc) is an indicator of the tissue’s ability to absorb or secrete [57]. Our results indicated that 10 μM forskolin increases Isc significantly, and this suggested the ion transport through ileum epithelium tissue was active. However, a higher dose (100 μM) of PA and curcumin tended to decrease the Isc in the ileum of the mice, which would suggest either an inherent change in epithelial transport or a decrease in fluid secretion. In addition, activation of ion transport in the intestine can be achieved in part through the production of hormones from enteroendocrine cells. Hormones, such as 5-HT, can in turn act on mucosal nerve endings and activate secretomotor neurons by activating Htr [58,59]. Therefore, whether PA-induced Isc change is caused by the indirect 5-HT signaling effect can be explored by the addition of selective blockers of Tph1 responsible for 5-HT synthesis or Htr antagonists in future studies. Also, one limitation of our present study is that the sample size is relatively small, so more sample sizes need to be replicated in future experiments to verify the above speculation. In addition, by using an EC-enriched monolayer system, scholars found that treatment with forskolin (10 μM) and the dietary nutrient curcumin (100 μM) stimulated 5-HT production by cells [60]. This result was partly consistent with our finding that Isc and Tph1 gene expression were elevated by forskolin. However, the findings regarding the ability of curcumin to stimulate 5-HT release were inconsistent with our experimental results, as evidenced by the fact that 100 μM curcumin treatment reduced Isc and Tph1 expression in our study. This might be due to the differences in the experimental systems used in the two studies [60].
4. Materials and Methods
4.1. Reagents
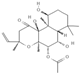
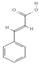
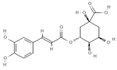
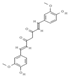
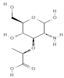
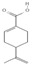
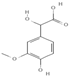
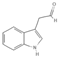
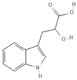
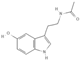
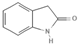
Primary antibody against Tph1 (Cat# 12339) was purchased from Cell Signaling Technology (Danvers, MA, USA). Glucose and salts were purchased from Sangon Biotech Co., Ltd. (Shanghai, China). Curcumin (#HY-N0005), forskolin (#HY-15371), indolelactic acid (ILA, #HY-113099), and 2-oxindole (#HY-Y0061) were obtained from Med Chem Express Co., Ltd. (Shanghai, China), and the purity of these reagents is more than 98%. Cinnamic acid (CA, #C80857), chlorogenic acid (CGA, #C3878), indoleacetaldehyde (IAld, #I1000), muramic acid (MurA, #M2503), N-acetylserotonin (NAS, #A1824), perillic acid (PA, #218359), and vanillylmandelic acid (VMA, #H0131) were purchased from Merck Sigma-Aldrich (Shanghai, China), and the purity is more than 95%. The compounds used in this study were dissolved in dimethyl sulfoxide (DMSO, final concentration 0.1%) and further diluted with Krebs’ solution for subsequent Ussing chamber experiments. The concentrations of the substances were selected with reference to the possible non-toxic dosages that have been reported in cell and mice experiments. Chemical structures and certain physicochemical properties of the above compounds used in this study are shown in Table 2.
Table 2.
Chemical structures and certain physicochemical properties of the selected compounds used in this study.
4.2. Animals
In this study, sixty 10~12 week-old C57BL/6 male mice with an average body weight of 27.0~30.0 g were purchased from Beijing HFK Bioscience (Beijing, China). Upon arrival, the mice were allowed to acclimate for 7 days before the experiment. The mice were housed in polycarbonate cages in a specific-pathogen-free environment and were kept at 22–25 °C and 45–55% relative humidity with a 12 h light/dark cycle and ad libitum water and feed. The standard rodent diet (Cat#1032, Beijing HFK Bioscience) used in the experiments was the same as that used in our previous study [30].
4.3. Ussing Chamber Experiments
Before Ussing chamber analysis, the mice were anesthetized with ether before they were euthanized by cervical dislocation, according to the previous method [31]. The proximal ileum was first rinsed with precooled Krebs’ solution (11.1 mM glucose, 118 mM NaCl, 4.8 mM KCl, 1.0 mM NaH2PO4, 1.2 mM MgSO4, 25 mM NaHCO3, 2.5 mM CaCl2, pH = 7.4) to remove the chyme and then opened along the mesenteric border.
A six-channel Ussing chamber system (MC6-6, Physiologic Instrument, Reno, NV, USA) was used to measure the short-circuit current (Isc) (as shown in Figure 6). The isolated ileum segments of 1.5 cm in length were then pinned onto Ussing chamber sliders (P2300, 0.2 cm2 apertures) within 15 min after sampling. After that, the sliders with the ileum were mounted into chambers (EasyMount Diffusion Chambers, Physiologic Instruments), and 5 mL of Krebs’ solution was added to the mucosal side and serosal side of the ileum. A final concentration of 5 mM mannitol was added to the mucosal side to limit active transport while maintaining osmotic balance. Ileal tissue was stabilized for 15 min before clamping the voltage to 0 V before treatment. Final concentrations of 10 μM or 100 μM of the selected compounds were added to the chamber for up to 40 min after the mounted tissues were stabilized. Forskolin (10 μM) was used as a positive control, as reported previously [61]. In this study, the number of ileal tissues tested used for each compound was equal to the number of animals used (n = 3~6). The Isc was continuously recorded by Acquire & Analyze software 2.3 (Physiologic Instruments, Reno, NV, USA). Tissue viability was assessed according to a previous method [61], and tissues with a <1 mV increase in the transepithelial potential difference were excluded from further analysis.

Figure 6.
A photo of the Ussing chamber system used in this study.
4.4. RNA Extraction and Quantitative Real-Time PCR Analysis
After Ussing chamber analysis, total RNA was extracted from the analyzed ileal tissues using an RNAiso Plus kit (Takara, Beijing, China). The integrity and purity of the RNA were determined using a Nanodrop P330 (Implen, Munich, Germany) and electrophoresis. RNA was then reverse-transcribed using a FastKing RT kit from TIANGEN Biotech Co., Ltd. (Beijing, China). Quantitative real-time PCR was performed using a SYBR Premix Ex Taq II (Takara, Beijing, China) with an ABI 7500 real-time PCR detection system (Thermo Fisher, Waltham, MA, USA). The sequences of the primers (Tph1, Maoa, Htr1a, Htr4, Htr7, and GAPDH) used for this study were described previously [30]. GAPDH was utilized as the internal reference. The 2−ΔΔCt method was used for the quantification of gene expression.
4.5. Western Blot Analysis
Ileum samples were homogenized in liquid nitrogen, and protein was extracted and measured for the abundance of Tph1 by Western blot analysis, according to a previous protocol [62]. Protein bands were measured via ECL Plus detection reagents (Thermo Fisher, Waltham, MA, USA) and visualized by the Image Quant LAS 4000 mini system (GE Healthcare, Piscataway, NJ, USA). The band intensity of each target protein was compared with that of GAPDH by Image J software version 1.53 (NIH, Bethesda, MD, USA).
4.6. Statistical Analysis
The values are presented as the means ± SEMs. Statistical differences between two groups were analyzed by t-tests. The data were assessed by one-way ANOVA followed by Duncan’s multiple comparison test. SPASS 22.0 (IBM, Armonk, NY, USA) and Prism 9.0 (GraphPad, Boston, MA, USA) were used for statistical analysis. p < 0.05 was used to indicate a significant difference.
5. Conclusions
In conclusion, 100 μM VMA or curcumin reduced Isc, and PA and NAS upregulated the gene expression of Tph1, but CGA (10 μM), IAld (10 μM), and ILA (10 μM) downregulated the gene expression of Tph1. The selected phytochemicals and Trp metabolites upregulated the expression of Htr4 and Htr7 compared to those in the control group. PA (10 μM) increased the Htr4/Htr7 ratio, while CGA decreased the Htr1a/Htr4, Htr1a/Htr7, and Htr4/Htr7 ratios. These findings may help to elucidate the regulatory role of phytochemicals and Trp metabolites in gut 5-HT signaling-mediated gut ion transport and homeostasis, thereby providing potential dietary intervention options for improving gut function and health in humans and animals.
Author Contributions
L.J. and Z.D. designed the experiments and wrote the manuscript; L.J., Y.H., Q.L. and Z.D. conducted the experiments; L.J. and Z.D. revised the manuscript; all the authors have read and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Key R&D Program of China (2022YFD1300402) and the National Natural Science Foundation of China (32072689).
Institutional Review Board Statement
All the animal experimental procedures followed the guidelines of the Institutional Animal Care and Use Committee of China Agricultural University (approval No. Aw11013202-1-2 and approval date: 11 October 2023).
Informed Consent Statement
Not Applicable.
Data Availability Statement
All data presented in the study were included in the manuscript.
Acknowledgments
The authors would like to thank Zhenlong Wu and Junjun Wang for their helpful suggestions on this work and reviewers for their constructive comments.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Koh, A.; Backhed, F. From association to causality: The role of the gut microbiota and its functional products on host metabolism. Mol. Cell 2020, 78, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, G.R.; Chang, P.V. Deciphering the chemical lexicon of host-gut microbiota interactions. Trends Pharmacol. Sci. 2019, 40, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.; Weersma, R.K.; Vich Vila, A. The emerging role of the small intestinal microbiota in human health and disease. Gut Microbes 2023, 15, 2201155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, S.; Ma, N.; Johnston, L.J.; Wu, C.; Ma, X. Metabolites of microbiota response to tryptophan and intestinal mucosal immunity: A therapeutic target to control intestinal inflammation. Med. Res. Rev. 2021, 41, 1061–1088. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, Z.; Xu, C.; Kan, S.; Chen, D. Disturbances of the gut microbiota and microbiota-derived metabolites in inflammatory bowel disease. Nutrients 2022, 14, 5140. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Ding, Y.; Saeidi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2019, 28, 3285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.; Wang, N.; Li, Q.; Zhang, L.; Han, N.; Yan, T.; Ren, D.; Zhang, B.; Zhao, Y.; et al. Gut bacterial indole-3-acetic acid induced immune promotion mediates preventive effects of fu brick tea polyphenols on experimental colitis. J. Agric. Food Chem. 2023, 71, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Paudel, S.; Mishra, N.; Agarwal, R. Phytochemicals as immunomodulatory molecules in cancer therapeutics. Pharmaceuticals 2023, 16, 1652. [Google Scholar] [CrossRef]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry. J. Anim. Sci. 2008, 86, E140–E148. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.R.; Davoodi, H. Herbal plants and their derivatives as growth and health promoters in animal nutrition. Vet. Res. Commun. 2011, 35, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, C.; Badgujar, P.C.; Gundev, P.; Upadhyay, A. Review of toxicological assessment of d-limonene, a food and cosmetics additive. Food Chem. Toxicol. 2018, 120, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Raphael, T.J.; Kuttan, G. Immunomodulatory activity of naturally occurring monoterpenes carvone, limonene, and perillic acid. Immunopharmacol. Immunotoxicol. 2003, 25, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Raphael, T.J.; Kuttan, G. Protective role of perillic acid against radiation-induced oxidative stress, cytokine profile, DNA damage, and intestinal toxicity in mice. J. Environ. Pathol. Toxicol. Oncol. 2010, 29, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, H.; You, Y.; Zhong, G.; Ruan, Z.; Liao, J.; Zhang, H.; Pan, J.; Tang, Z.; Hu, L. Multi-omics reveals the protective effects of curcumin against afb1-induced oxidative stress and inflammatory damage in duckling intestines. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2024, 276, 109815. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; He, J.; Ahmad, H.; Shen, M.; Zhao, Y.; Gan, Z.; Zhang, L.; Zhong, X.; Wang, C.; Wang, T. Dietary curcumin supplementation increases antioxidant capacity, upregulates nrf2 and hmox1 levels in the liver of piglet model with intrauterine growth retardation. Nutrients 2019, 11, 2978. [Google Scholar] [CrossRef] [PubMed]
- Gan, Z.; Wei, W.; Wu, J.; Zhao, Y.; Zhang, L.; Wang, T.; Zhong, X. Resveratrol and curcumin improve intestinal mucosal integrity and decrease m(6)a rna methylation in the intestine of weaning piglets. ACS Omega 2019, 4, 17438–17446. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef]
- Yazdi, M.; Nafari, A.; Azadpour, M.; Alaee, M.; Hadipour Moradi, F.; Choghakhori, R.; Hormozi, M.; Ahmadvand, H. Protective effects of cinnamic acid against hyperglycemia induced oxidative stress and inflammation in hepg2 cells. Rep. Biochem. Mol. Biol. 2023, 12, 1–12. [Google Scholar] [CrossRef]
- Lee, A.G.; Kang, S.; Im, S.; Pak, Y.K. Cinnamic acid attenuates peripheral and hypothalamic inflammation in high-fat diet-induced obese mice. Pharmaceutics 2022, 14, 1675. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, T.; Koci, M.; Wang, Y.; Fu, Y.; Ma, M.; Ma, Q.; Zhao, L. Chlorogenic acid alleviated afb1-induced hepatotoxicity by regulating mitochondrial function, activating nrf2/ho-1, and inhibiting noncanonical nf-kappab signaling pathway. Antioxidants 2023, 12, 2027. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Chen, P.; Wang, Y.; Xu, L.; Zhang, K.; Zhao, J.; Liu, H. Chlorogenic acid protects against intestinal inflammation and injury by inactivating the mtdna-cgas-sting signaling pathway in broilers under necrotic enteritis challenge. Poult. Sci. 2023, 103, 103274. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiao, G.; Cheng, P.; Zeng, J.; Liu, Y. Protective application of chinese herbal compounds and formulae in intestinal inflammation in humans and animals. Molecules 2023, 28, 6811. [Google Scholar] [CrossRef] [PubMed]
- Kendig, D.M.; Grider, J.R. Serotonin and colonic motility. Neurogastroenterol. Motil. 2015, 27, 899–905. [Google Scholar] [CrossRef]
- Gill, R.K.; Saksena, S.; Tyagi, S.; Alrefai, W.A.; Malakooti, J.; Sarwar, Z.; Turner, J.R.; Ramaswamy, K.; Dudeja, P.K. Serotonin inhibits na+/h+ exchange activity via 5-ht4 receptors and activation of pkc alpha in human intestinal epithelial cells. Gastroenterology 2005, 128, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, O. 5-hydroxytryptamine, enterotoxins, and intestinal fluid secretion. Gastroenterology 1998, 115, 1009–1012. [Google Scholar] [CrossRef] [PubMed]
- Keszthelyi, D.; Troost, F.J.; Masclee, A.A. Understanding the role of tryptophan and serotonin metabolism in gastrointestinal function. Neurogastroenterol. Motil. 2009, 21, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.H.; Wang, H.; Denou, E.; Ghia, J.E.; Rossi, L.; Fontes, M.E.; Bernier, S.P.; Shajib, M.S.; Banskota, S.; Collins, S.M.; et al. Modulation of gut microbiota composition by serotonin signaling influences intestinal immune response and susceptibility to colitis. Cell Mol. Gastroenterol. Hepatol. 2019, 7, 709–728. [Google Scholar] [CrossRef]
- Wang, B.; Sun, S.; Liu, M.; Chen, H.; Liu, N.; Wu, Z.; Wu, G.; Dai, Z. Dietary l-tryptophan regulates colonic serotonin homeostasis in mice with dextran sodium sulfate-induced colitis. J. Nutr. 2020, 150, 1966–1976. [Google Scholar] [CrossRef]
- Hao, Y.; Jiang, L.; Han, D.; Si, D.; Sun, Z.; Wu, Z.; Dai, Z. Limosilactobacillus mucosae and lactobacillus amylovorus protect against experimental colitis via upregulation of colonic 5-hydroxytryptamine receptor 4 and transforming growth factor-beta2. J. Nutr. 2023, 153, 2512–2522. [Google Scholar] [CrossRef] [PubMed]
- Fabà, L.; de Groot, N.; Ramis, G.; Cabrera-Gómez, C.G.; Doelman, J. Serotonin receptors and their association with the immune system in the gastrointestinal tract of weaning piglets. Porc. Health Manag. 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The diverse metabolic roles of peripheral serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut microbiota-produced tryptamine activates an epithelial g-protein-coupled receptor to increase colonic secretion. Cell Host Microbe 2018, 23, 775–785 e775. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Han, D.; Hao, Y.; Song, Z.; Sun, Z.; Dai, Z. Linking serotonin homeostasis to gut function: Nutrition, gut microbiota and beyond. Crit. Rev. Food Sci. Nutr. 2023, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jiang, L.; Wu, Z.; Dai, Z. L-tryptophan differentially regulated glucose and amino acid transporters in the small intestine of rat challenged with lipopolysaccharide. Animals 2022, 12, 3045. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., 3rd; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Z.H.; Zabed, H.M.; Yun, J.; Zhang, G.; Qi, X. An insight into the roles of dietary tryptophan and its metabolites in intestinal inflammation and inflammatory bowel disease. Mol. Nutr. Food Res. 2021, 65, e2000461. [Google Scholar] [CrossRef]
- Yu, K.; Li, Q.; Sun, X.; Peng, X.; Tang, Q.; Chu, H.; Zhou, L.; Wang, B.; Zhou, Z.; Deng, X.; et al. Bacterial indole-3-lactic acid affects epithelium-macrophage crosstalk to regulate intestinal homeostasis. Proc. Natl. Acad. Sci. USA 2023, 120, e2309032120. [Google Scholar] [CrossRef]
- Ben Shahar, Y.; Sukhotnik, I.; Bitterman, N.; Pollak, Y.; Bejar, J.; Chepurov, D.; Coran, A.; Bitterman, A. Effect of n-acetylserotonin on intestinal recovery following intestinal ischemia-reperfusion injury in a rat. Eur. J. Pediatr. Surg. 2016, 26, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Proietti, E.; Pauwels, R.W.M.; de Vries, A.C.; Orecchini, E.; Volpi, C.; Orabona, C.; Peppelenbosch, M.P.; Fuhler, G.M.; Mondanelli, G. Modulation of indoleamine 2,3-dioxygenase 1 during inflammatory bowel disease activity in humans and mice. Int. J. Tryptophan Res. 2023, 16, 11786469231153109. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Jeong, S.; Kim, J.; Ju, S.; Im, E.; Heo, G.; Park, S.; Yoo, J.W.; Lee, J.; Yoon, I.S.; et al. N-acetylserotonin is an oxidation-responsive activator of nrf2 ameliorating colitis in rats. J. Pineal Res. 2023, 74, e12835. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, A.M.; Pacheco, A.R.; Henrick, B.M.; Taft, D.; Xu, G.; Huda, M.N.; Mishchuk, D.; Goodson, M.L.; Slupsky, C.; Barile, D.; et al. Indole-3-lactic acid associated with bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 2020, 20, 357. [Google Scholar] [CrossRef] [PubMed]
- Knopp, F.M.; da Rocha Olivieri de Barros, R.; Drummond, B.S.; Siani, A.C.; Ferrara, M.A.; Bon, E.P.S. Production of perillic acid from orange essential oil by yarrowia lipolytica using a top-aerated bioreactor. Braz. J. Microbiol. 2023, 54, 2663–2670. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi de Alvarenga, J.F.; Lei Preti, C.; Santos Martins, L.; Noronha Hernandez, G.; Genaro, B.; Lamesa Costa, B.; Gieseler Dias, C.; Purgatto, E.; Fiamoncini, J. Identification of d-limonene metabolites by lc-hrms: An exploratory metabolic switching approach in a mouse model of diet-induced obesity. Metabolites 2022, 12, 1246. [Google Scholar] [CrossRef] [PubMed]
- Grzelczyk, J.; Budryn, G.; Peña-García, J.; Szwajgier, D.; Gałązka-Czarnecka, I.; Oracz, J.; Pérez-Sánchez, H. Evaluation of the inhibition of monoamine oxidase a by bioactive coffee compounds protecting serotonin degradation. Food Chem. 2021, 348, 129108. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.S.; Ahmed-Farid, O.A.; Nassan, M.A.; El-Tarabany, M.S. Dietary curcumin improves energy metabolism, brain monoamines, carcass traits, muscle oxidative stability and fatty acid profile in heat-stressed broiler chickens. Antioxidants 2021, 10, 1265. [Google Scholar] [CrossRef]
- Kabra, A.; Garg, R.; Brimson, J.; Živković, J.; Almawash, S.; Ayaz, M.; Nawaz, A.; Hassan, S.S.U.; Bungau, S. Mechanistic insights into the role of plant polyphenols and their nano-formulations in the management of depression. Front. Pharmacol. 2022, 13, 1046599. [Google Scholar] [CrossRef]
- Arbabi Jahan, A.; Rad, A.; Ghanbarabadi, M.; Amin, B.; Mohammad-Zadeh, M. The role of serotonin and its receptors on the anticonvulsant effect of curcumin in pentylenetetrazol-induced seizures. Life Sci. 2018, 211, 252–260. [Google Scholar] [CrossRef]
- Liu, Y.; Song, M.; Bai, H.; Wang, C.; Wang, F.; Yuan, Q. Curcumin improves the egg quality, antioxidant activity, and intestinal microbiota of quails during the late laying period. Poult. Sci. 2024, 103, 103233. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.I.; Li, D.; Wang, B.; Zahoruiko, Y.; Gerasymchuk, M.; Hudson, D.; Kovalchuk, O.; Kovalchuk, I. Anti-inflammatory effects of serotonin receptor and transient receptor potential channel ligands in human small intestinal epithelial cells. Curr. Issues Mol. Biol. 2023, 45, 6743–6774. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wu, S.; Li, J.; Wang, R.; Xie, X.; Yu, X.; Pan, J.; Xu, Y.; Zheng, L. The effect of curcumin on the brain-gut axis in rat model of irritable bowel syndrome: Involvement of 5-ht-dependent signaling. Metab. Brain Dis. 2015, 30, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Renner, U.; Zeug, A.; Woehler, A.; Niebert, M.; Dityatev, A.; Dityateva, G.; Gorinski, N.; Guseva, D.; Abdel-Galil, D.; Frohlich, M.; et al. Heterodimerization of serotonin receptors 5-ht1a and 5-ht7 differentially regulates receptor signalling and trafficking. J. Cell Sci. 2012, 125, 2486–2499. [Google Scholar] [CrossRef] [PubMed]
- Rodnyy, A.Y.; Kondaurova, E.M.; Bazovkina, D.V.; Kulikova, E.A.; Ilchibaeva, T.V.; Kovetskaya, A.I.; Baraboshkina, I.A.; Bazhenova, E.Y.; Popova, N.K.; Naumenko, V.S. Serotonin 5-ht(7) receptor overexpression in the raphe nuclei area produces antidepressive effect and affects brain serotonin system in male mice. J. Neurosci. Res. 2022, 100, 1506–1523. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yin, Y.; Li, T.; Huang, R.; Xie, M.; Wu, Z.; Wu, G. Use of the ussing chamber technique to study nutrient transport by epithelial tissues. Front. Biosci. 2013, 18, 1266–1274. [Google Scholar] [CrossRef]
- Clarke, L.L. A guide to ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Wang, Y.Z.; Frieling, T.; Wood, J.D. Neural 5-hydroxytryptamine receptors regulate chloride secretion in guinea pig distal colon. Am. J. Physiol. 1991, 261, G833–G840. [Google Scholar] [CrossRef]
- Sikander, A.; Rana, S.V.; Prasad, K.K. Role of serotonin in gastrointestinal motility and irritable bowel syndrome. Clin. Chim. Acta 2009, 403, 47–55. [Google Scholar] [CrossRef]
- Wang, Y.; Sims, C.E.; Allbritton, N.L. Enterochromaffin cell-enriched monolayer platform for assaying serotonin release from human primary intestinal cells. Anal. Chem. 2020, 92, 12330–12337. [Google Scholar] [CrossRef]
- Brighton, C.A.; Rievaj, J.; Kuhre, R.E.; Glass, L.L.; Schoonjans, K.; Holst, J.J.; Gribble, F.M.; Reimann, F. Bile acids trigger glp-1 release predominantly by accessing basolaterally located g protein-coupled bile acid receptors. Endocrinology 2015, 156, 3961–3970. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Feng, D.; Zhang, Y.; Dahanayaka, S.; Li, X.; Yao, K.; Wang, J.; Wu, Z.; Dai, Z.; Wu, G. Regulation of leucine catabolism by metabolic fuels in mammary epithelial cells. Amino Acids 2012, 43, 2179–2189. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).