
Arsenic Nanoparticles Trigger Apoptosis via Anoikis Induction in OECM-1 Cells

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. AsNP Preparation and Characterization
2.2. AsNPs Inhibit OECM-1 Cell Viability by Compromising Cell Adhesion
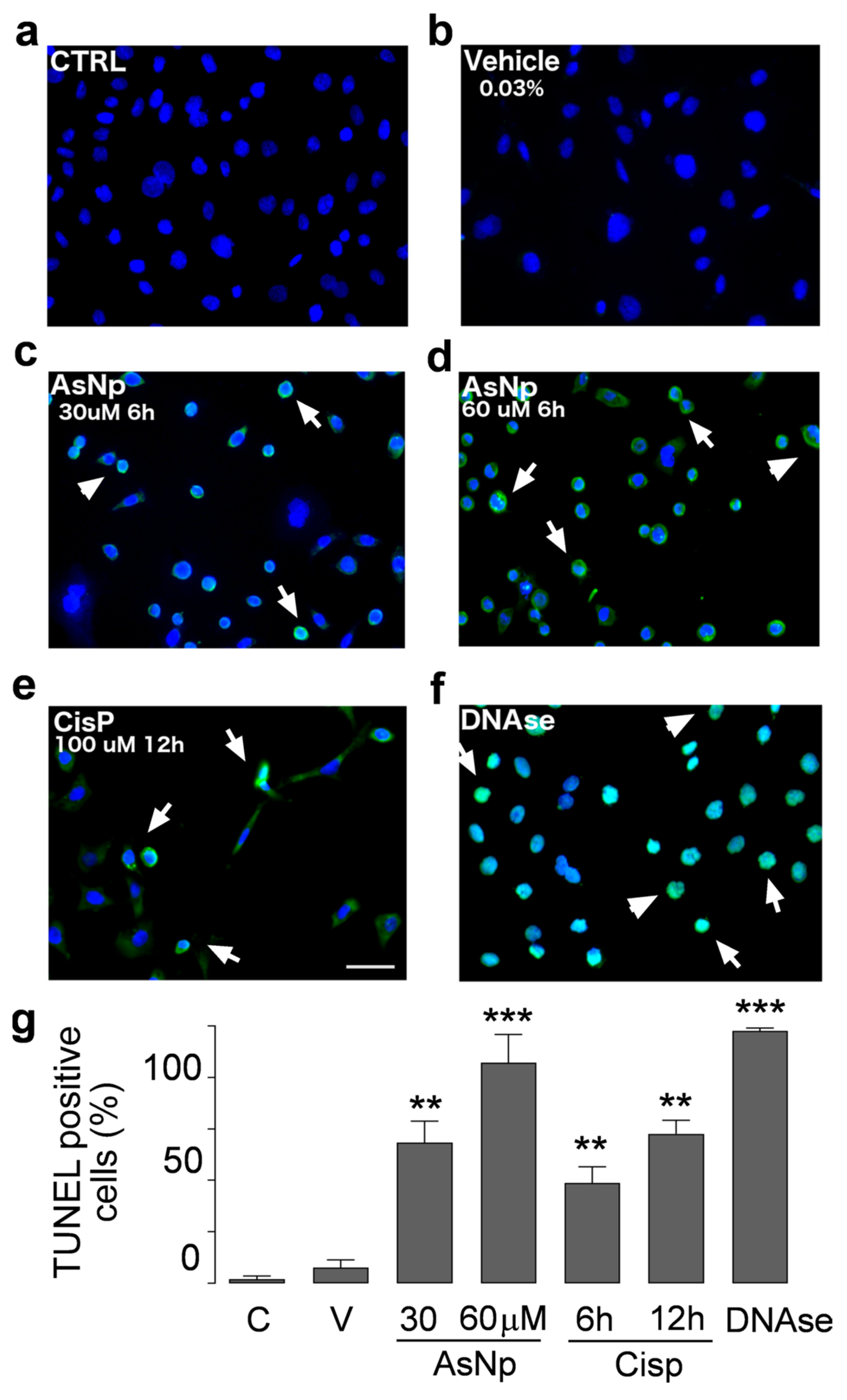
2.3. AsNPs Induce Apoptosis in OECM-1 Cells
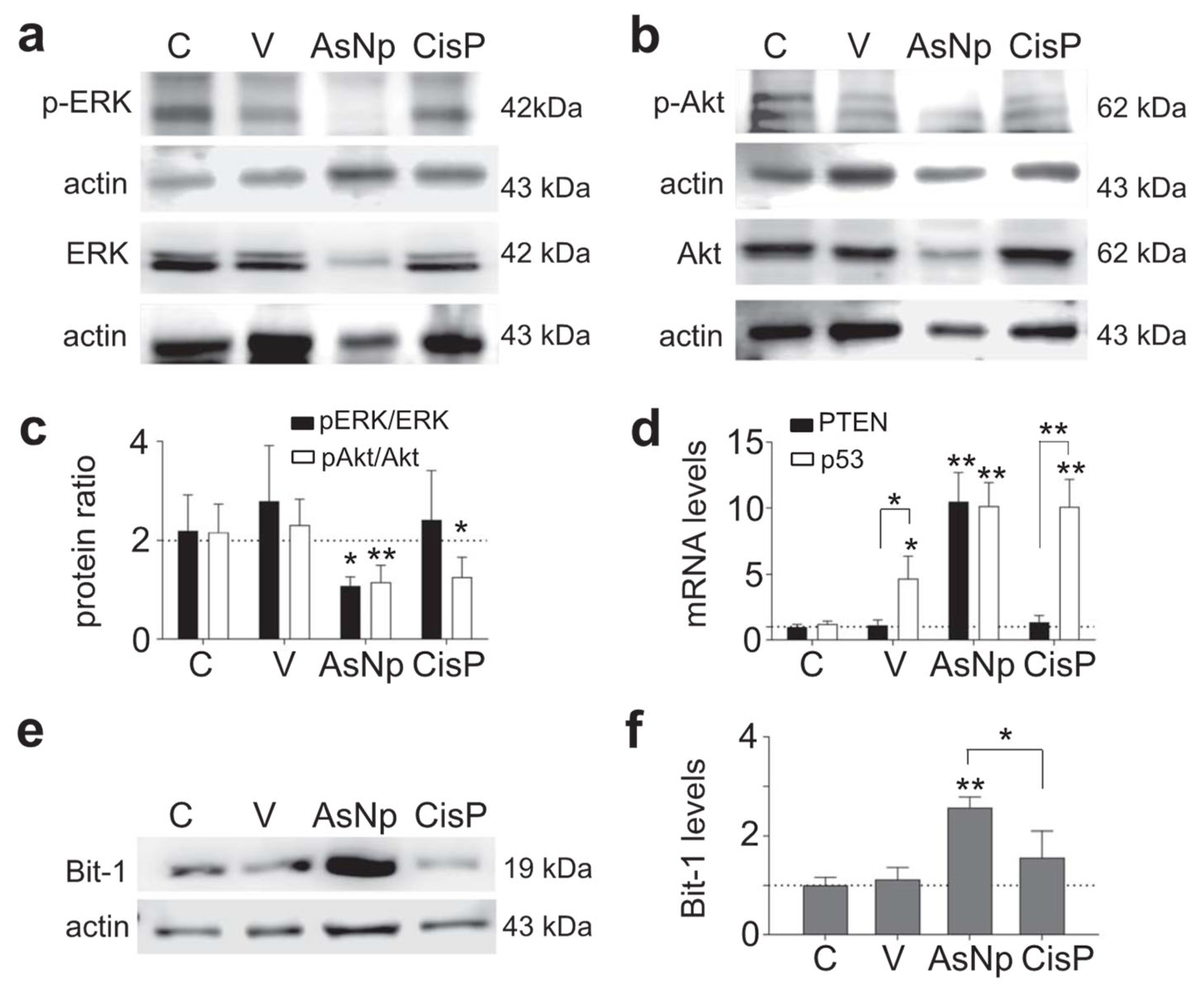
2.4. AsNPs Induce Anoikis Signaling in OECM-1 Cells
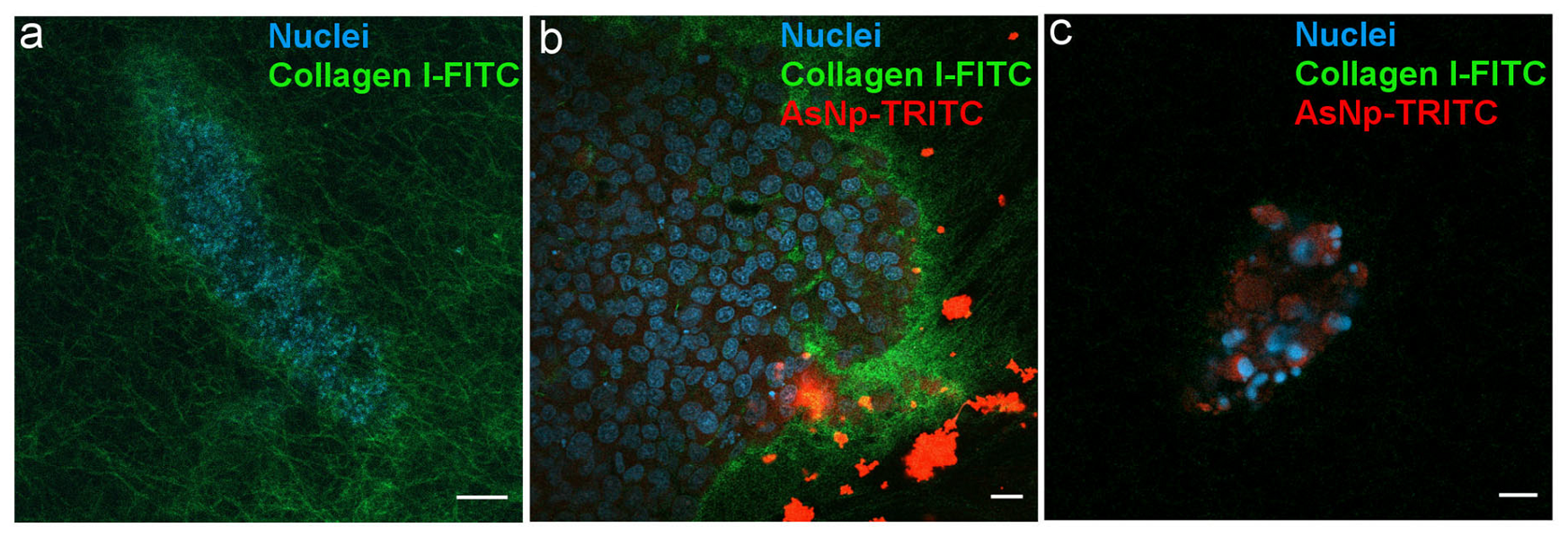
2.5. Interactions of AsNPs with OECM-1 Spheroids
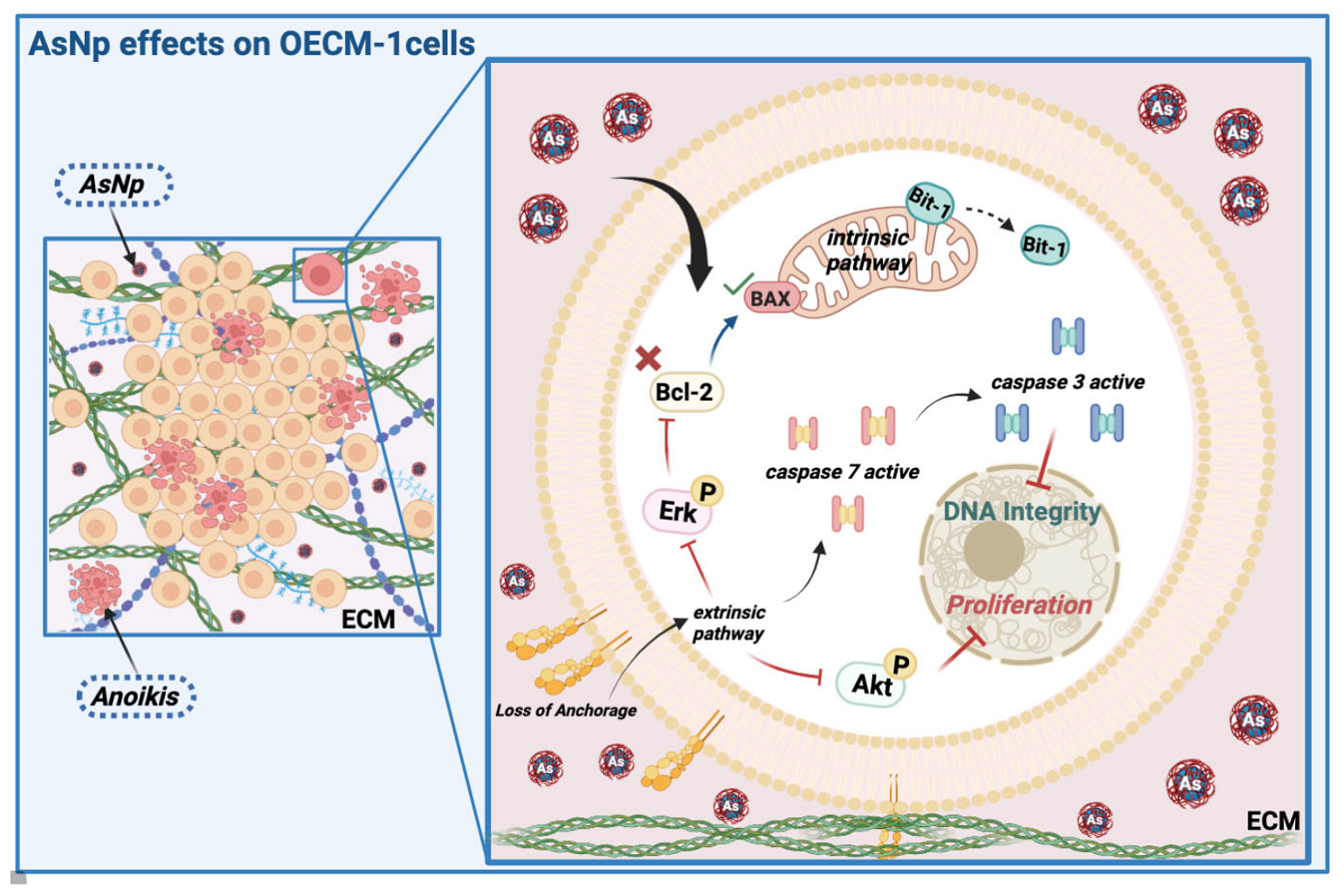
3. Discussion
4. Materials and Methods
4.1. Nanoparticles Preparation and Characterization
4.2. Cell Culture
4.3. Resazurin Reduction Assay
4.4. Methylene Blue Staining
4.5. Immunoblotting
4.6. Caspase 3/7 Activity Assay
4.7. TUNEL Assay
4.8. qPCR
4.9. 3D Culture of OECM-1 Cells Spheroids
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhatnagar, S.; Venuganti, V.V.K. Cancer Targeting: Responsive Polymers for Stimuli-Sensitive Drug Delivery. J. Nanosci. Nanotechnol. 2015, 15, 1925–1945. [Google Scholar] [CrossRef] [PubMed]
- Baetke, S.C.; Lammers, T.; Kiessling, F. Applications of Nanoparticles for Diagnosis and Therapy of Cancer. Br. J. Radiol. 2015, 88, 20150207. [Google Scholar] [CrossRef] [PubMed]
- Baláž, P.; Sedlák, J. Arsenic in Cancer Treatment: Challenges for Application of Realgar Nanoparticles (A Minireview). Toxins 2010, 2, 1568–1581. [Google Scholar] [CrossRef] [PubMed]
- Majeed, S.; bin Abdullah, M.S.; Nanda, A.; Ansari, M.T. In Vitro Study of the Antibacterial and Anticancer Activities of Silver Nanoparticles Synthesized from Penicillium Brevicompactum (MTCC-1999). J. Taibah Univ. Sci. 2016, 10, 614–620. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Tay, C.Y.; Bay, B.H.; Leong, D.T. Gold Nanoparticles Induced Endothelial Leakiness Depends on Particle Size and Endothelial Cell Origin. ACS Nano 2017, 11, 5020–5030. [Google Scholar] [CrossRef] [PubMed]
- Tay, C.Y.; Setyawati, M.I.; Leong, D.T. Nanoparticle Density: A Critical Biophysical Regulator of Endothelial Permeability. ACS Nano 2017, 11, 2764–2772. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, L.; Peng, F.; Shi, X.; Leong, D.T. Targeting Endothelial Cell Junctions with Negatively Charged Gold Nanoparticles. Chem. Mater. 2018, 30, 3759–3767. [Google Scholar] [CrossRef]
- Ge, H.; Wang, D.; Pan, Y.; Guo, Y.; Li, H.; Zhang, F.; Zhu, X.; Li, Y.; Zhang, C.; Huang, L. Sequence-Dependent DNA Functionalization of Upconversion Nanoparticles and Their Programmable Assemblies. Angew. Chem.-Int. Ed. 2020, 59, 8133–8137. [Google Scholar] [CrossRef]
- Ganbold, T.; Han, S.; Hasi, A.; Baigude, H. Receptor-Mediated Delivery of Therapeutic RNA by Peptide Functionalized Curdlan Nanoparticles. Int. J. Biol. Macromol. 2019, 126, 633–640. [Google Scholar] [CrossRef]
- Jia, X.; Guo, M.; Han, Q.; Tian, Y.; Yuan, Y.; Wang, Z.; Qian, Y.; Wang, W. Synergetic Tumor Probes for Facilitating Therapeutic Delivery by Combined-Functionalized Peptide Ligands. Anal. Chem. 2020, 92, 5650–5655. [Google Scholar] [CrossRef]
- Hongy, Z.; Xiaoy, M.; Yang, Y.; Han, Z.; Cao, Y.; Li, C.; Wu, Y.; Gong, Q.; Zhou, X.; Xu, D.; et al. Arsenic Disulfide Synergizes with the Phosphoinositide 3-Kinase Inhibitor PI-103 to Eradicate Acute Myeloid Leukemia Stem Cells by Inducing Differentiation. Carcinogenesis 2011, 32, 1550–1558. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.C.; Costa, P.J.; Velho, S.; Amaral, M.H. Functionalizing Nanoparticles with Cancer-Targeting Antibodies: A Comparison of Strategies. J. Control. Release 2020, 320, 180–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Xu, R.; Yan, L.; Wu, Z.; Wei, Y.; Zhao, W.; Wang, X.; Xie, Q.; Li, H. Properties of Realgar Bioleaching Using an Extremely Acidophilic Bacterium and Its Antitumor Mechanism as an Anticancer Agent. Biol. Res. 2017, 50, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Waxman, S.; Anderson, K.C. History of the Development of Arsenic Derivatives in Cancer Therapy. Oncologist 2001, 6, 3–10. [Google Scholar] [CrossRef]
- Miodragović, Ð.; Swindell, E.P.; Sattar Waxali, Z.; Bogachkov, A.; O’Halloran, T.V. Beyond Cisplatin: Combination Therapy with Arsenic Trioxide. Inorganica Chim. Acta 2019, 496, 119030. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Kantarjian, H.; Ravandi, F. Acute Promyelocytic Leukemia Current Treatment Algorithms. Blood Cancer J. 2021, 11, 123. [Google Scholar] [CrossRef]
- Alimoghaddam, K. A Review of Arsenic Trioxide and Acute Promyelocytic Leukemia. Int. J. Hematol.-Oncol. Stem Cell Res. 2014, 8, 44–54. [Google Scholar]
- Sönksen, M.; Kerl, K.; Bunzen, H. Current Status and Future Prospects of Nanomedicine for Arsenic Trioxide Delivery to Solid Tumors. Med. Res. Rev. 2022, 42, 374–398. [Google Scholar] [CrossRef]
- Subastri, A.; Arun, V.; Sharma, P.; Preedia babu, E.; Suyavaran, A.; Nithyananthan, S.; Alshammari, G.M.; Aristatile, B.; Dharuman, V.; Thirunavukkarasu, C. Synthesis and Characterisation of Arsenic Nanoparticles and Its Interaction with DNA and Cytotoxic Potential on Breast Cancer Cells. Chem. Biol. Interact. 2018, 295, 73–83. [Google Scholar] [CrossRef]
- Dong, X.; Ma, N.; Liu, M.; Liu, Z. Effects of As2O3 Nanoparticles on Cell Growth and Apoptosis of NB4 Cells. Exp. Ther. Med. 2015, 10, 1271–1276. [Google Scholar] [CrossRef]
- Pastorek, S.J.; Gronesova, M.; Cholujova, P.; Hunakova, D.; Bujnakova, L.; Balaz, Z.; Duraj, P.; Lee, J. Realgar (As4S4) nanoparticles and arsenic trioxide (As2O3) induced autophagy and apoptosis in human melanoma cells in vitro. Neoplasma 2013, 60, 607–616. [Google Scholar] [CrossRef]
- Hu, J.; Dong, Y.; Ding, L.; Dong, Y.; Wu, Z.; Wang, W.; Shen, M.; Duan, Y. Local Delivery of Arsenic Trioxide Nanoparticles for Hepatocellular Carcinoma Treatment. Signal Transduct. Target. Ther. 2019, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, H. Development of Realgar Nanotherapeutics for Cancer Treatments. In Nanotechnology in Regenerative Medicine and Drug Delivery Therapy; Springer: Singapore, 2020; pp. 421–454. [Google Scholar]
- Patil, S.; Chandrasekaran, R. Biogenic Nanoparticles: A Comprehensive Perspective in Synthesis, Characterization, Application and Its Challenges. J. Genet. Eng. Biotechnol. 2020, 18, 67. [Google Scholar] [CrossRef]
- Mughal, B.; Zaidi, S.Z.J.; Zhang, X.; Hassan, S.U. Biogenic Nanoparticles: Synthesis, Characterisation and Applications. Appl. Sci. Switz. 2021, 11, 2598. [Google Scholar] [CrossRef]
- Sharma, D.; Kanchi, S.; Bisetty, K. Biogenic Synthesis of Nanoparticles: A Review. Arab. J. Chem. 2019, 12, 3576–3600. [Google Scholar] [CrossRef]
- Loshchinina, E.A.; Vetchinkina, E.P.; Kupryashina, M.A. Diversity of Biogenic Nanoparticles Obtained by the Fungi-Mediated Synthesis: A Review. Biomimetics 2023, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.E.; Escudero, L.V.; Tebes-Cayo, C.; Acosta, M.; Encalada, O.; Fernández-Moroso, S.; Demergasso, C. First Draft Genome Sequence of a Strain from the Genus Fusibacter Isolated from Salar de Ascotán in Northern Chile. Stand. Genomic Sci. 2017, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Grinok, M.; Vázquez, S.; Guiliani, N.; Marín, S.; Demergasso, C. Looking for the Mechanism of Arsenate Respiration of Fusibacter Sp. Strain 3D3, Independent of ArrAB. Front. Microbiol. 2022, 13, 1029886. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, X.; Wang, P.; Wu, Q.; Qi, X.; Han, X.; Chen, L.; Meng, X.; Xu, K. Delivery of Arsenic Trioxide by Multifunction Nanoparticles to Improve the Treatment of Hepatocellular Carcinoma. ACS Appl. Mater. Interfaces 2020, 12, 8016–8029. [Google Scholar] [CrossRef]
- Das, B.; Pal, A.; Pal, R.; Bodo, V.; Newme, D.; Chakraborty, S.; Sengupta, M. Arsenic Nanoparticles Are Effective in Reducing 3-Methylcholanthrene Induced Carcinogenesis in Murine Fibrosarcoma by Promoting Anti-Tumorigenic Inflammation. BioNanoScience 2022, 12, 555–570. [Google Scholar] [CrossRef]
- Wang, T.; Meng, J.; Wang, C.; Wen, T.; Jia, M.; Ge, Y.; Xie, L.; Hao, S.; Liu, J.; Xu, H. Inhibition of Murine Breast Cancer Metastases by Hydrophilic As4S4 Nanoparticles Is Associated with Decreased ROS and HIF-1α Downregulation. Front. Oncol. 2019, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Yu, L.; Wang, X.; Wu, Z.; Zhi, D.; Yang, J.; Guo, Z.; Wu, T.; Sun, Y.; Zhao, L.; et al. A Novel Realgar-Indigo Naturalis Formula More Effectively Induces Apoptosis in NB4 Cells. Pak. J. Pharm. Sci. 2019, 32, 957–962. [Google Scholar]
- Tse, W.P.; Cheng, C.H.K.; Che, C.T.; Lin, Z.X. Arsenic Trioxide, Arsenic Pentoxide, and Arsenic Iodide Inhibit Human Keratinocyte Proliferation through the Induction of Apoptosis. J. Pharmacol. Exp. Ther. 2008, 326, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sridhar, S.; Guo, L.; Lau, S.K.P.; Xu, Y.; Woo, P.C.Y. Successful Treatment of Plantar Warts Using Topical Zijinding, a Traditional Chinese Medicine Preparation: A Case Series. J. Cosmet. Dermatol. 2020, 19, 946–950. [Google Scholar] [CrossRef]
- Zhao, Q.H.; Zhang, Y.; Liu, Y.; Wang, H.L.; Shen, Y.Y.; Yang, W.J.; Wen, L.P. Anticancer Effect of Realgar Nanoparticles on Mouse Melanoma Skin Cancer in Vivo via Transdermal Drug Delivery. Med. Oncol. 2010, 27, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Farooq, I.; Bugshan, A. Oral Squamous Cell Carcinoma: Metastasis, Potentially Associated Malignant Disorders, Etiology and Recent Advancements in Diagnosis. F1000Research 2020, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Neville, B.W.; Day, T.A. Oral Cancer and Precancerous Lesions. CA Cancer J. Clin. 2002, 52, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Demergasso, E.; Serrano, A. Method for Producing Arsenic Sulphide Nanostructures from a Microorganism and Produced Nanostructures. Chile Patent WO2017100959, 27 December 2018. [Google Scholar]
- Debela, D.T.; Muzazu, S.G.Y.; Heraro, K.D.; Ndalama, M.T.; Mesele, B.W.; Haile, D.C.; Kitui, S.K.; Manyazewal, T. New Approaches and Procedures for Cancer Treatment: Current Perspectives. SAGE Open Med. 2021, 9, 1–10. [Google Scholar] [CrossRef]
- Gratton, S.E.A.; Ropp, P.A.; Pohlhaus, P.D.; Luft, J.C.; Madden, V.J.; Napier, M.E.; DeSimone, J.M. The Effect of Particle Design on Cellular Internalization Pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 11613–11618. [Google Scholar] [CrossRef]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13, 339. [Google Scholar] [CrossRef]
- Richards, C.J.; Burgers, T.C.Q.; Vlijm, R.; Roos, W.H.; Åberg, C. Rapid Internalization of Nanoparticles by Human Cells at the Single Particle Level. ACS Nano 2023, 17, 16517–16529. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wang, X.; Xi, R.; Pan, W.; Jiang, S.; Li, Z.; Zhao, Y.; Gao, G.; Liu, D. Enhanced Antitumor Activity of Realgar Mediated by Milling It to Nanosize. Int. J. Nanomed. 2014, 9, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, R.L.R.; De Araújo, T.B.S.; Valverde, L.F.; Silva, V.A.O.; Cavalcante, B.R.R.; Rossi, E.A.; Allahdadi, K.J.; Dos Reis, M.G.; Pereira, T.A.; Coletta, R.D.; et al. Arsenic Trioxide Triggers Apoptosis of Metastatic Oral Squamous Cells Carcinoma with Concomitant Downregulation of GLI1 in Hedgehog Signaling. Biomedicines 2022, 10, 3293. [Google Scholar] [CrossRef] [PubMed]
- Azizi, M.; Ghourchian, H.; Yazdian, F.; Bagherifam, S.; Bekhradnia, S.; Nyström, B. Anti-Cancerous Effect of Albumin Coated Silver Nanoparticles on MDA-MB 231 Human Breast Cancer Cell Line. Sci. Rep. 2017, 7, 5178. [Google Scholar] [CrossRef] [PubMed]
- Platanias, L.C. Biological Responses to Arsenic Compounds. J. Biol. Chem. 2009, 284, 18583–18587. [Google Scholar] [CrossRef] [PubMed]
- Jahangirnejad, R.; Goudarzi, M.; Kalantari, H.; Najafzadeh, H.; Rezaei, M. Subcellular Organelle Toxicity Caused by Arsenic Nanoparticles in Isolated Rat Hepatocytes. Int. J. Occup. Environ. Med. 2020, 11, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Taddei, M.L.; Giannoni, E.; Fiaschi, T.; Chiarugi, P. Anoikis: An Emerging Hallmark in Health and Diseases. J. Pathol. 2012, 226, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis Molecular Pathways and Its Role in Cancer Progression. Biochim. Biophys. Acta-Mol. Cell Res. 2013, 1833, 3481–3498. [Google Scholar] [CrossRef] [PubMed]
- Cholujova, D.; Bujnakova, Z.; Dutkova, E.; Hideshima, T.; Groen, R.W.; Mitsiades, C.S.; Richardson, P.G.; Dorfman, D.M.; Balaz, P.; Anderson, K.C.; et al. Realgar nanoparticles versus ATO arsenic compounds induce in vitro and in vivo activity against multiple myeloma. Br. J. Haematol. 2017, 179, 756–771. [Google Scholar] [CrossRef]
- Han, Y.-H.; Wang, Y.; Lee, S.-J.; Jin, M.-H.; Sun, H.-N.; Kwon, T. Regulation of Anoikis by Extrinsic Death Receptor Pathways. Cell Commun. Signal. 2023, 21, 227. [Google Scholar] [CrossRef]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt Signal Transduction for Cancer Therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.; Mills, G.B. Exploiting the PI3K/AKT Pathway for Cancer Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef] [PubMed]
- Jan, Y.; Matter, M.; Pai, J.T.; Chen, Y.L.; Pilch, J.; Komatsu, M.; Ong, E.; Fukuda, M.; Ruoslahti, E. A Mitochondrial Protein, Bit1, Mediates Apoptosis Regulated by Integrins and Groucho/TLE Corepressors. Cell 2004, 116, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Jenning, S.; Pham, T.; Ireland, S.K.; Ruoslahti, E.; Biliran, H. Bit1 in Anoikis Resistance and Tumor Metastasis. Cancer Lett. 2013, 333, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hu, A.; Chen, H.; Liang, J.; Gu, M.; Xiong, Y.; Mu, C.-F. The Osteogenic Niche-Targeted Arsenic Nanoparticles Prevent Colonization of Disseminated Breast Tumor Cells in the Bone. Acta Pharm. Sin. B 2022, 12, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Hollow, S.E.; Johnstone, T.C. Realgar and Arsenene Nanomaterials as Arsenic-Based Anticancer Agents. Curr. Opin. Chem. Biol. 2023, 72, 102229. [Google Scholar] [CrossRef]
- Yu, X.; Zhu, L. Nanoparticles for the Treatment of Bone Metastasis in Breast Cancer: Recent Advances and Challenges. Int. J. Nanomed. 2024, 19, 1867–1886. [Google Scholar] [CrossRef] [PubMed]
- Teraoka, S.; Kakei, Y.; Akashi, M.; Iwata, E.; Hasegawa, T.; Miyawaki, D.; Sasaki, R.; Komori, T. Gold Nanoparticles Enhance X-ray Irradiation-induced Apoptosis in Head and Neck Squamous Cell Carcinoma In vitro. Biomed. Rep. 2018, 9, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Qin, M.; Chen, X.; Wang, Q.; Zhang, Z.; Sun, X. Combining Photothermal Therapy and Immunotherapy against Melanoma by Polydopamine-Coated Al2O3 Nanoparticles. Theranostics 2018, 8, 2229–2241. [Google Scholar] [CrossRef]
- Wu, J.; Tang, C.; Yin, C. Co-Delivery of Doxorubicin and Interleukin-2 via Chitosan Based Nanoparticles for Enhanced Antitumor Efficacy. Acta Biomater. 2017, 47, 81–90. [Google Scholar] [CrossRef]
- Baharlouei, P.; Rahman, A. Chitin and Chitosan: Prospective Biomedical Applications in Drug Delivery, Cancer Treatment, and Wound Healing. Mar. Drugs 2022, 20, 460. [Google Scholar] [CrossRef]
- Diaz, M.J.; Natarelli, N.; Aflatooni, S.; Aleman, S.J.; Neelam, S.; Tran, J.T.; Taneja, K.; Lucke-Wold, B.; Forouzandeh, M. Nanoparticle-Based Treatment Approaches for Skin Cancer: A Systematic Review. Curr. Oncol. 2023, 30, 7112–7131. [Google Scholar] [CrossRef] [PubMed]
- Ugbaja, R.N.; Ogungbemi, K.; James, A.S.; Peter Folorunsho, A.; Abolade, S.O.; Ajamikoko, S.O.; Atayese, E.O.; Adedeji, O.V. Chitosan from Crabs (Scylla Serrata) Represses Hyperlipidemia-Induced Hepato-Renal Dysfunctions in Rats: Modulation of CD43 and P53 Expression. Pathophysiology 2021, 28, 224–237. [Google Scholar] [CrossRef]
- Hiwatashi, Y.; Tadokoro, H.; Henmi, K.; Arai, M.; Kaise, T.; Tanaka, S.; Hirano, T. Antiproliferative and Anti-Invasive Effects of Inorganic and Organic Arsenic Compounds on Human and Murine Melanoma Cells In Vitro. J. Pharm. Pharmacol. 2011, 63, 1202–1210. [Google Scholar] [CrossRef]
- Aljawish, A.; Chevalot, I.; Jasniewski, J.; Scher, J.; Muniglia, L. Enzymatic Synthesis of Chitosan Derivatives and Their Potential Applications. J. Mol. Catal. B Enzym. 2015, 112, 25–39. [Google Scholar] [CrossRef]
- Pedroso-Santana, S.; Fleitas-Salazar, N.; Sarabia-Sainz, A.; Silva-Campa, E.; Burgara-Estrella, A.; Angulo-Molina, A.; Melendrez, R.; Pedroza-Montero, M.; Riera, R. A Nanodiamond-Fluorescein Conjugate for Cell Studies. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 015013. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2003–2007. [Google Scholar] [CrossRef]
- Chen, P.N.; Lin, C.W.; Yang, S.F.; Chang, Y.C. Oral Submucous Fibrosis Stimulates Invasion and Epithelial-Mesenchymal Transition in Oral Squamous Cell Carcinoma by Activating MMP-2 and IGF-IR. J. Cell. Mol. Med. 2021, 25, 9814–9825. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Covarrubias, A.A.; Reyna-Jeldes, M.; Pedroso-Santana, S.; Marín, S.; Madero-Mendoza, C.; Demergasso, C.; Coddou, C. Arsenic Nanoparticles Trigger Apoptosis via Anoikis Induction in OECM-1 Cells. Int. J. Mol. Sci. 2024, 25, 6723. https://doi.org/10.3390/ijms25126723
Covarrubias AA, Reyna-Jeldes M, Pedroso-Santana S, Marín S, Madero-Mendoza C, Demergasso C, Coddou C. Arsenic Nanoparticles Trigger Apoptosis via Anoikis Induction in OECM-1 Cells. International Journal of Molecular Sciences. 2024; 25(12):6723. https://doi.org/10.3390/ijms25126723
Chicago/Turabian StyleCovarrubias, Alejandra A., Mauricio Reyna-Jeldes, Seidy Pedroso-Santana, Sabrina Marín, Carolina Madero-Mendoza, Cecilia Demergasso, and Claudio Coddou. 2024. "Arsenic Nanoparticles Trigger Apoptosis via Anoikis Induction in OECM-1 Cells" International Journal of Molecular Sciences 25, no. 12: 6723. https://doi.org/10.3390/ijms25126723
APA StyleCovarrubias, A. A., Reyna-Jeldes, M., Pedroso-Santana, S., Marín, S., Madero-Mendoza, C., Demergasso, C., & Coddou, C. (2024). Arsenic Nanoparticles Trigger Apoptosis via Anoikis Induction in OECM-1 Cells. International Journal of Molecular Sciences, 25(12), 6723. https://doi.org/10.3390/ijms25126723