Abstract
Water deficit is the major stress factor magnified by climate change that causes the most reductions in plant productivity. Knowledge of photosystem II (PSII) response mechanisms underlying crop vulnerability to drought is critical to better understanding the consequences of climate change on crop plants. Salicylic acid (SA) application under drought stress may stimulate PSII function, although the exact mechanism remains essentially unclear. To reveal the PSII response mechanism of celery plants sprayed with water (WA) or SA, we employed chlorophyll fluorescence imaging analysis at 48 h, 96 h, and 192 h after watering. The results showed that up to 96 h after watering, the stroma lamellae of SA-sprayed leaves appeared dilated, and the efficiency of PSII declined, compared to WA-sprayed plants, which displayed a better PSII function. However, 192 h after watering, the stroma lamellae of SA-sprayed leaves was restored, while SA boosted chlorophyll synthesis, and by ameliorating the osmotic potential of celery plants, it resulted in higher relative leaf water content compared to WA-sprayed plants. SA, by acting as an antioxidant under drought stress, suppressed phototoxicity, thereby offering PSII photoprotection, together with enhanced effective quantum yield of PSII photochemistry (ΦPSII) and decreased quantity of singlet oxygen (1O2) generation compared to WA-sprayed plants. The PSII photoprotection mechanism induced by SA under drought stress was triggered by non-photochemical quenching (NPQ), which is a strategy to protect the chloroplast from photo-oxidative damage by dissipating the excess light energy as heat. This photoprotective mechanism, triggered by NPQ under drought stress, was adequate in keeping, especially in high-light conditions, an equal fraction of open PSII reaction centers (qp) as of non-stress conditions. Thus, under water deficit stress, SA activates a regulatory network of stress and light energy partitioning signaling that can mitigate, to an extent, the water deficit stress on PSII functioning.
1. Introduction
Anthropogenic activities intensify the negative consequences of climate change that compromise sufficient food production [1]. It is estimated that global food production may be reduced by 11–25% at the end of this century [2]. Water deficit is the major stress factor magnified by climate change that causes the most reductions in productivity when compared to other stress factors [3,4,5,6,7,8]. The increasing demand for sustainable agricultural solutions has brought biostimulants to the forefront as valuable tools for regenerative farming [9,10]. Biostimulants, with their varied biological activities, play a significant role in enhancing crop growth, improving nutrient efficiency, sustaining resilience to environmental stressors, and restoring soil health [10].
Salicylic acid (SA) is considered an important phytohormone, being essential for controlling a broad array of biochemical and physiological processes that are involved in many stages of growth and development, including seed germination, flowering, regulation of ripening, and plant defense responses to pathogens and abiotic stimuli [11,12,13,14,15,16,17,18]. It has been suggested to have evident potential for mitigating numerous abiotic stresses of major agricultural crops, for improving their stress resistance, acting as an antioxidant, osmo-regulator, and biostimulant [17,19,20,21].
SA biosynthesis in plants has been described to start (i) in the chloroplast, where chorismate is converted to isochorismate via the isochorismate pathway and then it is transported to the cytosol, or (ii) with the amino acid L-phenylalanine in the cytosol, via the phenylalanine ammonia-lyase pathway [22]. However, in Arabidopsis, it has been shown that SA biosynthesis is not from the phenylalanine ammonia-lyase pathway [23] but it remains unclear whether this pathway is involved in other plant species [20].
An increase in SA production and signaling during defense responses is associated with a simultaneous reduction in auxin biosynthesis and signaling, thereby coordinating defense and growth [20,24]. SA application can alleviate the negative effects of water deficit and salinity by improving osmotic potential, reducing membrane damage, regulating stomatal conductance and transpiration rate, restoring biochemical parameters and photosynthetic rates, and enhancing shoot and root biomass as well as nutrient uptake [25,26]. Nevertheless, the effects of exogenously applied SA on plant physiological processes under optimal growth conditions are contentious, with some studies reporting positive effects on plant growth while others highlight negative influences on various physiological processes [27]. For instance, spraying SA on corn and soybean increased photosynthetic rates, possibly due to enhanced enzyme activity related to CO2 uptake rather than increased stomatal opening [27,28]. Conversely, applying 0.5 mM of SA to maize plants decreased net photosynthesis and stomatal conductance under both low and high light conditions [29]. Yet, the same concentration provided protection against low-temperature damage [29] or mitigated the damaging effects of paraquat [30]. The exogenous application of SA mitigated the effects of mild water deficit in radish plants [31].
Water deficit stress results in stomatal closure, limiting CO2 assimilation and, thus, reducing carbon fixation, which results in excess light energy that cannot be used for photochemistry [6,32,33,34,35]. This excess light energy is prospective for injury if it is transferred to O2, producing photo-oxidative damage and reducing photosynthetic efficiency [36,37,38,39,40]. Thus, the efficient management of the excess light energy that cannot be assigned to photochemistry under water deficit stress is fundamental [35,36,37]. Plants can dissipate this excess excitation energy as heat, via the mechanism of non-photochemical quenching (NPQ), to protect the chloroplasts and especially photosystem II (PSII) from photo-oxidative damage [35,36,40]. Knowledge of the PSII response mechanisms underlying crop vulnerability to water deficit stress is critical to better understanding the consequences of climate change on crop plants [4,6,7,35]. Investigating PSII-adaptive strategies to water deficit should, thus, assist in fulfilling this knowledge gap. Photosynthetic light energy use efficiency is generally mentioned to govern crop yields [41,42,43,44]. One key approach to improve the light energy utilization efficiency is to boost the dissipation of excess excitation energy by the NPQ mechanism [45,46,47,48,49].
The exogenous application of SA can modulate the electron transport rate (ETR) in photosystem II (PSII), by the NPQ mechanism that dissipates the excess light energy as heat [27,50,51,52]. However, the mechanism of the SA-mediated response of PSII photochemistry under non-stress or drought stress conditions is diverse [51,52]. PSII photochemistry was reported to be enhanced by SA under non-stress conditions only under high light but under drought stress at both low light and high light conditions [52]. In addition, the SA mode of action has been described to differ between plant species [51,52,53]. These different effects of SA on distinct plant species can be attributed to variations in its biosynthesis and signaling pathways in different plants [22]. The plant species variety of the SA defense signaling is also apparent in SA accumulation, interplay with other hormones, and redox regulation [54].
Celery (Apium graveolens L.) is an important biennial vegetable crop, widely grown worldwide and dating back to the 7th century B.C. [55,56]. It originated in the Middle East and the Mediterranean area and now has a large geographical distribution and a long cultivation history [55,56]. It is used in medicine as an anti-inflammatory and antibacterial agent and in food and cosmetic industries [57,58]. Celery is considered a drought-tolerant species due to its antioxidant activities [56]. The exogenous application of proline has been shown to increase the salt tolerance of celery [59] while foliar fertilizers improve its yield and quality [60].
In our research work, we tested the hypothesis that the foliar application of SA would improve the light energy utilization efficiency in celery plants by boosting the dissipation of excess excitation energy via the NPQ mechanism, thus enhancing PSII function and improving drought tolerance. We hypothesize that SA-sprayed leaves under water deficit conditions would have an improved osmotic potential and would maintain a higher leaf water content that may enable an enhanced PSII function compared to WA-sprayed leaves with reduced leaf water content.
2. Results
2.1. Soil and Leaf Water Content under Treatments
Celery plants were watered, then foliar sprayed with water or SA once, while measurements were performed 48 h, 96 h, and 192 h after the spray. Forty-eight hours after watering, the soil water content of WA-sprayed plants decreased to 70 ± 3% of the full soil water capacity, while in SA-sprayed plants, the soil water content decreased to 82 ± 4% (Table 1). At the further elapsed time from watering, 96 h and 192 h, the soil water content of WA-sprayed plants decreased further to 27 ± 2% and 5 ± 1% of the full soil water capacity, respectively, while in the SA-sprayed plants, this decreased to 38 ± 3% and 9 ± 2%, respectively (Table 1).

Table 1.
Soil and leaf water content and chlorophyll content of celery plants under treatments.
The same trend was noticed in the leaf relative water content: at 48 h, 96 h, and 192 h after watering, in WA-sprayed plants, this content decreased to 81 ± 0.4%, 79 ± 0.2%, and 73 ± 0.1%, respectively; meanwhile, in the SA-sprayed plants, this decreased to 85 ± 0.3%, 83 ± 0.3%, and 80 ± 0.2%, respectively (Table 1).
2.2. Chlorophyll Content in Water-Sprayed and Salicylic Acid-Sprayed Plants
Chlorophyll content in WA-sprayed leaves 48 h after watering was higher compared to SA-sprayed leaves but at 96 h after watering, chlorophyll content was higher in SA-sprayed leaves (Table 1). At the water deficit conditions, 192 h after watering, an amplification of chlorophyll content was observed in SA-sprayed plants while in WA-sprayed plants, chlorophyll content decreased to the lowest level (Table 1).
2.3. Light Energy Partitioning in Water-Sprayed and Salicylic Acid-Sprayed Plants
The yield for photochemistry (ΦPSII) and the yield losses, regulated (ΦNPQ), and non-regulated (ΦNO), have a sum equal to 1 [61]. Forty-eight hours after watering, ΦPSII in WA-sprayed leaves was significantly higher than in SA-sprayed leaves at both low light (LL) (Figure 1a) and high light (HL) (Figure 1b). At 96 h after watering, ΦPSII decreased in SA-sprayed leaves under both LL (Figure 1a) and HL (Figure 1b) but in WA-sprayed leaves, ΦPSII decreased only under LL (Figure 1a). At the prolonged time from watering (192 h, water deficit conditions), ΦPSII in WA-sprayed leaves decreased to the lowest level under both LL (Figure 1a) and HL (Figure 1b); meanwhile, in SA-sprayed leaves at 192 h after watering, ΦPSII at LL remain the same to that of 96 h after watering (Figure 1a) and at HL, to that of 48 h after watering (Figure 1b).
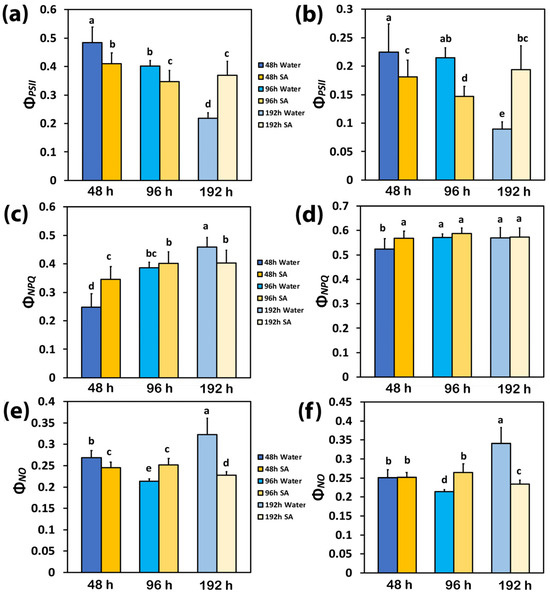
Figure 1.
Light energy partitioning at PSII. The effective quantum yield of PSII photochemistry (ΦPSII) at low light (LL) (a) and at high light (HL) (b); the quantum yield of regulated non-photochemical energy loss in PSII (ΦNPQ) at LL (c) and at HL (d); and the quantum yield of non-regulated energy loss in PSII (ΦNO) at LL (e) and at HL (f) of water-sprayed or salicylic acid-sprayed (SA), celery plants at 48 h, 96 h, and 192 h after watering. Standard deviations (SD) are shown as error bars (n = 9, 3 independent experiments with 3 plants for each treatment and time point measurement). Significant differences are expressed by different alphabet letters.
At 48 h after watering, the decreased yield for photochemistry (ΦPSII) in SA-sprayed plants, compared to WA-sprayed ones (Figure 1a), was overcompensated at LL by the increased yield of regulated non-photochemical energy loss in PSII (ΦNPQ) (Figure 1c), which resulted in a lower yield of non-regulated energy loss in PSII (ΦNO) (Figure 1e). At 48 h after watering at HL, the increased ΦNPQ in SA-sprayed leaves, compared to WA-sprayed ones (Figure 1d), resulted in the same level of ΦNO with the WA-sprayed ones (Figure 1f) despite its lower ΦPSII (Figure 1b).
At 96 h after watering, the decreased ΦPSII in WA-sprayed plants (Figure 1a), compared to WA-sprayed plants at 48 h after watering, was overcompensated at LL by the increased yield of ΦNPQ (Figure 1c), which resulted in reduced ΦNO compared to WA-sprayed plants at 48 h after watering (Figure 1e). At 96 h after watering at HL, the increased ΦNPQ in WA-sprayed leaves (Figure 1d) developed reduced ΦNO compared to WA-sprayed plants at 48 h after watering (Figure 1f).
At 192 h after watering, the lowest level of ΦPSII in WA-sprayed leaves under both LL (Figure 1a) and HL (Figure 1b) could not be compensated by the increased ΦNPQ (Figure 1c,d); thus, the highest levels of ΦNO were developed (Figure 1e,f). In contrast, in SA-sprayed leaves, the increased ΦNPQ at both LL and HL (Figure 1c,d) could decrease ΦNO to lower levels than those observed in SA-sprayed leaves at 48 h after watering (Figure 1e,f).
2.4. The Photoprotective Heat Dissipation and the Electron Transport Rate in Water-Sprayed and Salicylic Acid-Sprayed Plants
The photoprotective heat dissipation, via non-photochemical quenching (NPQ), increased with elapsed time from watering in both WA-sprayed and SA-sprayed leaves at both LL and HL (Figure 2a,b). The only exceptions were the WA-sprayed leaves at 192 h under both LL and HL, at which NPQ decreased, and the SA-sprayed leaves at 96 h under both LL and HL, at which NPQ remained at the level of 48 h after watering (Figure 2a,b).
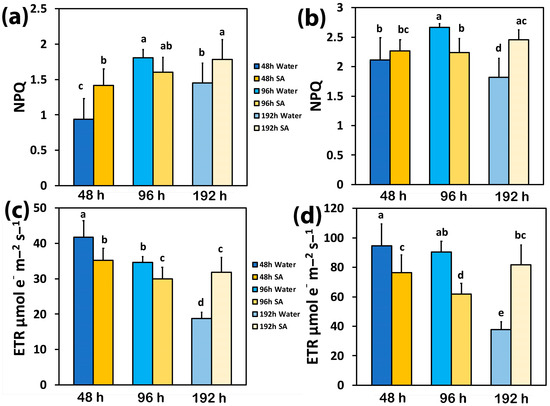
Figure 2.
The non-photochemical quenching (NPQ) at low light (LL) (a) and at high light (HL) (b) and the electron transport rate (ETR) at LL (c) and at HL (d) of water-sprayed or salicylic acid-sprayed (SA) celery plants at 48 h, 96 h, and 192 h after watering. Standard deviations (SD) are shown as error bars (n = 9, 3 independent experiments with 3 plants for each treatment and time point measurement). Significant differences are expressed by different alphabet letters.
Forty-eight hours after watering, the electron transport rate (ETR) in WA-sprayed leaves was significantly higher than in SA-sprayed leaves at both LL (Figure 2c) and HL (Figure 2d). At 96 h after watering, ETR decreased in SA-sprayed leaves under both LL and HL (Figure 2c,d) but in WA-sprayed leaves, ETR decreased only under LL (Figure 2c). Under water deficit conditions (192 h after watering), ETR decreased in WA-sprayed leaves to the lowest level under both LL (Figure 2c) and HL (Figure 2d); meanwhile, in SA-sprayed leaves at 192 h after watering, ETR remained at the same level as at 96 h after watering at LL (Figure 2c) and at that of 48 h after watering at HL (Figure 2d).
2.5. The Fraction of Open PSII Reaction Centers and Their Efficiency in Water-Sprayed and Salicylic Acid-Sprayed Plants
The redox state of quinone A (QA), or the photochemical quenching (qp), also representing the fraction of open PSII reaction centers (RCs), was significantly higher in WA-sprayed leaves compared to SA-sprayed leaves at 48 h and 96 h after watering at both LL (Figure 3a) and HL (Figure 3b). However, at 192 h after watering, a higher portion of open PSII RCs was observed in SA-sprayed leaves compared to WA-sprayed ones at both LL (Figure 3a) and HL (Figure 3b). Under both LL and HL, WA-sprayed leaves possessed the lowest fraction of open PSII RCs while SA-sprayed leaves could retain the same fraction of open PSII RCs to that observed at 48 h after watering (Figure 3a,b).
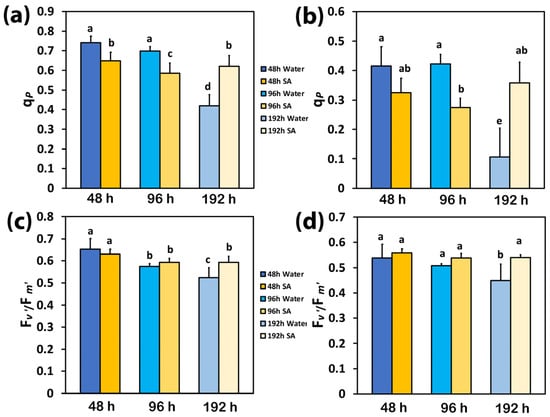
Figure 3.
The photochemical quenching (qp), representing the fraction of open PSII reaction centers (RCs) at low light (LL) (a) and at high light (HL) (b), and the efficiency of the open PSII RCs (Fv′/Fm′) at LL (c) and at HL (d) of water-sprayed or salicylic acid-sprayed (SA) celery plants at 48 h, 96 h, and 192 h after watering. Standard deviations (SD) are shown as error bars (n = 9, 3 independent experiments with 3 plants for each treatment and time point measurement). Significant differences are expressed by different alphabet letters.
The efficiency of the open PSII RCs (Fv′/Fm′) at both LL and HL did not differ between WA-sprayed and SA-sprayed leaves at 48 h and 96 h after watering (Figure 3c,d). However, at 192 h after watering, a higher efficiency of the open PSII RCs was observed in SA-sprayed leaves at both LL (Figure 3c) and HL (Figure 3d). At 192 h after watering at HL, the efficiency of the open PSII RCs in SA-sprayed leaves was similar to that at 48 h after watering (Figure 3d) while at LL, it was similar to that at 96 h after watering (Figure 3c).
2.6. Excess Excitation Energy and PSII Excitation Pressure in Water-Sprayed and Salicylic Acid-Sprayed Plants
At 48 h and 96 h after watering, the excitation pressure at PSII (1−qL) in SA-sprayed leaves was higher at both LL (Figure 4a) and HL (Figure 4b) compared to WA-sprayed leaves; but at 192 h after watering, the excitation pressure (1−qL) at both LL (Figure 4a) and HL (Figure 4b) was higher in WA-sprayed leaves.
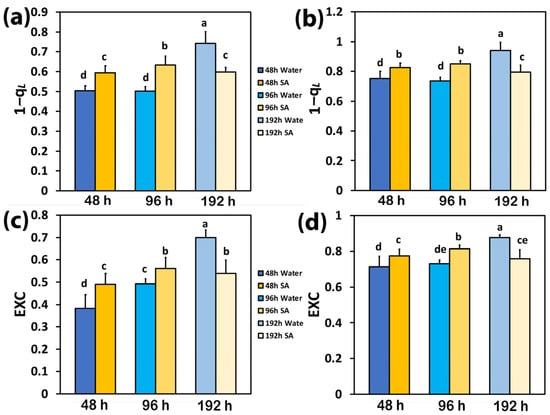
Figure 4.
The excitation pressure at PSII (1−qL) at low light (LL) (a) and at high light (HL) (b) and the excess excitation energy at PSII (EXC) at LL (c) and at HL (d) of water-sprayed or salicylic acid-sprayed (SA) celery plants at 48 h, 96 h, and 192 h after watering. Standard deviations (SD) are shown as error bars (n = 9, 3 independent experiments with 3 plants for each treatment and time point measurement). Significant differences are expressed by different alphabet letters.
The same trend with excitation pressure (1−qL) was observed in the excess excitation energy at PSII (EXC), which at both LL (Figure 4c) and HL (Figure 4d) was higher in SA-sprayed leaves, at both 48 h and 96 h after watering, compared to WA-sprayed leaves; but at 192 h after watering, the excess excitation energy (EXC) at both LL (Figure 4c) and HL (Figure 4d) was higher in WA-sprayed leaves.
2.7. The Spatiotemporal Heterogeneity of PSII Function in Water-Sprayed and Salicylic Acid-Sprayed Plants
The whole leaf area color-coded pictures of ΦPSII, ΦNPQ, ΦNO, and qp that were obtained with chlorophyll fluorescence imaging, at 48 h after watering (Figure 5) and at 192 h after watering (Figure 6), are presented. Forty-eight hours after watering, the yield for photochemistry (ΦPSII) decreased more in SA-sprayed leaves compared to WA-sprayed ones (Figure 5). However, a higher leaf heterogeneity of the parameter ΦPSII was observed in WA-sprayed leaves compared to SA-sprayed leaves (Figure 5). This higher leaf heterogeneity of the parameter ΦPSII in WA-sprayed leaves, compared to SA-sprayed ones, was noticed also for the parameter ΦNPQ (Figure 5). The higher increase in ΦNPQ in SA-sprayed leaves could overcompensate for the lower ΦPSII values compared to WA-sprayed leaves, resulting in decreased ΦNO values in SA-sprayed leaves (Figure 5). The higher ΦPSII values in WA-sprayed leaves also implied a higher fraction of open PSII RCs compared to SA-sprayed leaves (Figure 5). Thus, at 48 h after watering, the WA-sprayed leaves presented a better PSII functionality than SA-sprayed leaves.
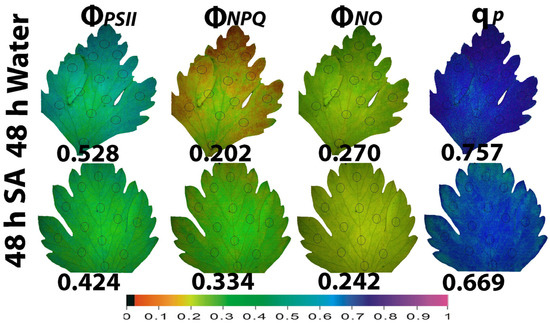
Figure 5.
Whole leaf area color-coded pictures of ΦPSII, ΦNPQ, ΦNO, and qp, obtained at 200 μmol photons m−2 s−1, in water-sprayed and salicylic acid-sprayed (SA) celery plants at 48 h after watering. The average whole leaf value of each chlorophyll parameter is provided. At the bottom, the color code indicates the corresponding parameter value as color with a scale from 0 to 1.
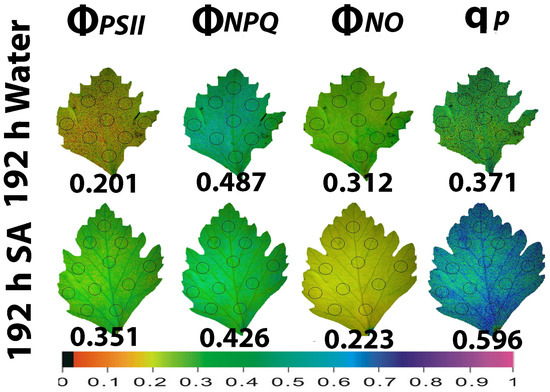
Figure 6.
Whole leaf area color-coded pictures of ΦPSII, ΦNPQ, ΦNO, and qp, obtained at 200 μmol photons m−2 s−1, in water-sprayed and salicylic acid-sprayed (SA) celery plants at 192 h after watering. The average whole leaf value of each chlorophyll parameter is provided. At the bottom, the color code indicates the corresponding parameter value as color with a scale from 0 to 1.
At the prolonged time from watering (192 h, water deficit conditions), ΦPSII in WA-sprayed leaves was significantly lower than in SA-sprayed leaves (Figure 6). This decreased ΦPSII in WA-sprayed leaves could not be balanced by the increased ΦNPQ having as a result an increased ΦNO in WA-sprayed leaves, compared to SA-sprayed leaves (Figure 6). At 192 h after watering, SA-sprayed leaves also possessed a higher fraction of open PSII RCs compared to WA-sprayed leaves (Figure 6). Thus, at 192 h after watering, a better PSII functionality was observed in SA-sprayed leaves compared to WA-sprayed leaves (Figure 6). SA-sprayed leaves could maintain at 48 h (Figure 5) and at 192 h (Figure 6) after watering, lower ΦNO values compared to WA-sprayed leaves.
2.8. Chloroplast Ultrastructure in Water-Sprayed and Salicylic Acid-Sprayed Plants
Most of the typical ultrastructural features of chloroplasts, such as plastid envelope integrity, abundance of grana, starch grains, and plastoglobuli, were noticed in celery leaves, resembling those that are constantly observed by transmission electron microscopy (TEM) in leaf cells (Figure 7). However, in the chloroplasts of SA-sprayed celery leaves, at 48 h and 96 h after watering, the stroma lamellae appeared dilated (Figure 7b,d) in comparison with those observed in WA-sprayed leaves (Figure 7a,c). Interestingly, the above dilation was not observed 192 h after watering (water deficit conditions) in chloroplasts of SA-sprayed leaves, the stroma lamellae of which (Figure 7f) appeared normal, like those of WA-sprayed leaves at 48 h and 96 h after watering.
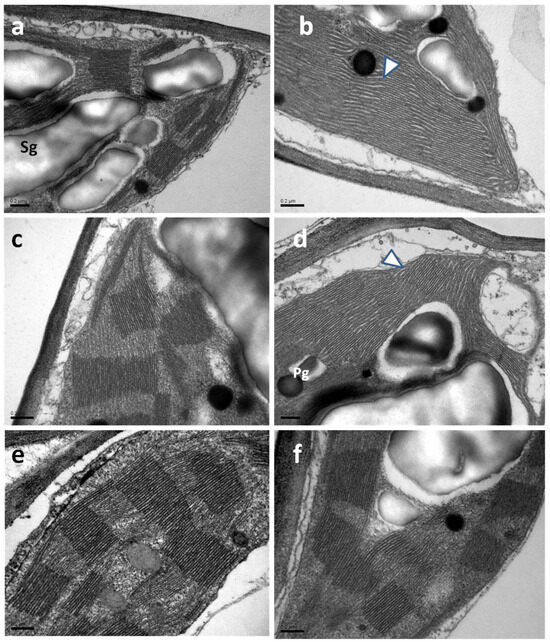
Figure 7.
TEM micrographs depicting chloroplast ultrastructure in leaves of WA-sprayed (a,c,e) and SA-sprayed (b,d,f) plants at 48 h (a,b), 96 h (c,d), and 192 h (e,f) after watering. Note the dilated appearance of stroma lamellae in chloroplasts of SA-sprayed plants 48 h and 96 h after watering [arrowheads in (b,d)]. Pg: plastoglobuli, Sg: starch grain, scale bars: 0.2 μm.
3. Discussion
Molecular mechanisms induced by biostimulants that can enhance osmolyte accumulation, e.g., proline, can counterbalance water loss and are efficient at maintaining high leaf water potential while soil water content is low [62,63,64]. Biostimulant-driven solutions can enhance crop drought tolerance and, thus, resolve a threatening agricultural challenge [64]. Many studies have demonstrated that the exogenous application of SA increases plant tolerance to drought stress by acting as an osmo-regulator [52,65]. This phenomenon is commonly referred to as osmotic adjustment, an essential physiological mechanism for drought tolerance [62,65]. In our experiment, at an increased time from watering, the soil water content in both WA-sprayed and SA-sprayed plants decreased, but SA-sprayed plants retained a higher percentage of soil water content under all time point measurements (Table 1). This was possible due to the reduced transpiration of SA-sprayed plants. In addition, SA-sprayed plants retaining a higher percentage of soil water content, they also maintained a higher leaf water content (Table 1).
At 48 h after watering, the chlorophyll content was higher in WA-sprayed leaves compared to SA-sprayed (Table 1). However, at 96 h and 192 h after watering, chlorophyll content enhanced in SA-sprayed leaves in a logarithmic-dependent manner to the leaf water loss (Table 1). A decrease in chlorophyll content under non-stress conditions and an increase under water deficit stress via the application of SA have been mentioned before [51,52]. Under deficit stress conditions (192 h after watering), chlorophyll content in WA-sprayed leaves diminished (Table 1). This may be ascribed to the oxidation of chlorophyll molecules in WA-sprayed leaves [66,67], which was reversed via the application of SA, which acted as an antioxidant in SA-sprayed leaves [52,68]. SA has been reported to act as a signal molecule activating chlorophyll catabolic genes [69]. An increasing or decreasing chlorophyll content through the application of SA linked to the plant species or even to the genotype [51,52,70,71,72] and/or to the concentration of SA used has been stated [73,74]. It has been frequently concluded that the mode of SA action varies considerably depending on the plant species, the genotype, the concentration used, the environmental conditions, and the duration of exposure [27,51,52,75,76].
Up to 96 h after watering, WA-sprayed celery leaves possessed a higher quantum yield of PSII photochemistry (ΦPSII) under both LL and HL compared to SA-sprayed leaves (Figure 1a,b). The dilation of stroma lamellae in the chloroplasts of SA-sprayed leaves (Figure 7b,d), but not in those of water-sprayed leaves (Figure 7a,c), could be an observable manifestation of the above difference. Thus, under non-stress conditions up to 96 h after watering, WA-sprayed celery leaves possessed higher ΦPSII values (Figure 1a,b) and ETR (Figure 2c,d) at both LL and HL compared to SA-sprayed celery leaves. These higher quantum yields of PSII photochemistry (ΦPSII) and electron transport rates (ETR) of WA-sprayed celery leaves were due to the higher fraction of open PSII RCs (Figure 3a,b) since the efficiency of the open PSII RCs (Figure 3c,d) did not differ between WA-sprayed and SA-sprayed leaves. In addition, WA-sprayed celery leaves, up to 96 h after watering, retained lower excess excitation energy at PSII (EXC) (Figure 4c,d) and lower excitation pressure (1−qL) (Figure 4a,b) compared to SA-sprayed leaves at both LL and HL. It seems that the delay in the enhancing response of PSII photochemistry to the SA spray was longer than 96 h.
WA-sprayed celery leaves 192 h after watering displayed the lowest quantum yield of PSII photochemistry (ΦPSII) under both LL and HL (Figure 1a,b) and the highest level of quantum yield of non-regulated energy loss in PSII (ΦNO), also under both LL and HL (Figure 1c,d). An increased ΦNO denotes an increased triplet chlorophyll state (3Chl*) population that results in the creation of singlet oxygen (1O2) [77,78,79]. Therefore, the possibility of 1O2 development can be calculated by ΦNO [79,80,81]. A decreased ΦNO, 192 h after watering in SA-sprayed celery leaves, compared to WA-sprayed ones (Figure 1c,d), reveals a lower 1O2 production [35,80,81,82] and suggests that SA acting as an antioxidant offers a better photoprotection. The lower fraction of open PSII reaction centers (qp) in WA-sprayed celery leaves 192 h after watering (Figure 3a,b), corresponding to a more reduced state of the plastoquinone (PQ) pool, was connected with a significant increase in 1O2 production (Figure 1c,d).
It is feasible that the increased generation of 1O2 production in WA-sprayed celery leaves 192 h after watering contributed to the increased photoinhibition of PSII [83]. Photoinhibition is described as an imbalance between PSII photodamage and PSII repair [84,85]. The photoprotective mechanism to prevent the photo-oxidative damage of excess excitation energy is the non-photochemical quenching (NPQ) [36,37,38,40,86]. The absorbed light energy that is not used for photochemistry, nor dissipated as heat for photoprotection via the NPQ mechanism, is the cause of PSII damage [87,88,89]. In WA-sprayed celery leaves 192 h after watering, the energy dissipation via NPQ (Figure 2a,b) was insufficient to prevent the over-excitation of the photosynthetic apparatus, resulting in increased 1O2 production (Figure 1c,d), which was supplemented by excess excitation energy at PSII (EXC) (Figure 4c,d) and increased excitation pressure (1−qL) (Figure 4a,b).
In contrast to WA-sprayed celery leaves—in which 192 h after watering, the 1O2 production increased—in SA-sprayed leaves, SA, acting as an antioxidant, decreased 1O2 production and enhanced the effective quantum yield of PSII (ΦPSII). The constructive role of SA under several environmental stress conditions is related to its capacity to reduce oxidative damage [68,90,91], serve as an antioxidant [51,52,68], and perform a controlling role in photosynthetic light reactions [92]. It is concluded that under deficit stress conditions, the high increase in NPQ in SA-sprayed leaves decreased 1O2 production, thereby inducing acclimation responses to water deficit stress. Singlet oxygen (1O2) is a reactive oxygen species (ROS) that can be developed in plants as a byproduct of photosynthesis [36,38,40,93]. ROS, such as superoxide anion radical (O2•−), hydrogen peroxide (H2O2), and singlet-excited oxygen (1O2), are constantly produced but are scavenged by the antioxidant enzymatic and non-enzymatic cellular mechanisms [40,94,95,96,97,98,99]. The function of chloroplast antioxidants is not to entirely remove ROS, but to accomplish a proper equilibrium between creation and removal so as to counterbalance the process of photosynthesis and document an efficient spread of signal wave [99,100,101,102]. ROS are fundamental signaling molecules that allow cells to respond speedily to diverse kinds of alterations to their homeostasis, contributing to the creation of defense mechanisms and plant resilience [103,104].
The photoprotective mechanism of NPQ is considered to be adequate, under stressful conditions, if it can retain an equal fraction of open PSII reaction centers (qp) as in non-stress conditions [4,99,105,106]. If not, an inconsistency between the absorbed light energy and the requirement occurs, indicating excess excitation energy [36,38,105]. The redox state of the PQ pool is recognized to be essential for retrograde signaling [107,108,109]. The fraction of open PSII reaction centers, or the redox state of the plastoquinone pool (qp), also involves a mechanism of plant acclimation to abiotic stresses by controlling the photosynthetic gene expression [98,110,111,112] and is of remarkable importance for antioxidant defense and signaling [113].
The induction of NPQ for preventing harmful ROS production, under environmentally stressful conditions [19,38,40,114], is connected with dynamic changes in protein–membrane association for regulating photosynthetic ETR [115]. It can be postulated that under water deficit conditions (192 h after watering), the high increase in NPQ in SA-sprayed leaves (Figure 2a,b), contributed to the restoration of the protein–membrane changes in the thylakoid structure (Figure 7f), thereby regulating the photosynthetic ETR of SA-sprayed leaves. SA protected PSII in Arabidopsis thaliana under HL by dissipating excess excitation energy and alleviating photoinhibition through the enhanced repair of the D1 protein [116]. Also, SA enhanced PSII efficiency in basil plants under non-stress conditions and improved photoprotection under mild drought stress by dissipating excess excitation energy [117].
To achieve a deeper understanding of SA-mediated defense networks and plant tolerance to environmental stresses, it is crucial to utilize a combination of plant physiology, molecular biology, computational biology, genomic, biochemistry, and bioinformatic approaches [118]. Future research based on these approaches can uncover the complex mechanisms behind SA-mediated defense pathways and their interactions with other signaling molecules for boosting plant stress resistance [118].
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Celery (Apium graveolens L.) plants grown in standard potting soil medium were purchased from the “Garden Center Vaseiliadis” and transferred to a growth room with a light intensity of 200 ± 10 µmol photons m−2 s−1 and a 14 h photoperiod with a 21 ± 1/19 ± 1 °C day/night temperature and a relative humidity of 55 ± 5/65 ± 5% day/night.
4.2. Salicylic Acid Treatments
All celery plants were irrigated at full soil water capacity and then sprayed with 15 mL of either distilled water (WA, control) or with 1 mM of salicylic acid (SA) [51] one time. Measurements were performed 48 h, 96 h, and 192 h after the foliar spray of the well-watered plants. The experiment was executed with three independent repetitions, each with three plants per treatment and time point measurement.
4.3. Soil Volumetric Water Content
Soil volumetric water content was assessed using the soil moisture sensor (5TE; Decagon Devices, Pullman, WA, USA) combined with the readout device (ProCheck; Decagon Devices). The results of the volumetric soil water content measured in m3 m−3 were expressed as the percentage of the well-watered celery plants.
4.4. Leaf Water Content
The leaf water content of celery plants was evaluated using the electronic moisture balance (MOC120H, Shimadzu, Tokyo, Japan) as described previously [119].
4.5. Chlorophyll Content
The chlorophyll content of celery plants was expressed in relative units after being measured photometrically with a portable chlorophyll content meter (Model Cl-01, Hansatech Instruments Ltd., Norfolk, UK) [120].
4.6. Chlorophyll Fluorescence Imaging Analysis
Chlorophyll fluorescence imaging analysis was performed using the modulated Imaging-PAM Fluorometer M-Series (Heinz Walz GmbH, Effeltrich, Germany) as described in detail previously [121]. Celery plants were dark-adapted for 30 min before measurements that were conducted 48 h, 96 h, and 192 h after watering using the actinic light (AL) of 200 μmol photons m−2 s−1 (low light, LL) or 900 μmol photons m−2 s−1 (high light, HL). The chlorophyll fluorescence parameters that were assessed, using the Win software (Heinz Walz GmbH, Effeltrich, Germany), are described in Supplementary Table S1. Color-coded images of WA-sprayed and SA-sprayed celery plants that were recorded 48 h and 192 h after watering are presented.
4.7. Transmission Electron Microscopy
Leaves of plants treated as described in Section 4.2 were free-hand-cut in pieces measuring ~2 × 2 mm2, which underwent immediate fixation in 3% glutarhaldehyde in 50 mM sodium cacodylate buffer (pH 7) for 4 h at room temperature. After 3 rinses, 15 min each, in the same buffer, the samples were post-fixed overnight in 1% osmium tetroxide at 4 °C. After rinsing as above, the samples were gradually dehydrated in an acetone series, treated 2 × 20 min with propylenoxide at 4 °C, infiltrated, and finally embedded in Spurr’s resin. Ultrathin sections (~70 nm) were cut with a diamond knife and collected on copper grids. Four grids per treatment were examined. After double staining with uranyl acetate and lead citrate, the sections were examined at 80 kV with a JEOL JEM 1011 (JEOL, Tokyo, Japan) transmission electron microscope. Electron micrographs were acquired with a GATAN 500 digital camera (Gatan, Pleasanton, CA, USA).
4.8. Statistical Analysis
All statistical analyses were performed in R software, version 4.3.1 (R Core Team, 2023). The data were tested for normality and homogeneity of variance with the Shapiro–Wilk test and Levene’s test. When the assumptions were not met, a log transformation was used. Consequently, a two-way ANOVA was performed for each photosynthetic parameter with treatment (SA or WA) and time (48 h, 96 h, and 192 h) as factors, followed by a post hoc analysis with Tukey’s honest significant difference method with the R package ‘multcomp’. Values were considered significantly different at p < 0.05.
5. Conclusions
Our data show that SA-sprayed leaves under water deficit conditions (192 h after watering) had an improved osmotic potential and maintained a higher leaf water content that enabled an enhanced PSII function, compared to WA-sprayed leaves that retained lower leaf water content. Our hypothesis that SA would improve the light energy utilization efficiency in celery plants by boosting the dissipation of excess excitation energy via the NPQ mechanism, thus enhancing PSII function and improving drought stress tolerance, was confirmed. It seems that under water deficit stress, SA activates a regulatory network of stress and light energy partition signaling that can mitigate water deficit stress on the PSII function; but under non-stress conditions, at least in celery plants, it reduces PSII efficiency. SA-sprayed celery leaves were less efficient in utilizing light energy up to 96 h after watering compared to WA-sprayed leaves. This trend was reversed 192 h after watering (water deficit conditions) when SA induced the NPQ mechanism to protect the chloroplast from photo-oxidative damage. This was accomplished by dissipating the excess light energy as heat and, thus, restoring the drought stress-induced decrease in PSII efficiency. The drought resilience of PSII in SA-sprayed leaves was achieved, especially at HL conditions, through the ability of the SA-sprayed leaves to retain an equal fraction of open PSII RCs (qp) as in non-stress conditions.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms25126721/s1. References [122,123,124,125,126] are cited in the supplementary materials.
Author Contributions
Conceptualization, M.M. and I.S.; methodology, E.P., J.M., T.A. and I.S.; validation, M.M., E.P., J.M., T.A., G.B. and I.S.; formal analysis, M.M., E.P., J.M., T.A., G.B. and I.S.; investigation, M.M., E.P., J.M., T.A., G.B. and I.S.; resources, M.M. and E.P.; data curation, E.P., J.M., T.A. and I.S.; writing—original draft preparation, M.M., E.P. and I.S.; writing—review and editing, M.M., E.P., J.M., T.A., G.B. and I.S.; visualization, M.M., E.P. and I.S.; supervision, M.M., E.P. and I.S.; project administration, M.M., E.P. and I.S. All authors have read and agreed to the published version of the manuscript.
Funding
E.P. is supported by the AUTh Research Committee, grant No. 91913, through funds of Schur Flexibles ABR SA.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fregonezi, B.F.; Pereira, A.E.S.; Ferreira, J.M.; Fraceto, L.F.; Gomes, D.G.; Oliveira, H.C. Seed priming with nanoencapsulated gibberellic acid triggers beneficial morphophysiological and biochemical responses of tomato plants under different water conditions. Agronomy 2024, 14, 588. [Google Scholar] [CrossRef]
- Wing, I.S.; De Cian, E.; Mistry, M.N. Global vulnerability of crop yields to climate change. J. Environ. Econ. Manag. 2021, 109, 102462. [Google Scholar] [CrossRef]
- Placide, R.; Hirut, G.B.; Stephan, N.; Fekadu, B. Assessment of drought stress tolerance in root and tuber crops. Afr. J. Plant Sci. 2014, 8, 214–224. [Google Scholar] [CrossRef][Green Version]
- Sperdouli, I.; Mellidou, I.; Moustakas, M. Harnessing chlorophyll fluorescence for phenotyping analysis of wild and cultivated tomato for high photochemical efficiency under water deficit for climate change resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Moustaka, J. Early drought stress warning: Color pictures of photosystem II photochemistry. Climate 2022, 10, 179. [Google Scholar] [CrossRef]
- Moustaka, J.; Moustakas, M. Early-stage detection of biotic and abiotic stress on plants by chlorophyll fluorescence imaging analysis. Biosensors 2023, 13, 796. [Google Scholar] [CrossRef]
- Akin, S.; Kaya, C. Impact of salicylic acid and sodium hydrosulfide applied singly or in combination on drought tolerance and grain yield in wheat plants. Food Energy Secur. 2024, 13, e532. [Google Scholar] [CrossRef]
- Zhang, J.; Corpas, F.J.; Li, J.; Xie, Y. Hydrogen sulfide and reactive oxygen species, antioxidant defense, abiotic stress tolerance mechanisms in plants. Int. J. Mol. Sci. 2022, 23, 9463. [Google Scholar] [CrossRef]
- Li, J.; Lardon, R.; Mangelinckx, S.; Geelen, D. Practical guide toward discovery of biomolecules with biostimulant activity. J. Exp. Bot. 2024; in press. [Google Scholar] [CrossRef]
- Martinez, C.; Pons, E.; Prats, G.; Leon, J. Salicylic acid regulates flowering time and links defence responses and reproductive development. Plant J. 2004, 37, 209–217. [Google Scholar] [CrossRef]
- Stevens, J.; Senaratna, T.; Sivasithamparam, K. Salicylic acid induces salinity tolerance in tomato (Lycopersicon esculentum cv. Roma): Associated changes in gas exchange, water relations and membrane stabilisation. Plant Growth Regul. 2006, 49, 77–83. [Google Scholar]
- Ramirez, A.; Rodriguez, D.; Reyes, D.; Angel Jimenez, J.; Nicolas, G.; Lopez Climent, M.; Gómez-Cadenas, A.; Nicolás, C. Evidence for a role of gibberellins in salicylic acid-modulated early plant responses to abiotic stress in Arabidopsis seeds. Plant Physiol. 2009, 150, 1335–1344. [Google Scholar] [CrossRef]
- Ding, P.; Ding, Y. Stories of salicylic acid: A plant defense hormone. Trends Plant Sci. 2020, 25, 549–565. [Google Scholar] [CrossRef]
- Koo, Y.M.; Heo, A.Y.; Choi, H.W. Salicylic acid as a safe plant protector and growth regulator. Plant Pathol. J. 2020, 36, 1–10. [Google Scholar] [CrossRef]
- Li, A.; Sun, X.; Liu, L. Action of salicylic acid on plant growth. Front. Plant Sci. 2022, 13, 878076. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, Z.; Chu, Z. Emerging roles of salicylic acid in plant saline stress tolerance. Int. J. Mol. Sci. 2023, 24, 3388. [Google Scholar] [CrossRef]
- Wu, X.; Li, J.; Song, L.Y.; Zeng, L.L.; Guo, Z.J.; Ma, D.N.; Wei, M.Y.; Zhang, L.D.; Wang, X.X.; Zheng, H.L. NADPH oxidase-dependent H2O2 production mediates salicylic acid-induced salt tolerance in mangrove plant Kandelia obovata by regulating Na+/K+ and redox homeostasis. Plant J. 2024, 118, 1119–1135. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Zhong, Q.; Hu, H.; Fan, B.; Zhu, C.; Chen, Z. Biosynthesis and roles of salicylic acid in balancing stress response and growth in plants. Int. J. Mol. Sci. 2021, 22, 11672. [Google Scholar] [CrossRef]
- Hanif, S.; Mahmood, A.; Javed, T.; Bibi, S.; Zia, M.A.; Asghar, S.; Naeem, Z.; Ercisli, S.; Rahimi, M.; Ali, B. Exogenous application of salicylic acid ameliorates salinity stress in barley (Hordeum vulgare L.). BMC Plant Biol. 2024, 24, 270. [Google Scholar] [CrossRef]
- Jia, X.; Wang, L.; Zhao, H.; Zhang, Y.; Chen, Z.; Xu, L.; Yi, K. The origin and evolution of salicylic acid signaling and biosynthesis in plants. Mol. Plant 2023, 16, 245–259. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, W.; Zhao, Q. Salicylic acid biosynthesis is not from phenylalanine in Arabidopsis. J. Integr. Plant Biol. 2023, 65, 881–887. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Ann. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Mimouni, H.; Wasti, S.; Manaa, A.; Gharbi, E.; Chalh, A.; Vandoorne, B.; Lutts, S.; Ben Ahmed, H. Does salicylic acid (SA) improve tolerance to salt stress in plants? A study of SA effects on tomato plant growth, water dynamics, photosynthesis, and biochemical parameters. OMICS 2016, 20, 180–910. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.M.S.; Hussain, H.A.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Role of exogenous-applied salicylic acid, zinc and glycine betaine to improve drought-tolerance in wheat during reproductive growth stages. BMC Plant Biol. 2021, 21, 574. [Google Scholar] [CrossRef]
- Janda, T.; Gondor, O.K.; Yordanova, R.; Szalai, G.; Pál, M. Salicylic acid and photosynthesis: Signalling and effects. Acta Physiol. Plant. 2014, 36, 2537–2546. [Google Scholar] [CrossRef]
- Khan, W.; Prithiviraj, B.; Smith, D.L. Photosynthetic responses of corn and soybean to foliar application of salicylates. J. Plant Physiol. 2003, 160, 485–492. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Tari, I.; Páldi, E. Hydroponic treatment with salicylic acid decreases the effects of chilling injury in maize (Zea mays L.) plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Ananieva, E.A.; Alexieva, V.S.; Popova, L.P. Treatment with salicylic acid decreases the effects of paraquat on photosynthesis. J. Plant Physiol. 2002, 159, 685–693. [Google Scholar] [CrossRef]
- Henschel, J.M.; Dantas, E.F.O.; Soares, V.A.; Santos, S.K.D.; Santos, L.W.O.D.; Dias, T.J.; Batista, D.S. Salicylic acid mitigates the effects of mild drought stress on radish (Raphanus sativus) growth. Funct. Plant Biol. 2022, 49, 822–831. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustaka, J.; Ouzounidou, G.; Moustakas, M. Leaf age-dependent photosystem II photochemistry and oxidative stress responses to drought stress in Arabidopsis thaliana are modulated by flavonoid accumulation. Molecules 2021, 26, 4157. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Ouzounidou, G.; Moustakas, M. Hormesis responses of photosystem II in Arabidopsis thaliana under water deficit stress. Int. J. Mol. Sci. 2023, 24, 9573. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Moustakas, M. Plant Photochemistry, Reactive Oxygen Species, and Photoprotection. Photochem 2022, 2, 5–8. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef]
- Wu, A.; Hammer, G.L.; Doherty, A.; von Caemmerer, S.; Farquhar, G.D. Quantifying impacts of enhancing photosynthesis on crop yield. Nat. Plants 2019, 5, 380388. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.J.; Masclaux-Daubresse, C.; Strittmatter, G.; Weber, A.P.M.; Taylor, S.H.; Harbinson, J.; Yin, X.; Long, S.; Paul, M.J.; Westhoff, P.; et al. Improving crop yield potential: Underlying biological processes and future prospects. Food Energy Secur. 2023, 12, e435. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Gaju, O.; Bowerman, A.F.; Buck, S.A.; Evans, J.R.; Furbank, R.T.; Gilliham, M.; Millar, A.H.; Pogson, B.J.; Reynolds, M.P.; et al. Enhancing crop yields through improvements in the efficiency of photosynthesis and respiration. New Phytol. 2023, 237, 60–77. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Ruban, A.V. Dynamic non-photochemical quenching in plants: From molecular mechanism to productivity. Plant J. 2020, 101, 885–896. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean photosynthesis and crop yield are improved by accelerating recovery from photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Enhancing the light reactions of photosynthesis: Strategies, controversies, and perspectives. Mol. Plant. 2023, 16, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D. Perspectives of improving rice photosynthesis for higher grain yield. Crop Environ. 2024, 3, 123–137. [Google Scholar] [CrossRef]
- Poór, P.; Tari, I. Regulation of stomatal movement and photosynthetic activity in guard cells of tomato abaxial epidermal peels by salicylic acid. Funct. Plant Biol. 2012, 39, 1028–1037. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Adamakis, I.-D.S.; Moustaka, J.; İşgören, S.; Şaş, B. Harnessing the role of foliar applied salicylic acid in decreasing chlorophyll content to reassess photosystem II photoprotection in crop plants. Int. J. Mol. Sci. 2022, 23, 7038. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Moustaka, J.; Şaş, B.; İşgören, S.; Morales, F. Mechanistic insights on salicylic acid mediated enhancement of photosystem II function in oregano seedlings subjected to moderate drought stress. Plants 2023, 12, 518. [Google Scholar] [CrossRef] [PubMed]
- Janda, K.; Hideg, E.; Szalai, G.; Kovács, L.; Janda, T. Salicylic acid may indirectly influence the photosynthetic electron transport. J. Plant Physiol. 2012, 169, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Chen, Y.H.; Ortega, M.A.; Tsai, C.J. The diversity of salicylic acid biosynthesis and defense signaling in plants: Knowledge gaps and future opportunities. Curr. Opin. Plant Biol. 2023, 72, 102349. [Google Scholar] [CrossRef] [PubMed]
- Fragkiska, M. Wild and cultivated vegetables, herbs and spices in Greek antiquity (900 B.C. to 400 B.C.). Environ. Archaeol. 2013, 10, 73–82. [Google Scholar]
- Liu, J.X.; Wang, H.; Feng, K.; Li, T.; Liu, Y.H.; Duan, A.Q.; Shu, S.; Liu, H.; Xiong, A.S. AgDHAR2, a chloroplast-located dehydroascorbate reductase, modulates the ascorbate accumulation and drought stress response in celery. Environ. Exp. Bot. 2022, 202, 105006. [Google Scholar] [CrossRef]
- Li, M.Y.; Hou, X.L.; Wang, F.; Tan, G.F.; Xu, Z.S.; Xiong, A.S. Advances in the research of celery, an important Apiaceae vegetable crop. Crit. Rev. Biotechnol. 2018, 38, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, N.; Bemani, N.M.; Mohammadinejad, A.; Mohajeri, S.A. Beneficial effects of celery (Apium graveolens) on metabolic syndrome: A review of the existing evidences. Phytother. Res. 2019, 33, 3040–3053. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, J.; Wang, C.; Han, K.; Hu, L.; Niu, T.; Yang, Y.; Chang, Y.; Xie, J. Exogenous proline enhances systemic defense against salt stress in celery by regulating photosystem, phenolic compounds, and antioxidant system. Plants 2023, 12, 928. [Google Scholar] [CrossRef]
- Ma, J.Z.; Zhang, M.; Liu, Z.G.; Wang, M.; Sun, Y.; Zheng, W.K.; Lu, H. Copper-based-zinc-boron foliar fertilizer improved yield, quality, physiological characteristics, and microelement concentration of celery (Apium graveolens L.). Environ. Pollut. Bioavailab. 2019, 31, 261–271. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. J. Plant Physiol. 2012, 169, 577–585. [Google Scholar] [CrossRef]
- Smeekens, S. Drought resistance: Spraying for yield. Nat. Plants 2017, 3, 17023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Rosental, L.; Ji, B.; Brotman, Y.; Dai, M. Metabolite-mediated adaptation of crops to drought and the acquisition of tolerance. Plant J. 2024, 118, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Loutfy, N.; El-Tayeb, M.A.; Hassanen, A.M.; Moustafa, M.F.; Sakuma, Y.; Inouhe, M. Changes in the water status and osmotic solute contents in response to drought and salicylic acid treatments in four different cultivars of wheat (Triticum aestivum). J. Plant Res. 2012, 125, 173–184. [Google Scholar] [PubMed]
- Shi, Q.; Bao, Z.; Zhu, Z.; Ying, Q.; Qian, Q. Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence and antioxidant enzyme activity in seedlings of Cucumis sativa L. Plant Growth Regul. 2006, 48, 127–135. [Google Scholar] [CrossRef]
- Abdelhameed, R.E.; Abdel Latef, A.A.H.; Shehata, R.S. Physiological responses of salinized fenugreek (Trigonella foenum-graecum L.) plants to foliar application of salicylic acid. Plants 2021, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Yang, S.; Chen, Y. Salicylic acid alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in two melon cultivars (Cucumis melo L.). Protoplasma 2015, 252, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Huang, X.; Xing, J.; Yao, J.; Yin, T.; Jiang, J.; Wang, P.; Xu, B. STAYGREEN-mediated chlorophyll a catabolism is critical for photosystem stability during heat-induced leaf senescence in perennial ryegrass. Plant Cell Environ. 2022, 45, 1412–1427. [Google Scholar] [CrossRef]
- Pancheva, T.V.; Popova, L.P.; Uzunova, A.M. Effect of salicylic acid on growth and photosynthesis in barley plants. J. Plant Physiol. 1996, 149, 57–63. [Google Scholar] [CrossRef]
- Anandhi, S.; Ramanujam, M.P. Effect of salicylic acid on black gram (Vigna mungo) cultivars. Ind. J. Plant Physiol. 1997, 2, 138–141. [Google Scholar]
- Chandra, A.; Bhatt, R.K. Biochemical and physiological response to salicylic acid in relation to the systemic acquired resistance. Photosynthetica 1998, 45, 255–258. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Hayat, S.; Ahmad, A. Salicylic acid influences net photosynthetic rate, carboxylation efficiency, nitrate reductase activity and seed yield in Brassica juncea. Photosynthetica 2003, 41, 281–284. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Arfan, M.; Athar, H.R.; Ashraf, M. Does exogenous application of salicylic acid through the rooting medium modulate growth and photosynthetic capacity in two differently adapted spring wheat cultivars under salt stress? J. Plant Physiol. 2007, 164, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Torun, H.; Novák, O.; Mikulík, J.; Strnad, M.; Ayaz, F.A. The effects of exogenous salicylic acid on endogenous phytohormone status in Hordeum vulgare L. under salt stress. Plants 2022, 11, 618. [Google Scholar] [CrossRef]
- Krieger-Liszkay, A. Singlet oxygen production in photosynthesis. J. Exp. Bot. 2005, 56, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.E.; Ivanov, A.G.; Öquist, G.; Grodzinski, B.; Sarhan, F.; Huner, N.P.A. Energy balance, organellar redox status, and acclimation to environmental stress. Can. J. Bot. 2006, 84, 1355–1370. [Google Scholar] [CrossRef]
- Gawroński, P.; Witoń, D.; Vashutina, K.; Bederska, M.; Betliński, B.; Rusaczonek, A.; Karpiński, S. Mitogen-activated protein kinase 4 is a salicylic acid-independent regulator of growth but not of photosynthesis in Arabidopsis. Mol. Plant 2014, 7, 1151–1166. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PSII quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Kasajima, I.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H. Estimation of the relative sizes of rate constants for chlorophyll de-excitation processes through comparison of inverse fluorescence intensities. Plant Cell Physiol. 2009, 50, 1600–1616. [Google Scholar] [CrossRef]
- Moustakas, M.; Dobrikova, A.; Sperdouli, I.; Hanć, A.; Moustaka, J.; Adamakis, I.-D.S.; Apostolova, E. Photosystem II tolerance to excess zinc exposure and high light stress in Salvia sclarea L. Agronomy 2024, 14, 589. [Google Scholar] [CrossRef]
- Mattila, H.; Sotoudehnia, P.; Kuuslampi, T.; Stracke, R.; Mishra, K.B.; Tyystjärvi, E. Singlet oxygen, flavonols and photoinhibition in green and senescing silver birch leaves. Trees 2021, 35, 1267–1282. [Google Scholar] [CrossRef]
- Adir, N.; Zer, H.; Shochat, S.; Ohad, I. Photoinhibition–a historical perspective. Photosynth. Res. 2003, 76, 343–370. [Google Scholar] [CrossRef]
- Li, L.; Aro, E.M.; Millar, A.H. Mechanisms of photodamage and protein turnover in photoinhibition. Trends Plant Sci. 2018, 23, 667–676. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 655–684. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Photosystem-II damage and repair cycle in chloroplasts: What modulates the rate of photodamage in vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D. The photoprotective molecular switch in the photosystem II antenna. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 167–181. [Google Scholar] [CrossRef]
- Zavafer, A.; Mancilla, C. Concepts of photochemical damage of photosystem II and the role of excessive excitation. J. Photochem. Photobiol. C Photochem. Rev. 2021, 47, 100421. [Google Scholar] [CrossRef]
- Yotsova, E.K.; Dobrikova, A.G.; Stefanov, M.A.; Kouzmanova, M.; Apostolova, E.L. Improvement of the rice photosynthetic apparatus defence under cadmium stress modulated by salicylic acid supply to roots. Theor. Exp. Plant Physiol. 2018, 30, 57–70. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Aziz, U.; Alsahli, A.A.; Alyemeni, M.N.; Ahmad, P. Influence of exogenous salicylic acid and nitric oxide on growth, photosynthesis, and ascorbate-glutathione cycle in salt stressed Vigna angularis. Biomolecules 2020, 10, 42. [Google Scholar] [CrossRef]
- Maslenkova, L.; Peeva, V.; Stojnova, Z.; Popova, L. Salicylic acid-induced changes in photosystem II reactions in barley plants. Biotechnol. Biotechnol. Equip. 2009, 23, 297–300. [Google Scholar] [CrossRef]
- Goggin, F.L.; Fischer, H.D. Singlet oxygen signalling and its potential roles in plant biotic interactions. Plant Cell Environ. 2024, 47, 1957–1970. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Moustaka, J.; Tanou, G.; Adamakis, I.D.; Eleftheriou, E.P.; Moustakas, M. Leaf age dependent photoprotective and antioxidative mechanisms to paraquat-induced oxidative stress in Arabidopsis thaliana. Int. J. Mol. Sci. 2015, 16, 13989–14006. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Sperdouli, I.; Adamakis, I.-D.S.; Şaş, B.; İşgören, S.; Moustaka, J.; Morales, F. Mechanistic approach on melatonin-induced hormesis of photosystem II function in the medicinal plant Mentha spicata. Plants 2023, 12, 4025. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Ruban, A.V.; Noctor, G. Viewing oxidative stress through the lens of oxidative signalling rather than damage. Biochem. J. 2017, 474, 877–883. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Adamakis, I.D.S. Editorial: Reactive oxygen species in chloroplasts and chloroplast antioxidants under abiotic stress. Front. Plant Sci. 2023, 14, 1208247. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signaling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Miloslavina, Y.; Jahns, P.; Holzwarth, A.R. On the relationship between non-photochemical quenching and photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Hanć, A.; Dobrikova, A.; Sperdouli, I.; Adamakis, I.D.S.; Apostolova, E. Spatial heterogeneity of cadmium effects on Salvia sclarea leaves revealed by chlorophyll fluorescence imaging analysis and laser ablation inductively coupled plasma mass spectrometry. Materials 2019, 12, 2953. [Google Scholar] [CrossRef]
- Pfalz, J.; Liebers, M.; Hirth, M.; Grübler, B.; Holtzegel, U.; Schröter, Y.; Dietzel, L.; Pfannschmidt, T. Environmental control of plant nuclear gene expression by chloroplast redox signals. Front. Plant Sci. 2012, 3, 257. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J.; Turkan, I.; Krieger-Liszkay, A. Redox- and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef]
- Leverne, L.; Roach, T.; Perreau, F.; Maignan, F.; Krieger-Liszkay, A. Increased drought resistance in state transition mutants is linked to modified plastoquinone pool redox state. Plant Cell Environ. 2023, 46, 3737–3747. [Google Scholar] [CrossRef] [PubMed]
- Bräutigam, K.; Dietzel, L.; Kleine, T.; Ströher, E.; Wormuth, D.; Dietz, K.J.; Radke, D.; Wirtz, M.; Hell, R.; Dörmann, P.; et al. Dynamic plastid redox signals integrate gene expression and metabolism to induce distinct metabolic states in photosynthetic acclimation in Arabidopsis. Plant Cell 2009, 21, 2715–2732. [Google Scholar] [CrossRef]
- Dietz, K.J.; Pfannschmidt, T. Novel regulators in photosynthetic redox control of plant metabolism and gene expression. Plant Physiol. 2011, 155, 1477–1485. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustaka, J.; Antonoglou, O.; Adamakis, I.D.S.; Dendrinou-Samara, C.; Moustakas, M. Leaf age dependent effects of foliar-sprayed CuZn nanoparticles on photosynthetic efficiency and ROS generation in Arabidopsis thaliana. Materials 2019, 12, 2498. [Google Scholar] [CrossRef] [PubMed]
- Borisova-Mubarakshina, M.M.; Vetoshkina, D.V.; Ivanov, B.N. Antioxidant and signaling functions of the plastoquinone pool in higher plants. Physiol. Plant. 2019, 166, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Messant, M.; Krieger-Liszkay, A.; Shimakawa, G. Dynamic changes in protein-membrane association for regulating photosynthetic electron transport. Cells 2021, 10, 1216. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Mao, H.T.; Wu, N.; Mohi Ud Din, A.; Khan, A.; Zhang, H.Y.; Yuan, S. Salicylic acid protects photosystem II by alleviating photoinhibition in Arabidopsis thaliana under high light. Int. J. Mol. Sci. 2020, 21, 1229. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Panteris, E.; Moustaka, J.; Aydın, T.; Bayçu, G.; Moustakas, M. Mechanistic insights on salicylic acid-induced enhancement of photosystem II function in basil plants under non-stress or mild drought stress. Int. J. Mol. Sci. 2024, 25, 5728. [Google Scholar] [CrossRef]
- Ali, A.; Kant, K.; Kaur, N.; Gupta, S.; Jindal, P.; Gill, S.S.; Naeem, M. Salicylic acid: Homeostasis, signalling and phytohormone crosstalk in plants under environmental challenges. S. Afr. J. Bot. 2024, 169, 314–335. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Spatio-temporal heterogeneity in Arabidopsis thaliana leaves under drought stress. Plant Biol. 2012, 14, 118–128. [Google Scholar] [CrossRef]
- Borek, M.; Bączek-Kwinta, R.; Rapacz, M. Photosynthetic activity of variegated leaves of Coleus × hybridus hort. cultivars characterised by chlorophyll fluorescence techniques. Photosynthetica 2016, 54, 331–339. [Google Scholar] [CrossRef]
- Moustaka, J.; Panteris, E.; Adamakis, I.D.S.; Tanou, G.; Giannakoula, A.; Eleftheriou, E.P.; Moustakas, M. High anthocyanin accumulation in poinsettia leaves is accompanied by thylakoid membrane unstacking, acting as a photoprotective mechanism, to prevent ROS formation. Environ. Exp. Bot. 2018, 154, 44–55. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Hendrickson, L.; Furbank, R.T.; Chow, W.S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fluorescence. Photosynth. Res. 2004, 82, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a non-intrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Series Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1994; Volume 100, pp. 49–70. [Google Scholar]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Schreiber, U.; Bock, M. Determination of the quantum efficiency of photosystem II and of non-photochemical quenching of chlorophyll fluorescence in the field. Oecologia 1995, 102, 425–432. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).