
Unveiling the Secrets of Acinetobacter baumannii: Resistance, Current Treatments, and Future Innovations

 , , , , ,
, , , , ,  ,
, 
Abstract
1. Introduction
2. Resistance Mechanisms
2.1. Enzymatic Inactivation
2.1.1. β-Lactamases
- Types of β-Lactamases Produced by A. baumannii
2.1.2. Aminoglycoside-Modifying Enzymes
2.2. Target Modification
- Target Modification in β-Lactam Resistance
- Target Modification in Aminoglycoside Resistance
- Target modification in colistin resistance
2.3. Reduced Permeability and Active Efflux
- Alterations in Outer Membrane Proteins
- LPS Modifications
- Capsule Production
- Biofilm-Associated Reduced Permeability
- Active Efflux
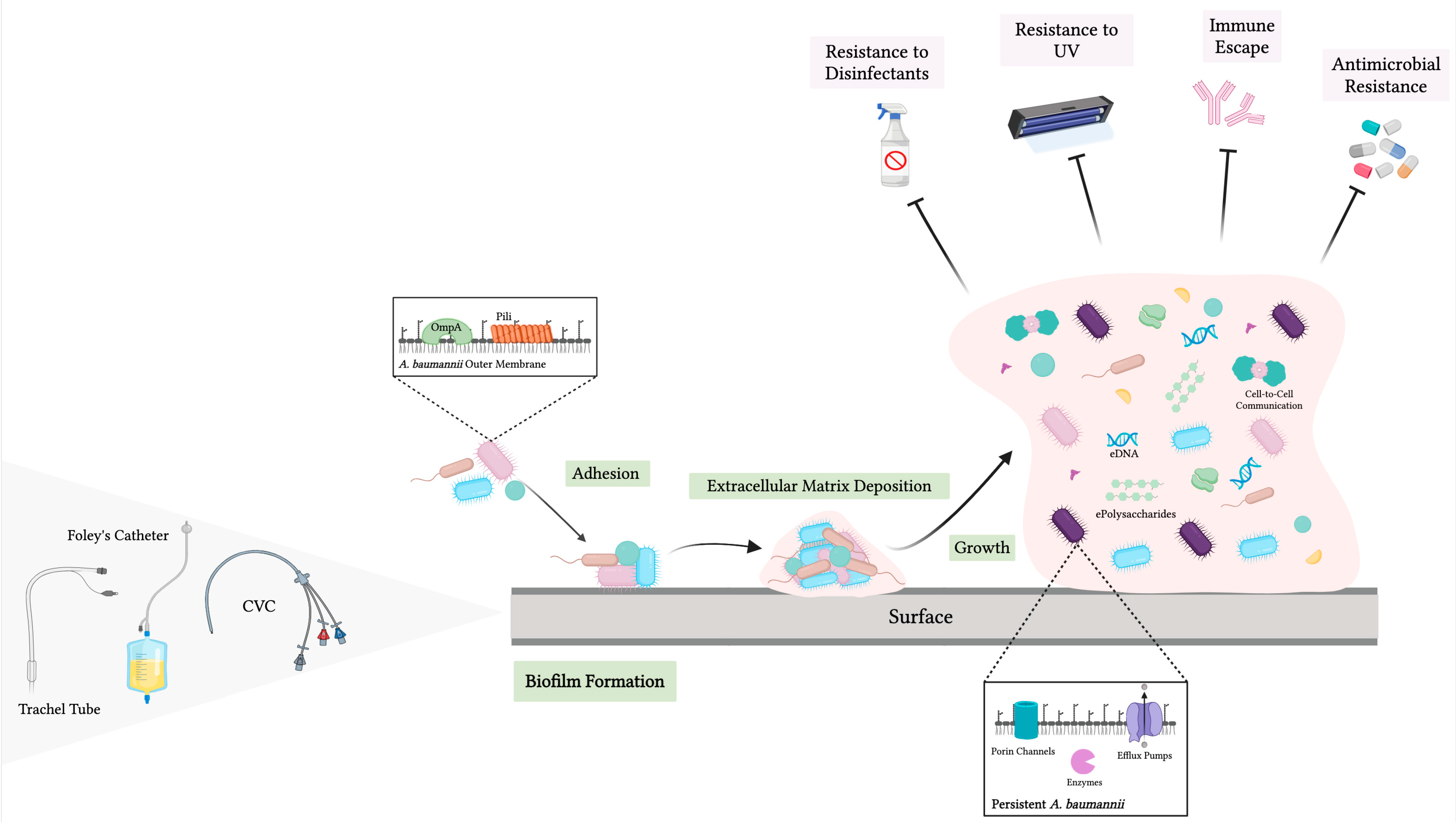
2.4. Biofilm-Associated Resistance
3. Treatment Options
3.1. Current Therapies
3.1.1. Sulbactam-Based Regimens
3.1.2. Tigecycline
3.1.3. Cefiderocol
3.1.4. Eravacycline
3.2. Innovative Approaches
3.2.1. Novel Beta-Lactams/Beta-Lactamase Inhibitor Combinations (BLICs)
Sulbactam/Durlobactam
Cefepime/Zidebactam
Imipenem/Cilastatin/Funobactam
Xeruborbactam
3.2.2. Phage Therapy
3.2.3. Other Strategies
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184. [Google Scholar] [CrossRef] [PubMed]
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 3 May 2024).
- Venkateswaran, P.; Vasudevan, S.; David, H.; Shaktivel, A.; Shanmugam, K.; Neelakantan, P.; Solomon, A.P. Revisiting ESKAPE Pathogens: Virulence, Resistance, and Combating Strategies Focusing on Quorum Sensing. Front. Cell Infect. Microbiol. 2023, 13, 1159798. [Google Scholar] [CrossRef]
- Nasr, P. Genetics, Epidemiology, and Clinical Manifestations of Multidrug-Resistant Acinetobacter baumannii. J. Hosp. Infect. 2020, 104, 4–11. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance Surveillance in Europe 2023—2021 Data; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2023 Guidance on the Treatment of Antimicrobial Resistant Gram-Negative Infections. Clin. Infect. Dis. 2023, ciad428. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.A.; Szabo, D. Mechanisms of Multidrug Resistance in Acinetobacter Species and Pseudomonas aeruginosa. Clin. Infect. Dis. 2006, 43, S49–S56. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, B. Genetic Mechanisms of Antibiotic Resistance and Virulence in Acinetobacter baumannii: Background, Challenges and Future Prospects. Mol. Biol. Rep. 2020, 47, 4037–4046. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Chowdhury, G.; Mukhopadhyay, A.K.; Dutta, S.; Basu, S. Convergence of Biofilm Formation and Antibiotic Resistance in Acinetobacter baumannii Infection. Front. Med. 2022, 9, 793615. [Google Scholar] [CrossRef]
- Waseem, H.; Ali, M.; Waseem, N.; Rashid, M.I.; Ali, A.; Waqas, U. Antimicrobial Resistance Mechanisms in Acinetobacter baumannii in Recent Decade. Saudi J. Pathol. Microbiol. Abbreviated Key Title Saudi J. Pathol. Microbiol. 2021, 6, 179–186. [Google Scholar] [CrossRef]
- Basatian-Tashkan, B.; Niakan, M.; Khaledi, M.; Afkhami, H.; Sameni, F.; Bakhti, S.; Mirnejad, R. Antibiotic Resistance Assessment of Acinetobacter baumannii Isolates from Tehran Hospitals due to the Presence of Efflux Pumps Encoding Genes (AdeA and AdeS Genes) by Molecular Method. BMC Res. Notes 2020, 13, 543. [Google Scholar] [CrossRef]
- Jeon, J.H.; Jang, K.M.; Lee, J.H.; Kang, L.W.; Lee, S.H. Transmission of Antibiotic Resistance Genes through Mobile Genetic Elements in Acinetobacter baumannii and Gene-Transfer Prevention. Sci. Total Environ. 2023, 857, 159497. [Google Scholar] [CrossRef] [PubMed]
- Cain, A.K.; Hamidian, M. Portrait of a Killer: Uncovering Resistance Mechanisms and Global Spread of Acinetobacter baumannii. PLoS Pathog. 2023, 19, e1011520. [Google Scholar] [CrossRef] [PubMed]
- Vrancianu, C.O.; Gheorghe, I.; Czobor, I.B.; Chifiriuc, M.C. Antibiotic Resistance Profiles, Molecular Mechanisms and Innovative Treatment Strategies of Acinetobacter baumannii. Microorganisms 2020, 8, 935. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Mendes, R.E.; Gales, A.C. Global Epidemiology and Mechanisms of Resistance of Acinetobacter baumannii-Calcoaceticus Complex. Clin. Infect. Dis. 2023, 76, S166–S178. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-J.; Xiao, Z.-G.; Lv, X.-J.; Huang, H.-T.; Liao, C.; Hui, C.-Y.; Xu, Y.; Li, H.-F. Drug-resistant Acinetobacter baumannii: From Molecular Mechanisms to Potential Therapeutics (Review). Exp. Ther. Med. 2023, 25, 209. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, X.; Wang, Y.; Tao, Y.; Shao, X.; Li, Y.; Li, W. Insight into Carbapenem Resistance and Virulence of Acinetobacter baumannii from a Children’s Medical Centre in Eastern China. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 47. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef]
- Lupo, A.; Haenni, M.; Madec, J.-Y. Antimicrobial Resistance in Acinetobacter Spp. and Pseudomonas Spp. Microbiol Spectr. 2018, 6, 377–393. [Google Scholar] [CrossRef]
- Cafiso, V.; Stracquadanio, S.; Lo Verde, F.; Gabriele, G.; Mezzatesta, M.L.; Caio, C.; Pigola, G.; Ferro, A.; Stefani, S. Colistin Resistant A. baumannii: Genomic and Transcriptomic Traits Acquired under Colistin Therapy. Front. Microbiol. 2019, 9, 3195. [Google Scholar] [CrossRef]
- Novović, K.; Jovčić, B. Colistin Resistance in Acinetobacter baumannii: Molecular Mechanisms and Epidemiology. Antibiotics 2023, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Thangaraj, P.; Chakrabarti, A. Acinetobacter baumannii: A Brief Account of Mechanisms of Multidrug Resistance and Current and Future Therapeutic Management. J. Clin. Diagn. Res. 2013, 7, 2602. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-Nodulation-Cell Division-Type Efflux Pump Involved in Aminoglycoside Resistance in Acinetobacter baumannii Strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.R.; Casella, L.G.; Jones, J.W.; Adams, M.D.; Zurawski, D.V.; Hazlett, K.R.O.; Doi, Y.; Ernst, R.K. Unique Structural Modifications Are Present in the Lipopolysaccharide from Colistin-Resistant Strains of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Akoolo, L.; Pires, S.; Kim, J.; Parker, D. The Capsule of Acinetobacter baumannii Protects against the Innate Immune Response. J. Innate Immun. 2022, 14, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Gedefie, A.; Demsis, W.; Ashagrie, M.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 14, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Nadeem, A.; Mushtaq, F.; Zlatkov, N.; Shahzad, M.; Zavialov, A.V.; Wai, S.N.; Uhlin, B.E. Csu Pili Dependent Biofilm Formation and Virulence of Acinetobacter baumannii. Npj Biofilms Microbiomes 2023, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.G.; Combo, S.I.; Allain, T.; Domingues, S.; Buret, A.G.; Da Silva, G.J. Co-Regulation of Biofilm Formation and Antimicrobial Resistance in Acinetobacter baumannii: From Mechanisms to Therapeutic Strategies. Eur. J. Clin. Microbiol. Infect. Dis. 2023, 42, 1405–1423. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.; Shrivastava, R.; Vashistt, J. Acinetobacter baumannii Biofilm Formation: Association with Antimicrobial Resistance and Prolonged Survival under Desiccation. Curr. Microbiol. 2022, 79, 361. [Google Scholar] [CrossRef]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm Formation in Acinetobacter baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef]
- Lysitsas, M.; Triantafillou, E.; Chatzipanagiotidou, I.; Antoniou, K.; Spyrou, V.; Billinis, C.; Valiakos, G. Phenotypic Investigation and Detection of Biofilm-Associated Genes in Acinetobacter baumannii Isolates, Obtained from Companion Animals. Trop. Med. Infect. Dis. 2024, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Muras, A.; Parga, A.; Romero, M.; Rumbo-Feal, S.; Poza, M.; Ramos-Vivas, J.; Otero, A. Quorum Sensing as a Target for Controlling Surface Associated Motility and Biofilm Formation in Acinetobacter baumannii ATCC® 17978TM. Front. Microbiol. 2020, 11, 565548. [Google Scholar] [CrossRef]
- Subhadra, B.; Oh, M.H.; Choi, C.H.; Subhadra, B.; Oh, M.H.; Choi, C.H. Quorum Sensing in Acinetobacter: With Special Emphasis on Antibiotic Resistance, Biofilm Formation and Quorum Quenching. AIMS Microbiol. 2016, 2, 27–41. [Google Scholar] [CrossRef]
- Sun, X.; Ni, Z.; Tang, J.; Ding, Y.; Wang, X.; Li, F. The AbaI/AbaR Quorum Sensing System Effects on Pathogenicity in Acinetobacter baumannii. Front. Microbiol. 2021, 12, 679241. [Google Scholar] [CrossRef]
- Ng, C.K.; How, K.Y.; Tee, K.K.; Chan, K.G. Characterization and Transcriptome Studies of Autoinducer Synthase Gene from Multidrug Resistant Acinetobacter baumannii Strain 863. Genes 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, J.M.; Rather, P.N. Insights Into Mechanisms of Biofilm Formation in Acinetobacter baumannii and Implications for Uropathogenesis. Front. Cell Infect. Microbiol. 2020, 10, 544784. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Martí, S.; Soto, S.; Fernández-Cuenca, F.; Cisneros, J.M.; Pachón, J.; Pascual, A.; Martínez-Martínez, L.; Mcqueary, C.; Actis, L.A.; et al. Biofilm Formation in Acinetobacter baumannii: Associated Features and Clinical Implications. Clin. Microbiol. Infect. 2008, 14, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii Biofilm Formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii Biofilm-Associated Protein Plays a Role in Adherence to Human Epithelial Cells. Infect. Immun. 2012, 80, 228–233. [Google Scholar] [CrossRef]
- Mayer, C.; Muras, A.; Romero, M.; López, M.; Tomás, M.; Otero, A. Multiple Quorum Quenching Enzymes Are Active in the Nosocomial Pathogen Acinetobacter baumannii ATCC17978. Front. Cell Infect. Microbiol. 2018, 8, 310. [Google Scholar] [CrossRef]
- Penwell, W.F.; Shapiro, A.B.; Giacobbe, R.A.; Gu, R.F.; Gao, N.; Thresher, J.; McLaughlin, R.E.; Huband, M.D.; DeJonge, B.L.M.; Ehmann, D.E.; et al. Molecular Mechanisms of Sulbactam Antibacterial Activity and Resistance Determinants in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.; Tanudra, A.; Chen, A.; Miller, A.A.; McLeod, S.M.; Tommasi, R. In Vitro Pharmacokinetics/Pharmacodynamics of the β-Lactamase Inhibitor, Durlobactam, in Combination with Sulbactam against Acinetobacter baumannii-Calcoaceticus Complex. Antimicrob. Agents Chemother. 2024, 68, e00312-23. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Paterson, D.L.; Tamma, P.D. Navigating Available Treatment Options for Carbapenem-Resistant Acinetobacter baumannii-Calcoaceticus Complex Infections. Clin. Infect. Dis. 2023, 76, S179–S193. [Google Scholar] [CrossRef] [PubMed]
- Wood, G.C.; Hanes, S.O.; Croce, M.A.; Fabian, T.C.; Boucher, B.A. Comparison of Ampicillin-Sulbactam and Imipenem-Cilastatin for the Treatment of Acinetobacter Ventilator-Associated Pneumonia. Clin. Infect. Dis. 2002, 34, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Principe, L.; Di Bella, S.; Conti, J.; Perilli, M.; Piccirilli, A.; Mussini, C.; Decorti, G. Acinetobacter baumannii Resistance to Sulbactam/Durlobactam: A Systematic Review. Antibiotics 2022, 11, 1793. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Xu, P.; Zhu, Y.; Wang, K.; Xiang, D.; Wang, F.; Banh, H.L. Clinical Experience with Tigecycline in the Treatment of Hospital-Acquired Pneumonia Caused by Multidrug Resistant Acinetobacter baumannii. BMC Pharmacol. Toxicol. 2019, 20, 19. [Google Scholar] [CrossRef]
- Karageorgopoulos, D.E.; Kelesidis, T.; Kelesidis, I.; Falagas, M.E. Tigecycline for the Treatment of Multidrug-Resistant (Including Carbapenem-Resistant) Acinetobacter Infections: A Review of the Scientific Evidence. J. Antimicrob. Chemother. 2008, 62, 45–55. [Google Scholar] [CrossRef]
- Lee, Y.T.; Tsao, S.M.; Hsueh, P.R. Clinical Outcomes of Tigecycline Alone or in Combination with Other Antimicrobial Agents for the Treatment of Patients with Healthcare-Associated Multidrug-Resistant Acinetobacter baumannii Infections. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1211–1220. [Google Scholar] [CrossRef]
- Sun, C.; Yu, Y.; Hua, X. Resistance Mechanisms of Tigecycline in Acinetobacter baumannii. Front. Cell Infect. Microbiol. 2023, 13, 1141490. [Google Scholar] [CrossRef]
- Liu, C.; Liu, J.; Lu, Q.; Wang, P.; Zou, Q. The Mechanism of Tigecycline Resistance in Acinetobacter baumannii under Sub-Minimal Inhibitory Concentrations of Tigecycline. Int. J. Mol. Sci. 2024, 25, 1819. [Google Scholar] [CrossRef]
- Stracquadanio, S.; Bonomo, C.; Marino, A.; Bongiorno, D.; Privitera, G.F.; Bivona, D.A.; Mirabile, A.; Bonacci, P.G.; Stefani, S. Acinetobacter baumannii and Cefiderocol, between Cidality and Adaptability. Microbiol. Spectr. 2022, 10, e0234722. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Stracquadanio, S.; Campanella, E.; Munafò, A.; Gussio, M.; Ceccarelli, M.; Bernardini, R.; Nunnari, G.; Cacopardo, B. Intravenous Fosfomycin: A Potential Good Partner for Cefiderocol. Clinical Experience and Considerations. Antibiotics 2023, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Echols, R.; Matsunaga, Y.; Ariyasu, M.; Doi, Y.; Ferrer, R.; Lodise, T.P.; Naas, T.; Niki, Y.; Paterson, D.L.; et al. Efficacy and Safety of Cefiderocol or Best Available Therapy for the Treatment of Serious Infections Caused by Carbapenem-Resistant Gram-Negative Bacteria (CREDIBLE-CR): A Randomised, Open-Label, Multicentre, Pathogen-Focused, Descriptive, Phase 3 Trial. Lancet Infect. Dis. 2021, 21, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, C.H.; Kaas, A.; Rasigade, J.P.; Aubrun, F.; Argaud, L.; Balanca, B.; Fellahi, J.L.; Richard, J.C.; Lukaszewicz, A.C.; Wallet, F.; et al. Cefiderocol in Difficult-to-Treat Nf-GNB in ICU Settings. Ann. Intensive Care 2024, 14, 73. [Google Scholar] [CrossRef] [PubMed]
- Viale, P.; Sandrock, C.E.; Ramirez, P.; Rossolini, G.M.; Lodise, T.P. Treatment of Critically Ill Patients with Cefiderocol for Infections Caused by Multidrug-Resistant Pathogens: Review of the Evidence. Ann. Intensive Care 2023, 13, 52. [Google Scholar] [CrossRef]
- Onorato, L.; de Luca, I.; Monari, C.; Coppola, N. Cefiderocol Either in Monotherapy or Combination versus Best Available Therapy in the Treatment of Carbapenem-Resistant Acinetobacter baumannii Infections: A Systematic Review and Meta-Analysis. J. Infect. 2024, 88, 106113. [Google Scholar] [CrossRef] [PubMed]
- Smoke, S.M.; Brophy, A.; Reveron, S.; Iovleva, A.; Kline, E.G.; Marano, M.; Miller, L.P.; Shields, R.K. Evolution and Transmission of Cefiderocol-Resistant Acinetobacter baumannii during an Outbreak in the Burn Intensive Care Unit. Clin. Infect. Dis. 2023, 76, e1261–e1265. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Arias, C.A. ESKAPE Pathogens: Antimicrobial Resistance, Epidemiology, Clinical Impact and Therapeutics. Nat. Rev. Microbiol. 2024, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Stefan, M.; Seifert, H.; Müller, C.; Stefanik, D.; Higgins, P.G.; Wohlfarth, E.; Kresken, M. In Vitro Activity of Cefiderocol against a Global Collection of Carbapenem-Resistant Acinetobacter baumannii Isolates. Antibiotics 2023, 12, 1172. [Google Scholar] [CrossRef]
- Mezcord, V.; Escalante, J.; Nishimura, B.; Traglia, G.M.; Sharma, R.; Vallé, Q.; Tuttobene, M.R.; Subils, T.; Marin, I.; Pasteran, F.; et al. Induced Heteroresistance in Carbapenem-Resistant Acinetobacter baumannii (CRAB) via Exposure to Human Pleural Fluid (HPF) and Its Impact on Cefiderocol Susceptibility. Int. J. Mol. Sci. 2023, 24, 11752. [Google Scholar] [CrossRef]
- Stracquadanio, S.; Nicolosi, A.; Privitera, G.F.; Massimino, M.; Marino, A.; Bongiorno, D.; Stefani, S. Role of Transcriptomic and Genomic Analyses in Improving the Comprehension of Cefiderocol Activity in Acinetobacter baumannii. mSphere 2024, 9, e00617-23. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.; Bianco, G.; Boattini, M.; Nordmann, P. In Vivo Development of Cefiderocol Resistance in Carbapenem-Resistant Acinetobacter baumannii Associated with the Downregulation of a TonB-Dependent Siderophore Receptor, PiuA. J. Antimicrob. Chemother. 2024, 79, 928–930. [Google Scholar] [CrossRef] [PubMed]
- Eucast: Cefiderocol Susceptibility Testing—An Update. Available online: https://www.eucast.org/eucast_news/news_singleview?tx_ttnews%5Btt_news%5D=554&cHash=46fef43d5bf84991a0af2d658deaf685 (accessed on 1 June 2024).
- Gatti, M.; Cosentino, F.; Giannella, M.; Viale, P.; Pea, F. In Reply to the Letter to the Editor Regarding “Clinical Efficacy of Cefiderocol-Based Regimens in Patients with Carbapenem-Resistant Acinetobacter baumannii Infections: A Systematic Review with Meta-Analysis”. Int. J. Antimicrob. Agents 2024, 63, 107168. [Google Scholar] [CrossRef] [PubMed]
- Serapide, F.; Guastalegname, M.; Gullì, S.P.; Lionello, R.; Bruni, A.; Garofalo, E.; Longhini, F.; Trecarichi, E.M.; Russo, A. Antibiotic Treatment of Carbapenem-Resistant Acinetobacter baumannii Infections in View of the Newly Developed β-Lactams: A Narrative Review of the Existing Evidence. Antibiotics 2024, 13, 506. [Google Scholar] [CrossRef]
- Scott, L.J. Eravacycline: A Review in Complicated Intra-Abdominal Infections. Drugs 2019, 79, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Letizia Barreca, M.; Sabatini, S.; Astolfi, A.; Russo, A.; Brauncajs, M.; Bielec, F.; Macieja, A.; Pastuszak-Lewandoska, D. In Vitro Activity of Eravacycline against Carbapenemase-Producing Gram-Negative Bacilli Clinical Isolates in Central Poland. Biomedicines 2023, 11, 1784. [Google Scholar] [CrossRef] [PubMed]
- Grossman, T.H.; Starosta, A.L.; Fyfe, C.; O’Brien, W.; Rothstein, D.M.; Mikolajka, A.; Wilson, D.N.; Sutcliffe, J.A. Target- and Resistance-Based Mechanistic Studies with TP-434, a Novel Fluorocycline Antibiotic. Antimicrob. Agents Chemother. 2012, 56, 2559–2564. [Google Scholar] [CrossRef] [PubMed]
- Solomkin, J.; Evans, D.; Slepavicius, A.; Lee, P.; Marsh, A.; Tsai, L.; Sutcliffe, J.A.; Horn, P. Assessing the Efficacy and Safety of Eravacycline vs. Ertapenem in Complicated Intra-Abdominal Infections in the Investigating Gram-Negative Infections Treated With Eravacycline (IGNITE 1) Trial: A Randomized Clinical Trial. JAMA Surg. 2017, 152, 224–232. [Google Scholar] [CrossRef]
- Solomkin, J.S.; Gardovskis, J.; Lawrence, K.; Montravers, P.; Sway, A.; Evans, D.; Tsai, L. IGNITE4: Results of a Phase 3, Randomized, Multicenter, Prospective Trial of Eravacycline vs. Meropenem in the Treatment of Complicated Intraabdominal Infections. Clin. Infect. Dis. 2019, 69, 921–929. [Google Scholar] [CrossRef]
- Newman, J.; Izmailyan, S.; Fyfe, C.; Tsai, L. 1963. Combined Microbiological Response Rates from Two Phase 3 Trials Demonstrating the Activity of Eravacycline in the Treatment of Complicated Intra-Abdominal Infections: A Pooled Analysis of IGNITE1 and IGNITE4. Open Forum Infect. Dis. 2018, 5, S568. [Google Scholar] [CrossRef]
- Kunz Coyne, A.J.; Alosaimy, S.; Lucas, K.; Lagnf, A.M.; Morrisette, T.; Molina, K.C.; DeKerlegand, A.; Schrack, M.R.; Kang-Birken, S.L.; Hobbs, A.L.V.; et al. Eravacycline, the First Four Years: Health Outcomes and Tolerability Data for 19 Hospitals in 5 U.S. Regions from 2018 to 2022. Microbiol. Spectr. 2024, 12, e0235123. [Google Scholar] [CrossRef]
- European Society of Clinical Microbiology and Infectious Diseases EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 9 May 2022).
- Clinical & Laboratory Standards Institute: CLSI Guidelines. Available online: https://clsi.org/ (accessed on 17 June 2024).
- Petropoulou, D.; Siopi, M.; Vourli, S.; Pournaras, S. Activity of Sulbactam-Durlobactam and Comparators Against a National Collection of Carbapenem-Resistant Acinetobacter baumannii Isolates from Greece. Front. Cell Infect. Microbiol. 2022, 11, 814530. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.P.; Bhavnani, S.M. The Pharmacokinetics/Pharmacodynamic Relationship of Durlobactam in Combination with Sulbactam in In Vitro and In Vivo Infection Model Systems Versus Acinetobacter baumannii-Calcoaceticus Complex. Clin. Infect. Dis. 2023, 76, S202. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.; Tanudra, A.; Chen, A.; Newman, J.; McLeod, S.M.; Tommasi, R. In Vivo Dose Response and Efficacy of the β-Lactamase Inhibitor, Durlobactam, in Combination with Sulbactam against the Acinetobacter baumannii-Calcoaceticus Complex. Antimicrob. Agents Chemother. 2024, 68, e0080023. [Google Scholar] [CrossRef]
- Arer, V.; Kar, D. Biochemical Exploration of β-Lactamase Inhibitors. Front. Genet. 2023, 13, 1060736. [Google Scholar] [CrossRef] [PubMed]
- Luci, G.; Mattioli, F.; Falcone, M.; Di Paolo, A. Pharmacokinetics of Non-β-Lactam β-Lactamase Inhibitors. Antibiotics 2021, 10, 769. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.S.; Shorr, A.F.; Wunderink, R.G.; Du, B.; Poirier, G.E.; Rana, K.; Miller, A.; Lewis, D.; O’Donnell, J.; Chen, L.; et al. Efficacy and Safety of Sulbactam-Durlobactam versus Colistin for the Treatment of Patients with Serious Infections Caused by Acinetobacter baumannii-Calcoaceticus Complex: A Multicentre, Randomised, Active-Controlled, Phase 3, Non-Inferiority Clinical Trial (ATTACK). Lancet Infect. Dis. 2023, 23, 1072–1084. [Google Scholar] [CrossRef]
- Sagan, O.; Yakubsevitch, R.; Yanev, K.; Fomkin, R.; Stone, E.; Hines, D.; O’Donnell, J.; Miller, A.; Isaacs, R.; Srinivasan, S. Pharmacokinetics and Tolerability of Intravenous Sulbactam-Durlobactam with Imipenem-Cilastatin in Hospitalized Adults with Complicated Urinary Tract Infections, Including Acute Pyelonephritis. Antimicrob. Agents Chemother. 2020, 64, e01506-19. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M.; Mushtaq, S.; Warner, M.; Vickers, A.; Woodford, N. In Vitro Activity of Cefepime/Zidebactam (WCK 5222) against Gram-Negative Bacteria. J. Antimicrob. Chemother. 2017, 72, 1373–1385. [Google Scholar] [CrossRef]
- Bassetti, M.; Castaldo, N.; Fantin, A.; Giacobbe, D.R.; Vena, A. Antibiotic Therapy for Nonfermenting Gram-Negative Bacilli Infections: Future Perspectives. Curr. Opin. Infect. Dis. 2023, 36, 615–622. [Google Scholar] [CrossRef]
- Liu, P.Y.; Ko, W.C.; Lee, W.S.; Lu, P.L.; Chen, Y.H.; Cheng, S.H.; Lu, M.C.; Lin, C.Y.; Wu, T.S.; Yen, M.Y.; et al. In Vitro Activity of Cefiderocol, Cefepime/Enmetazobactam, Cefepime/Zidebactam, Eravacycline, Omadacycline, and Other Comparative Agents against Carbapenem-Non-Susceptible Pseudomonas Aeruginosa and Acinetobacter baumannii Isolates Associated from Bloodstream Infection in Taiwan between 2018–2020. J. Microbiol. Immunol. Infect. 2022, 55, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, S.S.; Periasamy, H.; Takalkar, S.S.; Palwe, S.R.; Khande, H.N.; Patel, M.V. The Novel β-Lactam Enhancer Zidebactam Augments the In Vivo Pharmacodynamic Activity of Cefepime in a Neutropenic Mouse Lung Acinetobacter baumannii Infection Model. Antimicrob. Agents Chemother. 2019, 63, e02146-18. [Google Scholar] [CrossRef] [PubMed]
- Le Terrier, C.; Freire, S.; Nordmann, P.; Poirel, L. Multidrug-Resistant Gram-Negative Clinical Isolates with Reduced Susceptibility/Resistance to Cefiderocol: Which Are the Best Present and Future Therapeutic Alternatives? Eur. J. Clin. Microbiol. Infect. Dis. 2024, 43, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Bhavsar, S.; Joshi, S.; Pawar, S.; Pavase, L.; Mishra, A.; Jadhav, S.; Dabhade, S.; Kayastha, A.K.; Yeole, R.; Deshpande, P.; et al. Structure Activity Relationship (SAR) Driven Design and Discovery of WCK 5107 (Zidebactam): Novel β-Lactam Enhancer, Potent against Multidrug-Resistant Gram-Negative Pathogens. Med. Chem. Res. 2023, 32, 2245–2255. [Google Scholar] [CrossRef]
- Guo, Y.; Han, R.; Jiang, B.; Ding, L.; Yang, F.; Zheng, B.; Yang, Y.; Wu, S.; Yin, D.; Zhu, D.; et al. In Vitro Activity of New β-Lactam-β-Lactamase Inhibitor Combinations and Comparators against Clinical Isolates of Gram-Negative Bacilli: Results from the China Antimicrobial Surveillance Network (CHINET) in 2019. Microbiol. Spectr. 2022, 10, e0185422. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.S.; Abdelghani, S.; Snyder, J.W.; Thomson, G.K. Activity of Cefepime-Zidebactam against Multidrug-Resistant (MDR) Gram-Negative Pathogens. Antibiotics 2019, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Almarzoky Abuhussain, S.S.; Avery, L.M.; Abdelraouf, K.; Nicolaua, D.P. In Vivo Efficacy of Humanized WCK 5222 (Cefepime-Zidebactam) Exposures against Carbapenem-Resistant Acinetobacter baumannii in the Neutropenic Thigh Model. Antimicrob Agents Chemother 2019, 63, e01931-18. [Google Scholar] [CrossRef] [PubMed]
- Study Details|MED Study to Evaluate the Safety, Tolerability and Pharmacokinetics of Intravenous WCK 5222 (Zidebactam and Cefepime) in Healthy Volunteers|ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/study/NCT02707107 (accessed on 28 May 2024).
- Study Details|to Investigate The Pharmacokinetics of Intravenous WCK 5222 (FEP-ZID) in Patients with Renal Impairment|ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/study/NCT02942810 (accessed on 28 May 2024).
- Plasma and Intrapulmonary Concentrations Study of WCK 5222—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT03630094 (accessed on 28 May 2024).
- Evaluate the Effect of WCK 5222 on the QT/QTc Interval in Healthy Volunteers—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT03554304 (accessed on 28 May 2024).
- Study of Cefepime-Zidebactam (FEP-ZID) in Complicated Urinary Tract Infection (CUTI) or Acute Pyelonephritis (AP)—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04979806 (accessed on 28 May 2024).
- Li, Y.; Yan, M.; Xue, F.; Zhong, W.; Liu, X.; Chen, X.; Wu, Y.; Zhang, J.; Wang, Q.; Zheng, B.; et al. In Vitro and in Vivo Activities of a Novel β-Lactamase Inhibitor Combination Imipenem/XNW4107 against Recent Clinical Gram-Negative Bacilli from China. J. Glob. Antimicrob. Resist. 2022, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fratoni, A.J.; Berry, A.V.; Liu, X.; Chen, X.; Wu, Y.; Nicolau, D.P.; Abdelraouf, K. Imipenem/Funobactam (Formerly XNW4107) in Vivo Pharmacodynamics against Serine Carbapenemase-Producing Gram-Negative Bacteria: A Novel Modelling Approach for Time-Dependent Killing. J. Antimicrob. Chemother. 2023, 78, 2343–2353. [Google Scholar] [CrossRef]
- Study Details|Evaluation of the Efficacy and Safety of Intravenous Imipenem/Cilastatin/XNW4107 in Comparison with Meropenem in Hospitalized Adults with CUTI Including AP (EudraCT No. 2022-000061-40)|ClinicalTrials.Gov. Available online: https://www.clinicaltrials.gov/study/NCT05204368?term=XNW4107&rank=6 (accessed on 16 June 2024).
- Study Details|Imipenem/Cilastatin-XNW4107 versus Imipenem/Cilastatin/Relebactam for Treatment of Participants with Bacterial Pneumonia (XNW4107-302, REITAB-2)|ClinicalTrials.Gov. Available online: https://www.clinicaltrials.gov/study/NCT05204563?term=XNW4107&limit=50&page=1&rank=5 (accessed on 16 June 2024).
- Arya, R.; Goldner, B.S.; Shorr, A.F. Novel Agents in Development for Multidrug-Resistant Gram-Negative Infections: Potential New Options Facing Multiple Challenges. Curr. Opin. Infect. Dis. 2022, 35, 589–594. [Google Scholar] [CrossRef]
- Sun, D.; Tsivkovski, R.; Pogliano, J.; Tsunemoto, H.; Nelson, K.; Rubio-Aparicio, D.; Lomovskaya, O. Intrinsic Antibacterial Activity of Xeruborbactam In Vitro: Assessing Spectrum and Mode of Action. Antimicrob. Agents Chemother. 2022, 66, e0087922. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.; Rubio-Aparicio, D.; Sun, D.; Dudley, M.; Lomovskaya, O. In Vitro Activity of the Ultrabroad-Spectrum-Beta-Lactamase Inhibitor QPX7728 against Carbapenem-Resistant Enterobacterales with Varying Intrinsic and Acquired Resistance Mechanisms. Antimicrob. Agents Chemother. 2020, 64, e00757-20. [Google Scholar] [CrossRef] [PubMed]
- P1 Single and Multiple Ascending Dose (SAD/MAD) Study of IV QPX7728 Alone and Combined with QPX2014 in NHV—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04380207 (accessed on 28 May 2024).
- Phase 1 Oral QPX7831 SAD and MAD in Healthy Adults—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04578873 (accessed on 28 May 2024).
- Griffith, D.; Roberts, J.; Wallis, S.; Hernandez-Mitre, M.P.; Morgan, E.; Gehrke, S.; Dudley, M.; Loutit, J. 216. A Phase 1 Study of the Safety, Tolerability, and Pharmacokinetics of Multiple Doses of the Beta-Lactamase Inhibitor Xeruborbactam Alone and in Combination Meropenem in Healthy Adult Subjects. Open Forum Infect. Dis. 2022, 9, ofac492.294. [Google Scholar] [CrossRef]
- Tarazi, Z.; Roos, N.; Page, T.; Griffith, D. 615. Pharmacodynamics (PD) of the Beta-Lactamase Inhibitor Xeruborbactam When Administered in Combination with Meropenem. Open Forum Infect. Dis. 2022, 9, ofac492.667. [Google Scholar] [CrossRef]
- Griffith, D.; Roberts, J.; Wallis, S.; Hernandez-Mitre, M.P.; Morgan, E.; Dudley, M.; Loutit, J. 218. A Phase 1 Study of the Single-Dose Safety, Tolerability, and Pharmacokinetics of the Beta-Lactamase Inhibitor Xeruborbactam Administered as the Isobutyryloxymethyl Oral Prodrug to Healthy Adult Subjects. Open Forum Infect. Dis. 2022, 9, ofac492.296. [Google Scholar] [CrossRef]
- Drug-Drug Interaction Study of IV QPX2014 Combined with QPX7728 in Healthy Adult Subjects—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05072444 (accessed on 28 May 2024).
- P1, DDI & MAD PK and Safety Study of Xeruborbactam Oral Prodrug in Combo with Ceftibuten in Healthy Participants—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT06079775 (accessed on 28 May 2024).
- PK & Safety Study of Xeruborbactam Oral Prodrug Combined with Ceftibuten in Participants with Renal Impairment—Full Text View—ClinicalTrials.Gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT06157242 (accessed on 28 May 2024).
- Tu, Q.; Pu, M.; Li, Y.; Wang, Y.; Li, M.; Song, L.; Li, M.; An, X.; Fan, H.; Tong, Y. Acinetobacter baumannii Phages: Past, Present and Future. Viruses 2023, 15, 673. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Luo, T.; Yang, Y.; Dong, D.; Wang, R.; Wang, Y.; Xu, M.; Guo, X.; Hu, F.; He, P. Phage Therapy as a Promising New Treatment for Lung Infection Caused by Carbapenem-Resistant Acinetobacter baumannii in Mice. Front. Microbiol. 2018, 8, 318476. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, Y.; Galgano, S.; Houdijk, J.; Xie, W.; Jin, Y.; Lin, J.; Song, W.; Fu, Y.; Li, X.; et al. Recent Progress in Phage Therapy to Modulate Multidrug-Resistant Acinetobacter baumannii, Including in Human and Poultry. Antibiotics 2022, 11, 1406. [Google Scholar] [CrossRef]
- Eccleston, A. Macrocyclic Peptides Thwart Gram-Negative Bacteria. Nat. Rev. Drug Discov. 2024, 23, 171. [Google Scholar] [CrossRef]
- Bagcchi, S. New Candidate to Combat Drug-Resistant Bacteria. Lancet Infect. Dis. 2024, 24, e227. [Google Scholar] [CrossRef]
- Hawser, S.; Kothari, N.; Valmont, T.; Louvel, S.; Zampaloni, C. 2131. Activity of the Novel Antibiotic Zosurabalpin (RG6006) against Clinical Acinetobacter Isolates from China. Open Forum Infect Dis 2023, 10, ofad500.1754. [Google Scholar] [CrossRef]
- Gugger, M.K.; Hergenrother, P.J. A New Type of Antibiotic Targets a Drug-Resistant Bacterium. Nature 2024, 625, 451–452. [Google Scholar] [CrossRef]
- Huband, M.D.; Thompson, J.D.; Gurung, N.D.; Liu, Q.; Li, L.; Zhang, J.; Streit, J.M.; Castanheira, M. Activity of the Novel Aminomethylcycline KBP-7072 and Comparators against 1,057 Geographically Diverse Recent Clinical Isolates from the SENTRY Surveillance Program, 2019. Antimicrob. Agents Chemother. 2022, 66, e0139721. [Google Scholar] [CrossRef]
- Juhas, M.; Widlake, E.; Teo, J.; Huseby, D.L.; Tyrrell, J.M.; Polikanov, Y.S.; Ercan, O.; Petersson, A.; Cao, S.; Aboklaish, A.F.; et al. In Vitro Activity of Apramycin against Multidrug-, Carbapenem- and Aminoglycoside-Resistant Enterobacteriaceae and Acinetobacter baumannii. J. Antimicrob. Chemother. 2019, 74, 944–952. [Google Scholar] [CrossRef]
- Becker, K.; Aranzana-Climent, V.; Cao, S.; Nilsson, A.; Shariatgorji, R.; Haldimann, K.; Platzack, B.; Hughes, D.; Andrén, P.E.; Böttger, E.C.; et al. Efficacy of EBL-1003 (Apramycin) against Acinetobacter baumannii Lung Infections in Mice. Clin. Microbiol. Infect. 2021, 27, 1315–1321. [Google Scholar] [CrossRef]
- Li, L.; Tan, X.; Zhou, T.; Chi, S.; Zhu, Y.; Liu, Q.; Chen, Y.; Zhang, J. In Vivo Efficacy and PK/PD Analyses of Zifanocycline (KBP-7072), an Aminomethylcycline Antibiotic, against Acinetobacter baumannii in a Neutropenic Murine Thigh Infection Model. J. Infect. Chemother. 2024, 30, 34–39. [Google Scholar] [CrossRef]
- Mendes, R.E.; Sader, H.S.; Ryan Arends, S.J.; Cotroneo, N.; Critchley, I.A.; Critchley, I.A.; Castanheira, M. 1676. Activity of SPR206 and Comparator Agents against Pseudomonas Aeruginosa and Acinetobacter baumannii Causing Infections in United States Hospitals. Open Forum Infect. Dis. 2022, 9, ofac492.1306. [Google Scholar] [CrossRef]
- Castanheira, M.; Lindley, J.; Huynh, H.; Mendes, R.E.; Lomovskaya, O. 690. Activity of a Novel Polymyxin Analog, QPX9003, Tested against Resistant Gram-Negative Pathogens, Including Carbapenem-Resistant Acinetobacter, Enterobacterales, and Pseudomonas. Open Forum Infect. Dis. 2019, 6, S313. [Google Scholar] [CrossRef]
- Lepak, A.J.; Wang, W.; Andes, D.R. Pharmacodynamic Evaluation of MRX-8, a Novel Polymyxin, in the Neutropenic Mouse Thigh and Lung Infection Models against Gram-Negative Pathogens. Antimicrob. Agents Chemother. 2020, 64, e01517-20. [Google Scholar] [CrossRef]
- Li, X.; Ma, S. Advances in the Discovery of Novel Antimicrobials Targeting the Assembly of Bacterial Cell Division Protein FtsZ. Eur. J. Med. Chem. 2015, 95, 1–15. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Irudal, S.; AbuAlshaar, A.; Piazza, A.; Trespidi, G.; Barbieri, G.; Makarov, V.; Migliavacca, R.; De Rossi, E.; Buroni, S. Bactericidal and Anti-Biofilm Activity of the FtsZ Inhibitor C109 against Acinetobacter baumannii. Antibiotics 2022, 11, 1571. [Google Scholar] [CrossRef]
- Chai, W.C.; Whittall, J.J.; Polyak, S.W.; Foo, K.; Li, X.; Dutschke, C.J.; Ogunniyi, A.D.; Ma, S.; Sykes, M.J.; Semple, S.J.; et al. Cinnamaldehyde Derivatives Act as Antimicrobial Agents against Acinetobacter baumannii through the Inhibition of Cell Division. Front. Microbiol. 2022, 13, 967949. [Google Scholar] [CrossRef]
- Talat, A.; Khan, A.U. Artificial Intelligence as a Smart Approach to Develop Antimicrobial Drug Molecules: A Paradigm to Combat Drug-Resistant Infections. Drug Discov. Today 2023, 28, 103491. [Google Scholar] [CrossRef]
- Liu, G.; Catacutan, D.B.; Rathod, K.; Swanson, K.; Jin, W.; Mohammed, J.C.; Chiappino-Pepe, A.; Syed, S.A.; Fragis, M.; Rachwalski, K.; et al. Deep Learning-Guided Discovery of an Antibiotic Targeting Acinetobacter baumannii. Nat. Chem. Biol. 2023, 19, 1342–1350. [Google Scholar] [CrossRef]
- Marletta, S.; L’Imperio, V.; Eccher, A.; Antonini, P.; Santonicco, N.; Girolami, I.; Tos, A.P.D.; Sbaraglia, M.; Pagni, F.; Brunelli, M.; et al. Artificial Intelligence-Based Tools Applied to Pathological Diagnosis of Microbiological Diseases. Pathol. Res. Pract. 2023, 243, 154362. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Name of the Drug | Mechanism of Action | Clinical Trials |
|---|---|---|
| Sulbactam/durlobactam (SD, Xacduro®) | β-lactamase inhibitor combined with a non-beta-lactam beta-lactamase inhibitor. | 1. A randomized, active-controlled study to evaluate the efficacy and safety of intravenous sulbactam/etx2514 in the treatment of patients with infections caused by Acinetobacter baumannii-calcoaceticus complex (ATTACK) (NCT03894046). 2. A double-blind, randomized, placebo-controlled study to evaluate the safety and efficacy of intravenous sulbactam/etx2514 in the treatment of hospitalized adults with complicated urinary tract infections, including acute pyelonephritis (NCT03445195). |
| Cefepime/zidebactam (FEP-ZID; WCK5222) | Fourth-generation cephalosporin that disrupts the synthesis of the peptidoglycan layer combined with a non-β-lactam β-lactamase inhibitor. | 1. A randomized, double-blind, placebo-controlled study to evaluate the safety, tolerability, and pharmacokinetics of multiple escalating doses of intravenous WCK 5222 (zidebactam and cefepime) in healthy adult human subjects (NCT02707107). 2. A phase 1, open-label, single-dose study to investigate the pharmacokinetics of intravenous WCK 5222 (FEP-ZID) in patients with renal impairment (NCT02942810). 3. A phase 1, multiple-dose, open-label study to determine and compare plasma and intrapulmonary concentrations of WCK 5222 (cefepime and zidebactam) in healthy adult human subjects (NCT03630094). 4. A randomized, double-blind, double-dummy, placebo- and positive-controlled, crossover study to evaluate the effect of WCK 5222 on the QT/QTc interval in healthy volunteers (NCT03554304). 5. A phase 3, randomized, double-blind, multicenter, comparative study to determine the efficacy and safety of cefepime/zidebactam vs. meropenem in the treatment of complicated urinary tract infection or acute pyelonephritis in adults (NCT04979806). |
| Imipenem/cilastatin/funobactam | Broad-spectrum carbapenem, combined with a dehydropeptidase I inhibitor and a novel diazabicyclooctane. | 1. Four phase 1 studies evaluated pk properties, safety, and tolerability of funobactam, alone and in combination with imipenem/cilastatin (NCT04801043, NCT04802863, NCT04787562, and NCT04482569). 2. An ongoing multicenter, randomized, double-blind, double-dummy, comparative, phase 3 study evaluating the efficacy and safety of imipenem/cilastatin/funobactam vs. meropenem in hospitalized adults with complicated urinary tract infections, including acute pyelonephritis (NCT05204368). 3. A multicenter, randomized, double-blind, comparative, phase 3 study assessing the efficacy and safety of iv imipenem/cilastatin/funobactam vs. imipenem/cilastatin/relebactam in adults with hospital-acquired bacterial pneumonia or ventilator-associated bacterial pneumonia (NCT05204563). |
| Xeruborbactam (XER, QPX7728) | Ultra-broad-spectrum cyclic boronate inhibitor of serine and MBL. | 1. A phase 1, randomized, double-blind, placebo-controlled, ascending single and multiple-dose study of the safety, tolerability, and pharmacokinetics of intravenous (IV) QPX7728 alone and in combination with QPX2014 in healthy adult subjects (NCT04380207). 2. A phase 1, randomized, double-blind, placebo-controlled, ascending single- and multiple-dose study of the safety, tolerability, and pharmacokinetics of oral QPX7831 in healthy adult subjects (NCT04578873). 3. A phase 1, randomized, double-blind, single-dose, drug–drug interaction study to determine the impact of co-administration of QPX7728 on the pharmacokinetics of QPX2014 in healthy adult subjects (NCT05072444). 4. A phase 1, open-label, drug–drug interaction, and randomized, double-blind, controlled, multiple-dose pharmacokinetics and safety study of xeruborbactam oral prodrug (QPX7831) in combination with ceftibuten in healthy adult participants (NCT06079775). 5. A phase 1, open-label, single-dose study to determine the safety and pharmacokinetics of oravance (ceftibuten/xeruborbactam oral prodrug [QPX7831]) in participants with renal impairment (NCT06157242). |
| Zosurabalpin (RG6006) | Macrocyclic peptides that inhibit LPS transportation. | 1. Phase 1, multicenter, single-dose, uncontrolled, open-label (NCT05614895). 2. Phase 1, randomized, sponsor-open, adaptive, single- and multiple-ascending dose, placebo-controlled study (NCT04605718). |
| Apramycin (EBL-1003) | Amynoglicoside derivate. | A phase 1, open-label study to evaluate the plasma PK profile of apramycin and lung penetration of apramycin in epithelial lining fluid and alveolar macrophages after a single IV apramycin dose in healthy subjects, a secondary objective is to assess safety and tolerability (NCT05590728). |
| Zifanocycline (KBP-7072) | Third-generation tetracycline. | Four phase 1 studies assessing safety and tolerability (NCT02454361, NCT05507463, NCT04532957, NCT02654626). |
| MRX-8 QPX9003 SPR206 | Polymixins. | Phase 1 studies to assess safety and tolerability (NCT04649541, NCT04808414, and NCT04868292). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, A.; Augello, E.; Stracquadanio, S.; Bellanca, C.M.; Cosentino, F.; Spampinato, S.; Cantarella, G.; Bernardini, R.; Stefani, S.; Cacopardo, B.; et al. Unveiling the Secrets of Acinetobacter baumannii: Resistance, Current Treatments, and Future Innovations. Int. J. Mol. Sci. 2024, 25, 6814. https://doi.org/10.3390/ijms25136814
Marino A, Augello E, Stracquadanio S, Bellanca CM, Cosentino F, Spampinato S, Cantarella G, Bernardini R, Stefani S, Cacopardo B, et al. Unveiling the Secrets of Acinetobacter baumannii: Resistance, Current Treatments, and Future Innovations. International Journal of Molecular Sciences. 2024; 25(13):6814. https://doi.org/10.3390/ijms25136814
Chicago/Turabian StyleMarino, Andrea, Egle Augello, Stefano Stracquadanio, Carlo Maria Bellanca, Federica Cosentino, Serena Spampinato, Giuseppina Cantarella, Renato Bernardini, Stefania Stefani, Bruno Cacopardo, and et al. 2024. "Unveiling the Secrets of Acinetobacter baumannii: Resistance, Current Treatments, and Future Innovations" International Journal of Molecular Sciences 25, no. 13: 6814. https://doi.org/10.3390/ijms25136814
APA StyleMarino, A., Augello, E., Stracquadanio, S., Bellanca, C. M., Cosentino, F., Spampinato, S., Cantarella, G., Bernardini, R., Stefani, S., Cacopardo, B., & Nunnari, G. (2024). Unveiling the Secrets of Acinetobacter baumannii: Resistance, Current Treatments, and Future Innovations. International Journal of Molecular Sciences, 25(13), 6814. https://doi.org/10.3390/ijms25136814