
Ectopic Activation of Fgf8 in Dental Mesenchyme Causes Incisor Agenesis and Molar Microdontia
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
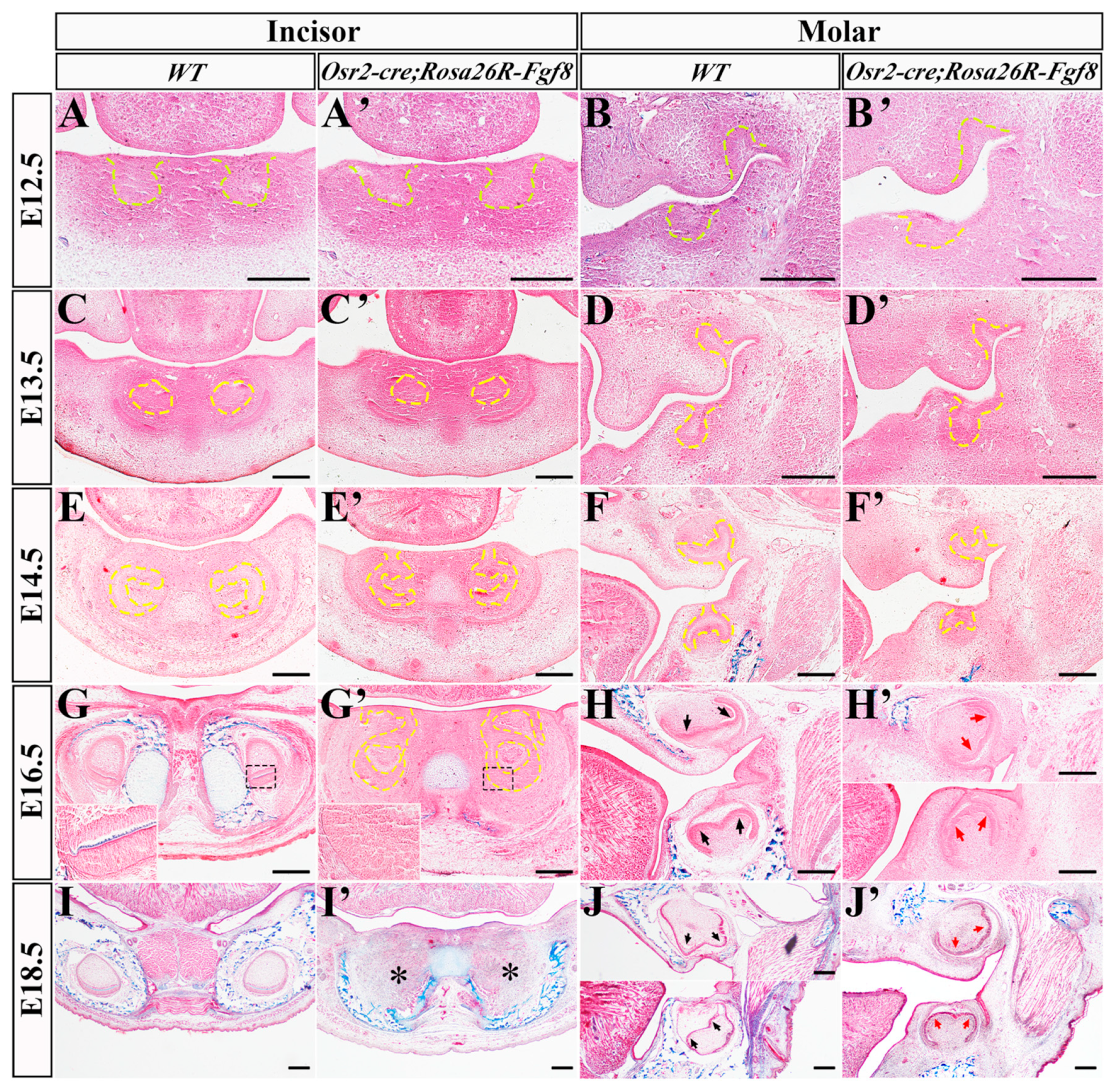
2.1. The Regressed Incisor Germs in Osr2-creKI; Rosa26R-Fgf8 Mice
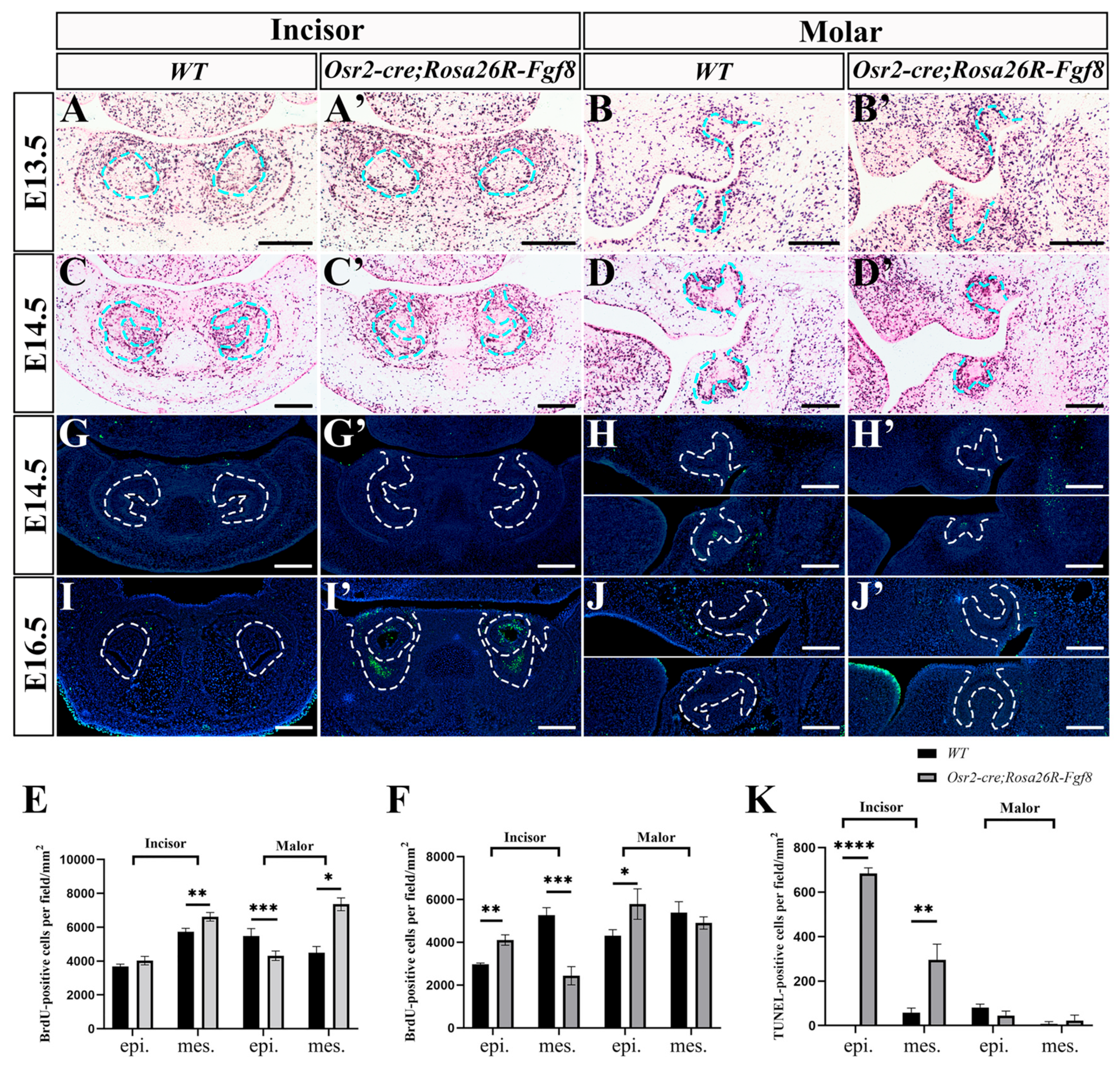
2.2. Enhanced Cell Apoptosis in Osr2-creKI;Rosa26R-Fgf8 Incisor Germs
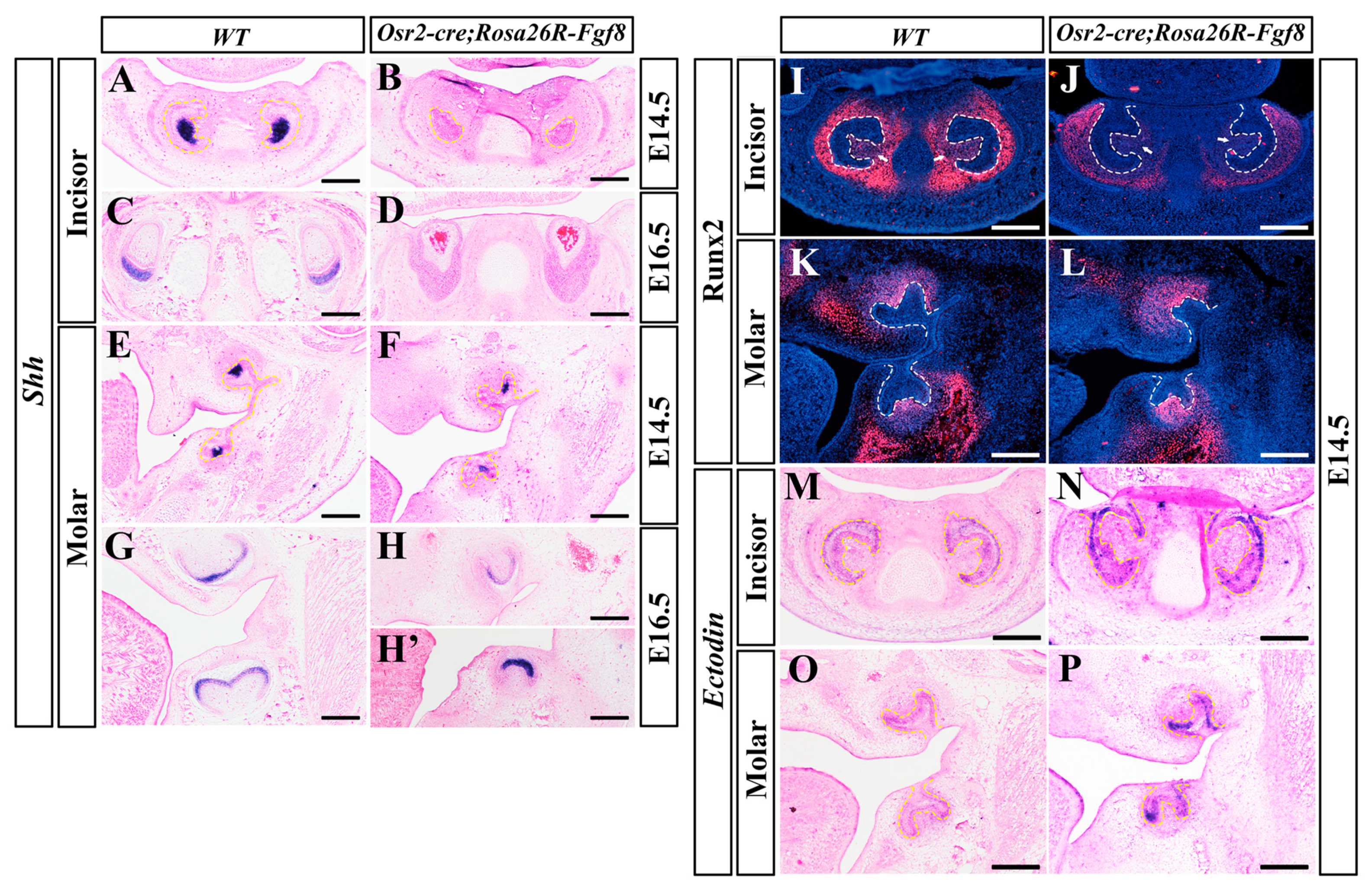
2.3. Differential Gene Expression between Osr2-creKI; Rosa26R-Fgf8 Incisor and Molar Germs
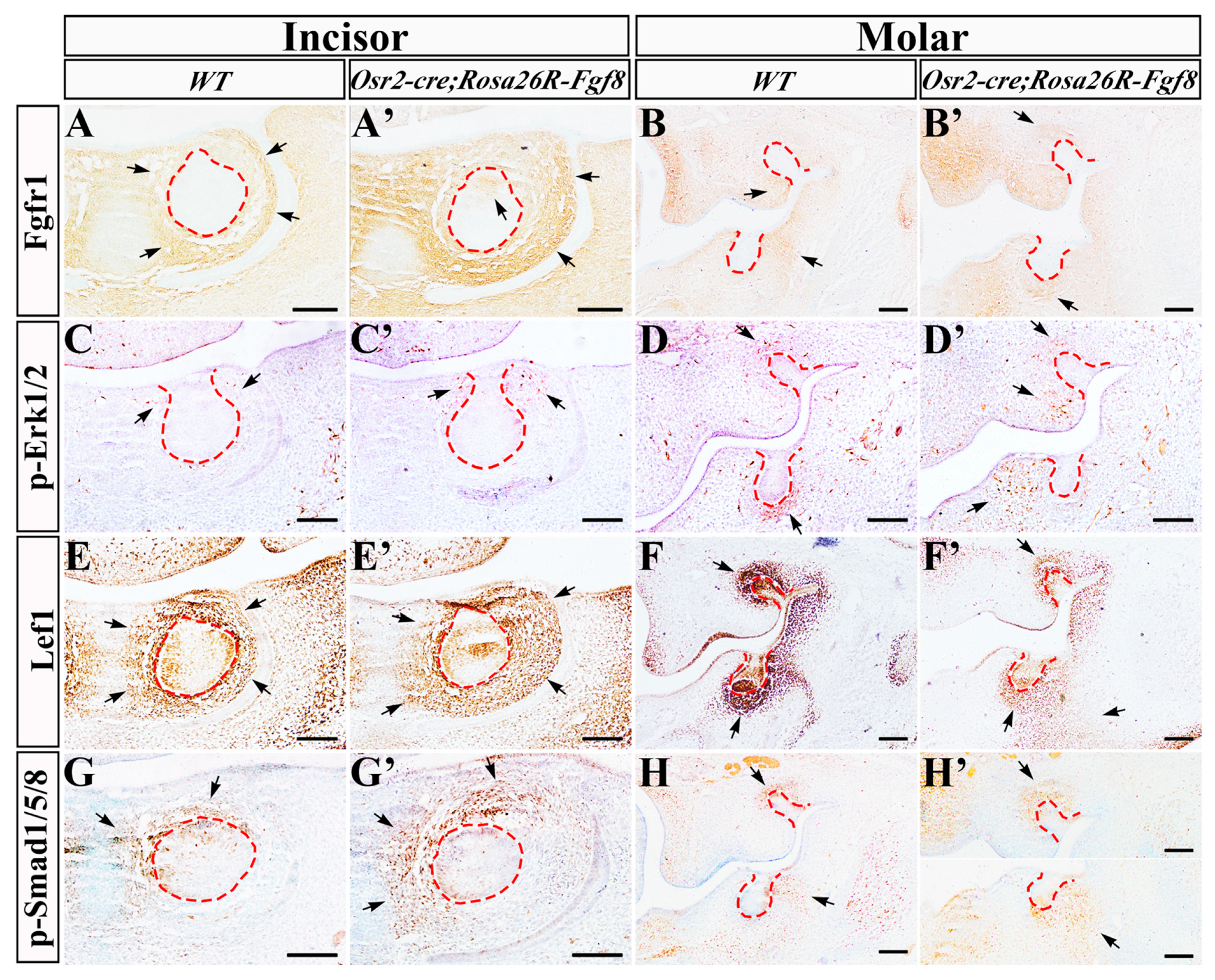
2.4. Altered Gene Expression in Osr2-creKI; Rosa26R-Fgf8 Incisor and Molar Germs
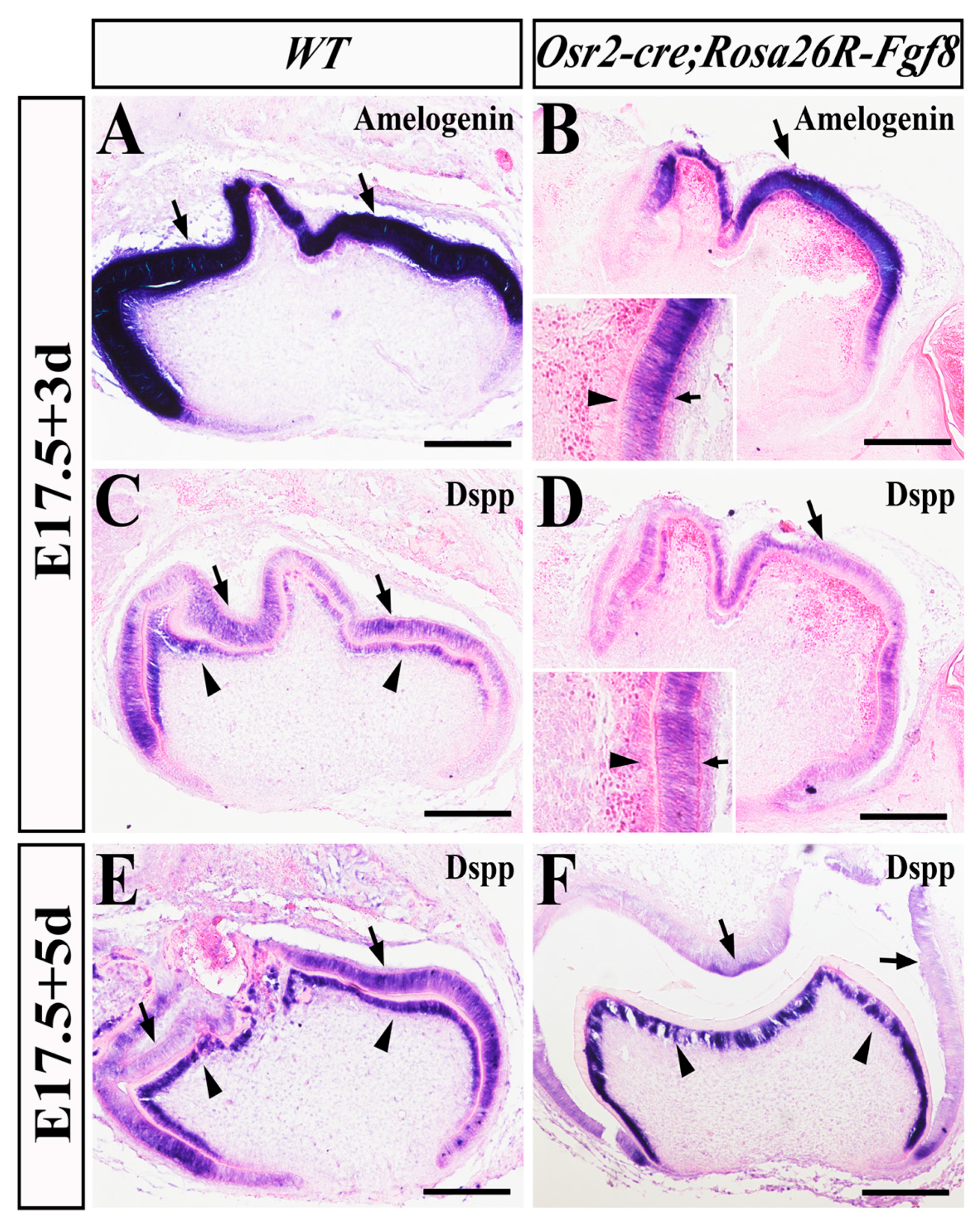
2.5. Delayed Odontoblast Differentiation and Enhanced Ameloblast Degeneration in Osr2-creKI; Rosa26R-Fgf8 Molars
2.6. Impaired Mineralization and Reduced Size of Osr2-creKI; Rosa26R-Fgf8 Molars
3. Discussion
4. Materials and Methods
4.1. Mouse Lines
4.2. Histological and Cryostat Section
4.3. BrdU Labeling Assay
4.4. TUNEL Assay
4.5. Statistical Assay
4.6. In Situ Hybridization
4.7. Immunohistochemistry and Immunofluorescence
4.8. X-ray Plain Film for Sub-Renal Capsule Implanted Molar Germs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, T.; Klein, O.D. Molecular and cellular mechanisms of tooth development, homeostasis and repair. Development 2020, 147, dev184754. [Google Scholar] [CrossRef] [PubMed]
- Porntaveetus, T.; Otsuka-Tanaka, Y.; Basson, M.A.; Moon, A.M.; Sharpe, P.T.; Ohazama, A. Expression of fibroblast growth factors (Fgfs) in murine tooth development. J. Anat. 2011, 218, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Prochazka, J.; Goodwin, A.F.; Klein, O.D. Fibroblast growth factor signaling in mammalian tooth development. Odontology 2014, 102, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Prochazka, J.; Prochazkova, M.; Du, W.; Spoutil, F.; Tureckova, J.; Hoch, R.; Shimogori, T.; Sedlacek, R.; Rubenstein, J.L.; Wittmann, T.; et al. Migration of Founder Epithelial Cells Drives Proper Molar Tooth Positioning and Morphogenesis. Dev. Cell 2015, 35, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Klein, O.D.; Minowada, G.; Peterkova, R.; Kangas, A.; Yu, B.D.; Lesot, H.; Peterka, M.; Jernvall, J.; Martin, G.R. Sprouty genes control diastema tooth development via bidirectional antagonism of epithelial-mesenchymal FGF signaling. Dev. Cell 2006, 11, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, P.; Charles, C.; Ahn, Y.; Seidel, K.; Jheon, A.; Ganss, B.; Krumlauf, R.; Viriot, L.; Klein, O.D. Downregulation of FGF Signaling by Spry4 Overexpression Leads to Shape Impairment, Enamel Irregularities, and Delayed Signaling Center Formation in the Mouse Molar. JBMR Plus 2019, 3, e10205. [Google Scholar] [CrossRef] [PubMed]
- Häärä, O.; Harjunmaa, E.; Lindfors, P.H.; Huh, S.H.; Fliniaux, I.; Åberg, T.; Jernvall, J.; Ornitz, D.M.; Mikkola, M.L.; Thesleff, I. Ectodysplasin regulates activator-inhibitor balance in murine tooth development through Fgf20 signaling. Development 2012, 139, 3189–3199. [Google Scholar] [CrossRef] [PubMed]
- Aberg, T.; Wang, X.P.; Kim, J.H.; Yamashiro, T.; Bei, M.; Rice, R.; Ryoo, H.M.; Thesleff, I. Runx2 mediates FGF signaling from epithelium to mesenchyme during tooth morphogenesis. Dev. Biol. 2004, 270, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.P.; Suomalainen, M.; Felszeghy, S.; Zelarayan, L.C.; Alonso, M.T.; Plikus, M.V.; Maas, R.L.; Chuong, C.M.; Schimmang, T.; Thesleff, I. An integrated gene regulatory network controls stem cell proliferation in teeth. PLoS Biol. 2007, 5, e159. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Toyono, T.; Toyoshima, K.; Yamasaki, M.; Itoh, N.; Kato, S.; Sekine, K.; Ohuchi, H. FGF10 maintains stem cell compartment in developing mouse incisors. Development 2002, 129, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Nunn, J.H.; Carter, N.E.; Gillgrass, T.J.; Hobson, R.S.; Jepson, N.J.; Meechan, J.G.; Nohl, F.S. The interdisciplinary management of hypodontia: Background and role of paediatric dentistry. Br. Dent. J. 2003, 194, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Maurice, J.M.; Craig, W.D. Tooth agenesis: An overview of diagnosis, aetiology and management. Jpn. Dent. Sci. Rev. 2023, 59, 209–218. [Google Scholar]
- Järvinen, E.; Shimomura-Kuroki, J.; Balic, A.; Jussila, M.; Thesleff, I. Mesenchymal Wnt/beta-catenin signaling limits tooth number. Development 2018, 145, dev158048. [Google Scholar] [CrossRef] [PubMed]
- Hlouskova, A.; Bielik, P.; Bonczek, O.; Balcar, V.J.; Šerý, O. Mutations in AXIN2 gene as a risk factor for tooth agenesis and cancer: A review. Neuro Endocrinol. Lett. 2017, 38, 131–137. [Google Scholar] [PubMed]
- Ryoo, H.M.; Wang, X.P. Control of tooth morphogenesis by Runx2. Crit. Rev. Eukaryot. Gene Expr. 2006, 16, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, A.H.; Antoun, J.S.; Thomson, W.M.; Merriman, T.R.; Farella, M. Hypodontia: An Update on Its Etiology, Classification, and Clinical Management. BioMed. Res. Int. 2017, 2017, 9378325. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.L. Microdontia. Oral Surg. Oral Med. Oral Pathol. 1967, 23, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Reis, L.M.; Maheshwari, M.; Capasso, J.; Atilla, H.; Dudakova, L.; Thompson, S.; Zitano, L.; Lay-Son, G.; Lowry, R.B.; Black, J.; et al. Axenfeld-Rieger syndrome: More than meets the eye. J. Med. Genet. 2023, 60, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.H.; Kyun, C.J.; Kim, J.W.; Lee, S. Lacrimo-auriculo-dento-digital syndrome: A novel mutation in a Korean family and review of literature. Mol. Genet. Genom. Med. 2020, 8, e1412. [Google Scholar] [CrossRef] [PubMed]
- Moussaid, Y.; Griffiths, D.; Richard, B.; Dieux, A.; Lemerrer, M.; Léger, J.; Lacombe, D.; Bailleul-Forestier, I. Oral manifestations of patients with Kenny-Caffey Syndrome. Eur. J. Med. Genet. 2012, 55, 441–445. [Google Scholar] [PubMed]
- Jamshidi, F.; Shokouhian, E.; Mohseni, M.; Kahrizi, K.; Najmabadi, H.; Babanejad, M. Identification of a homozygous frameshift mutation in the FGF3 gene in a consanguineous Iranian family: First report of labyrinthine aplasia, microtia, and microdontia syndrome in Iran and literature review. Mol. Genet. Genom. Med. 2023, 11, e2168. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, A.; Shalehin, N.; Takebe, H.; Shimo, T.; Irie, K. Sonic Hedgehog Signaling and Tooth Development. Int. J. Mol. Sci. 2020, 21, 1587. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Liu, C.; Song, Y.; Ye, W.; He, W.; Yuan, G.; Gu, S.; Lin, C.; Ma, L.; Zhang, Y.; et al. FGF8 signaling sustains progenitor status and multipotency of cranial neural crest-derived mesenchymal cells in vivo and in vitro. J. Mol. Cell Biol. 2015, 7, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gu, S.; Sun, C.; Ye, W.; Song, Z.; Zhang, Y.; Chen, Y. FGF signaling sustains the odontogenic fate of dental mesenchyme by suppressing beta-catenin signaling. Development 2013, 140, 4375–4385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gong, X.; Xu, X.; Wang, X.; Sun, Y. Tooth number abnormality: From bench to bedside. Int. J. Oral Sci. 2023, 15, 5. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Parada, C.; Chai, Y. Cellular and molecular mechanisms of tooth root development. Development 2017, 144, 374–384. [Google Scholar] [CrossRef]
- Hao, Y.; Tang, S.; Yuan, Y.; Liu, R.; Chen, Q. Roles of FGF8 subfamily in embryogenesis and oral-maxillofacial diseases. Int. J. Oncol. 2019, 54, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, J.; Li, N.; Wang, Y.; Zhou, N.; Zhu, L.; Shi, Y.; Wu, Y.; Xiao, J.; Liu, C. Mesenchymal Wnt/beta-catenin signaling induces Wnt and BMP antagonists in dental epithelium. Organogenesis 2019, 15, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, X.; Xu, X.; Mayo, J.; Bringas, P., Jr.; Jiang, R.; Wang, S.; Chai, Y. SMAD4-mediated WNT signaling controls the fate of cranial neural crest cells during tooth morphogenesis. Development 2011, 138, 1977–1989. [Google Scholar] [CrossRef] [PubMed]
- Mashyakhy, M.; Alkahtani, A.; Abumelha, A.S.; Sharroufna, R.J.; Alkahtany, M.F.; Jamal, M.; Robaian, A.; Binalrimal, S.; Chohan, H.; Patil, V.R.; et al. Taurine Augments Telomerase Activity and Promotes Chondrogenesis in Dental Pulp Stem Cells. J. Pers. Med. 2021, 11, 491. [Google Scholar] [CrossRef]
- Zhang, W.; Yelick, P.C. Tooth Repair and Regeneration: Potential of Dental Stem Cells. Trends. Mol. Med. 2021, 27, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ou, Q.; Shi, S.; Shao, C. Immunomodulatory properties of mesenchymal stem cells/dental stem cells and their therapeutic applications. Cell. Mol. Immunol. 2023, 20, 558–569. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhou, N.; Li, N.; Xu, T.; Chen, X.; Zhou, H.; Xie, A.; Liu, H.; Zhu, L.; Wang, S.; et al. Disrupted tenogenesis in masseter as a potential cause of micrognathia. Int. J. Oral Sci. 2022, 14, 50. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, J.; Xu, T.; Yang, S.; Wang, X.; Zhu, L.; Li, N.; Liu, B.; Xiao, J.; Liu, C. Ectopic Activation of Fgf8 in Dental Mesenchyme Causes Incisor Agenesis and Molar Microdontia. Int. J. Mol. Sci. 2024, 25, 7045. https://doi.org/10.3390/ijms25137045
Wang Y, Wang J, Xu T, Yang S, Wang X, Zhu L, Li N, Liu B, Xiao J, Liu C. Ectopic Activation of Fgf8 in Dental Mesenchyme Causes Incisor Agenesis and Molar Microdontia. International Journal of Molecular Sciences. 2024; 25(13):7045. https://doi.org/10.3390/ijms25137045
Chicago/Turabian StyleWang, Yu, Jingjing Wang, Tian Xu, Shuhui Yang, Xinran Wang, Lei Zhu, Nan Li, Bo Liu, Jing Xiao, and Chao Liu. 2024. "Ectopic Activation of Fgf8 in Dental Mesenchyme Causes Incisor Agenesis and Molar Microdontia" International Journal of Molecular Sciences 25, no. 13: 7045. https://doi.org/10.3390/ijms25137045
APA StyleWang, Y., Wang, J., Xu, T., Yang, S., Wang, X., Zhu, L., Li, N., Liu, B., Xiao, J., & Liu, C. (2024). Ectopic Activation of Fgf8 in Dental Mesenchyme Causes Incisor Agenesis and Molar Microdontia. International Journal of Molecular Sciences, 25(13), 7045. https://doi.org/10.3390/ijms25137045