
Netrin-1 Is an Important Mediator in Microglia Migration
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Netrin-1 Promotes BV2 Elongation and Migration
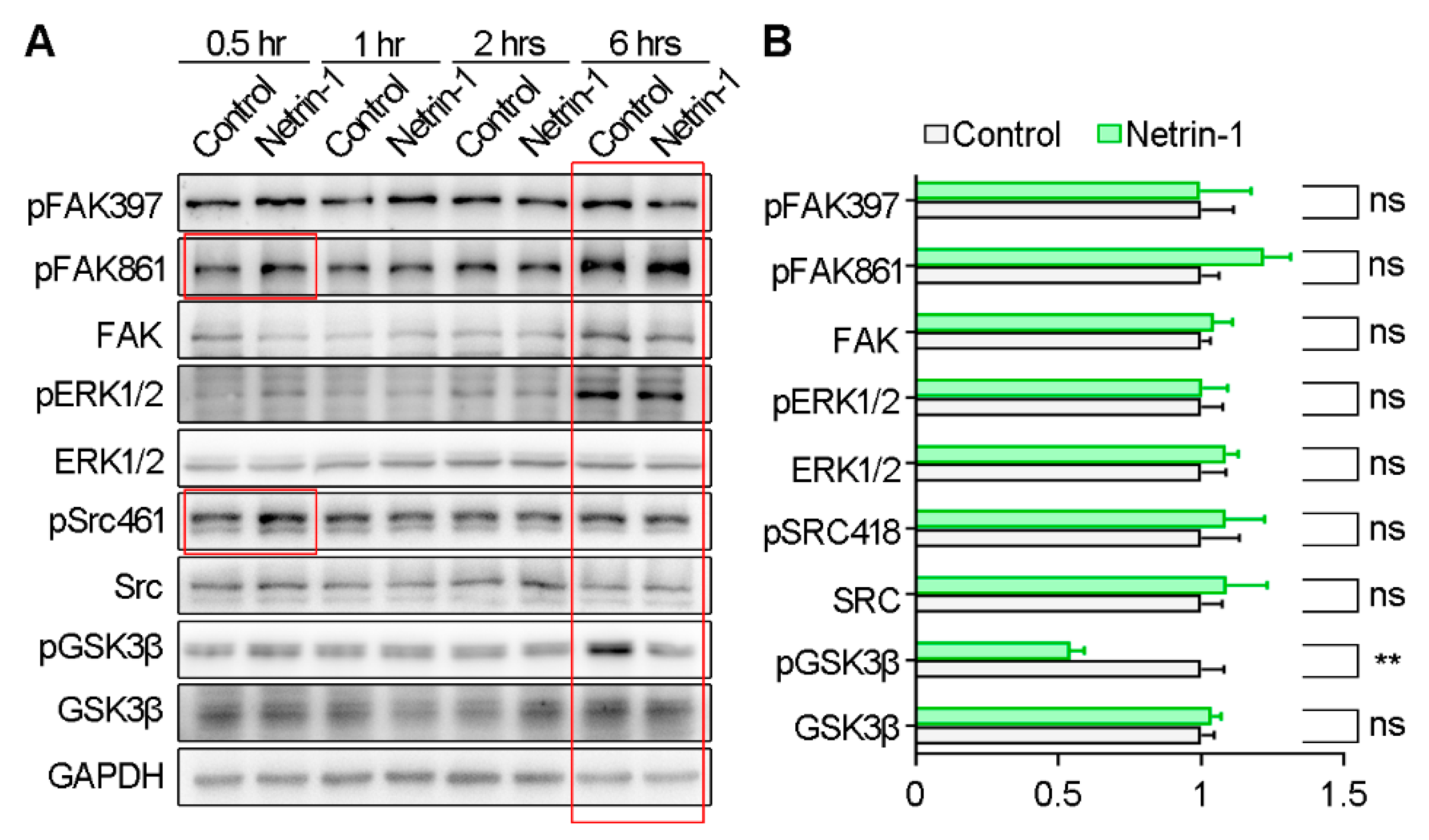
2.2. Netrin-1 Increases GSK3β Activity by Suppressing GSK3β Phosphorylation at Serine 9
2.3. Netrin-1 Regulates BV2 Elongation and Migration through Activating GSK3β Activity
2.4. Integrin α6/β1 Is Highly Expressed in Microglia and Interacts with Netrin-1
2.5. Activating Integrin α6/β1 Mimics Netrin-1-Induced GSK3β Activity as well as BV2 Elongation and Migration
2.6. Netrin-1 Is Highly Expressed in the Developing Cerebral Cortex
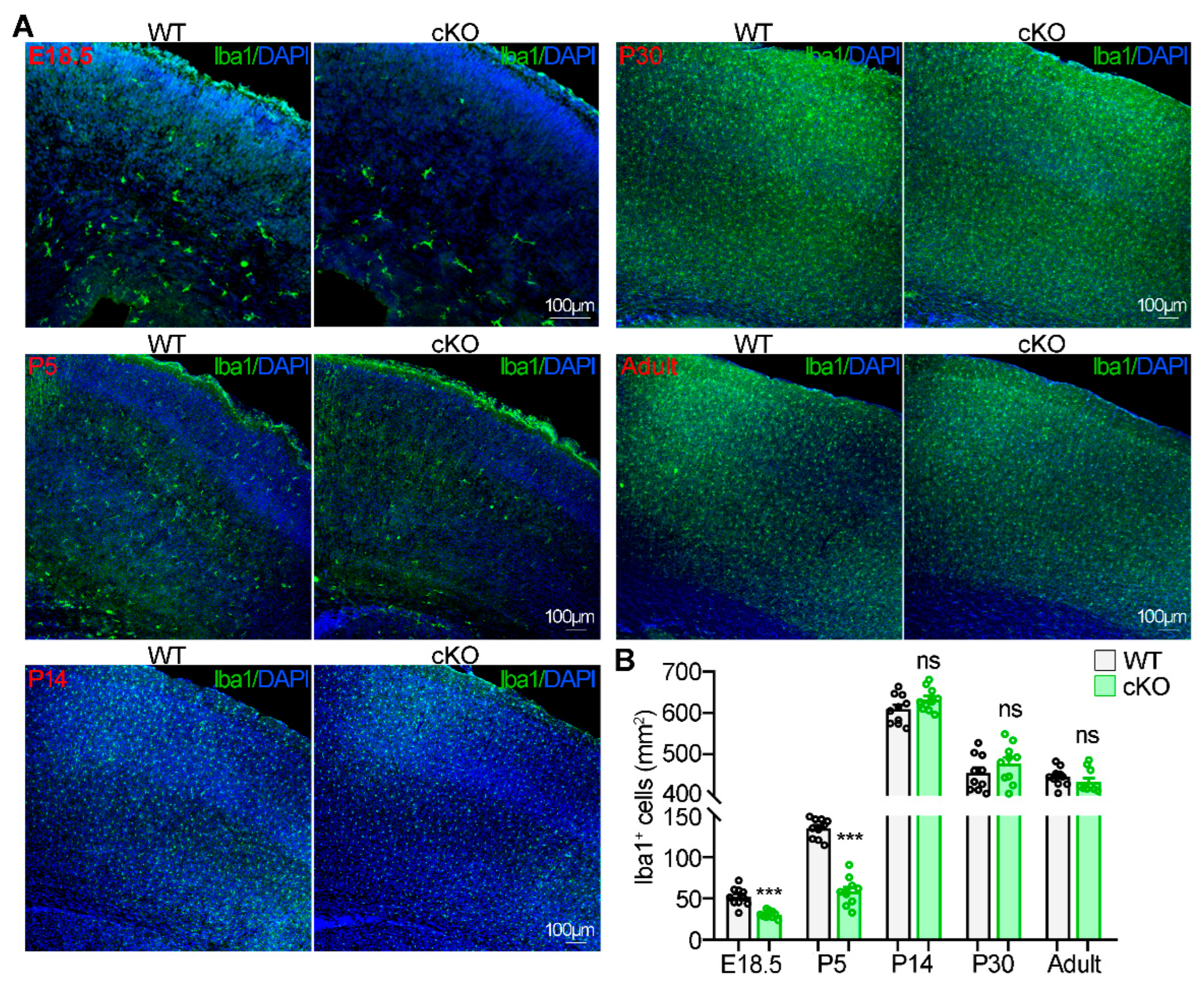
2.7. Netrin-1 Knockout Suppresses Microglia Migration into the Cerebral Cortex during Early Brain Development
3. Discussion
4. Materials and Method
4.1. Animals
4.2. Reagents
4.3. Cell Culture and Collection of Netrin-1 Conditioned Medium
4.4. In Vitro Cell Migration Assay
4.5. Immunostaining
4.6. Immunoblotting
4.7. Microscale Thermophoresis (MST) Measurement
4.8. Imaging and Quantification
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kierdorf, K.; Erny, D.; Goldmann, T.; Sander, V.; Schulz, C.; Perdiguero, E.G.; Wieghofer, P.; Heinrich, A.; Riemke, P.; Hölscher, C.; et al. Microglia emerge from erythromyeloid precursors via Pu.1- and Irf8-dependent pathways. Nat. Neurosci. 2013, 16, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Martínez-Cerdeño, V.; Noctor, S.C. Microglia Regulate the Number of Neural Precursor Cells in the Developing Cerebral Cortex. J. Neurosci. 2013, 33, 4216–4233. [Google Scholar] [CrossRef] [PubMed]
- Shigemoto-Mogami, Y.; Hoshikawa, K.; Goldman, J.E.; Sekino, Y.; Sato, K. Microglia Enhance Neurogenesis and Oligodendrogenesis in the Early Postnatal Subventricular Zone. J. Neurosci. 2014, 34, 2231–2243. [Google Scholar] [CrossRef] [PubMed]
- Squarzoni, P.; Oller, G.; Hoeffel, G.; Pont-Lezica, L.; Rostaing, P.; Low, D.; Bessis, A.; Ginhoux, F.; Garel, S. Microglia Modulate Wiring of the Embryonic Forebrain. Cell Rep. 2014, 8, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Pont-Lezica, L.; Beumer, W.; Colasse, S.; Drexhage, H.; Versnel, M.; Bessis, A. Microglia shape corpus callosum axon tract fasciculation: Functional impact of prenatal inflammation. Eur. J. Neurosci. 2014, 39, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The Classical Complement Cascade Mediates CNS Synapse Elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic Pruning by Microglia Is Necessary for Normal Brain Development. Science 2011, 333, 1456–1458. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Ellis, S.E.; Ashar, F.N.; Moes, A.; Bader, J.S.; Zhan, J.; West, A.B.; Arking, D.E. Transcriptome analysis reveals dysregulation of innate immune response genes and neuronal activity-dependent genes in autism. Nat. Commun. 2014, 5, 5748. [Google Scholar] [CrossRef]
- Kenk, M.; Selvanathan, T.; Rao, N.; Suridjan, I.; Rusjan, P.; Remington, G.; Meyer, J.H.; Wilson, A.A.; Houle, S.; Mizrahi, R. Imaging Neuroinflammation in Gray White Matter in Schizophrenia: An In-Vivo PETStudy with [18F.]-FEPPA. Schizophr. Bull. 2015, 41, 85–93. [Google Scholar] [CrossRef]
- Xu, J.; Wang, T.; Wu, Y.; Jin, W.; Wen, Z. Microglia Colonization of Developing Zebrafish Midbrain Is Promoted by Apoptotic Neuron and Lysophosphatidylcholine. Dev. Cell 2016, 38, 214–222. [Google Scholar] [CrossRef]
- Arnò, B.; Grassivaro, F.; Rossi, C.; Bergamaschi, A.; Castiglioni, V.; Furlan, R.; Greter, M.; Favaro, R.; Comi, G.; Becher, B.; et al. Neural progenitor cells orchestrate microglia migration and positioning into the developing cortex. Nat. Commun. 2014, 5, 5611. [Google Scholar] [CrossRef]
- Wu, S.; Xue, R.; Hassan, S.; Nguyen, T.M.L.; Wang, T.; Pan, H.; Xu, J.; Liu, Q.; Zhang, W.; Wen, Z. Il34-Csf1r Pathway Regulates the Migration and Colonization of Microglial Precursors. Dev. Cell 2018, 46, 552–563.e4. [Google Scholar] [CrossRef] [PubMed]
- Brignani, S.; Raj, D.D.; Schmidt, E.R.; Düdükcü, Ö.; Adolfs, Y.; De Ruiter, A.A.; Rybiczka-Tesulov, M.; Verhagen, M.G.; van der Meer, C.; Broekhoven, M.H.; et al. Remotely Produced and Axon-Derived Netrin-1 Instructs GABAergic Neuron Migration and Dopaminergic Substantia Nigra Development. Neuron 2020, 107, 684–702. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-L.; Peng, Y.; Zhao, Y.; Lan, Y.-S.; Wang, B.; Zhao, L.; Sun, D.; Pan, J.-X.; Dong, Z.-Q.; Mei, L.; et al. Myosin X Interaction with KIF13B, a Crucial Pathway for Netrin-1-Induced Axonal Development. J. Neurosci. 2020, 40, 9169–9185. [Google Scholar] [CrossRef] [PubMed]
- van Gils, J.M.; Derby, M.C.; Fernandes, L.R.; Ramkhelawon, B.; Ray, T.D.; Rayner, K.J.; Parathath, S.; Distel, E.; Feig, J.L.; Alvarez-Leite, J.I.; et al. The neuroimmune guidance cue netrin-1 promotes atherosclerosis by inhibiting the emigration of macrophages from plaques. Nat. Immunol. 2012, 13, 136–143. [Google Scholar] [CrossRef]
- Aherne, C.M.; Collins, C.B.; Masterson, J.C.; Tizzano, M.; Boyle, T.A.; Westrich, J.A.; Parnes, J.A.; Furuta, G.T.; Rivera-Nieves, J.; Eltzschig, H.K. Neuronal guidance molecule netrin-1 attenuates inflammatory cell trafficking during acute experimental colitis. Gut 2011, 61, 695–705. [Google Scholar] [CrossRef]
- Liu, L.; Liu, K.-J.; Cao, J.-B.; Yang, J.; Yu, H.-L.; He, X.-X.; He, Z.-X.; Zhu, X.-J. A Novel Netrin-1-Derived Peptide Enhances Protection against Neuronal Death and Mitigates of Intracerebral Hemorrhage in Mice. Int. J. Mol. Sci. 2021, 22, 4829. [Google Scholar] [CrossRef]
- Ren, X.-R.; Ming, G.-L.; Xie, Y.; Hong, Y.; Sun, D.-M.; Zhao, Z.-Q.; Feng, Z.; Wang, Q.; Shim, S.; Chen, Z.-F.; et al. Focal adhesion kinase in netrin-1 signaling. Nat. Neurosci. 2004, 7, 1204–1212. [Google Scholar] [CrossRef]
- The Tabula Muris Consortium; Overall coordination; Logistical coordination; Organ collection and processing; Library preparation and sequencing; Computational data analysis; Cell type annotation; Writing group; Supplemental text writing group; Principal investigators. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 2018, 562, 367–372. [Google Scholar] [CrossRef]
- Lee, H.K.; Seo, I.A.; Shin, Y.K.; Lee, S.H.; Seo, S.-Y.; Suh, D.J.; Park, H.T. Netrin-1 Specifically Enhances Cell Spreading on Fibronectin in Human Glioblastoma Cells. Korean J. Physiol. Pharmacol. 2008, 12, 225–230. [Google Scholar] [CrossRef]
- Lee, S.-J.; Jung, Y.H.; Oh, S.Y.; Yong, M.S.; Ryu, J.M.; Han, H.J. Netrin-1 Induces MMP-12-Dependent E-Cadherin Degradation Via the Distinct Activation of PKCα and FAK/Fyn in Promoting Mesenchymal Stem Cell Motility. Stem Cells Dev. 2014, 23, 1870–1882. [Google Scholar] [CrossRef] [PubMed]
- Lemons, M.; Abanto, M.; Dambrouskas, N.; Clements, C.; DeLoughery, Z.; Garozzo, J.; Condic, M. Integrins and cAMP mediate netrin-induced growth cone collapse. Brain Res. 2013, 1537, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.A.; Sanchez-Gomez, P.; Muñoz-Palma, E.; Puvogel, S.; Casas, B.S.; Arriagada, C.; Peña-Villalobos, I.; Lois, P.; Orellana, M.R.; Lubieniecki, F.; et al. The Netrin-1-Neogenin-1 signaling axis controls neuroblastoma cell migration via integrin-β1 and focal adhesion kinase activation. Cell Adhes. Migr. 2021, 15, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Freitas, V.M.; Vilas-Boas, V.F.; Pimenta, D.C.; Loureiro, V.; Juliano, M.A.; Carvalho, M.R.; Pinheiro, J.J.; Camargo, A.C.; Moriscot, A.S.; Hoffman, M.P.; et al. SIKVAV, a laminin α1-Derived Peptide, interacts with integrins and increases protease activity of a human salivary gland adenoid cystic carcinoma cell line through the ERK 1/2 signaling pathway. Am. J. Pathol. 2007, 171, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.S.; Ashour, M.A.; Magdesian, M.H.; Tritsch, N.X.; Harris, S.N.; Christofi, N.; Chemali, R.; Stern, Y.E.; Thompson-Steckel, G.; Gris, P.; et al. Netrin-1 Promotes Excitatory Synaptogenesis between Cortical Neurons by Initiating Synapse Assembly. J. Neurosci. 2013, 33, 17278–17289. [Google Scholar] [CrossRef] [PubMed]
- Easley-Neal, C.; Foreman, O.; Sharma, N.; Zarrin, A.A.; Weimer, R.M. CSF1R Ligands IL-34 and CSF1 Are Differentially Required for Microglia Development and Maintenance in White and Gray Matter Brain Regions. Front. Immunol. 2019, 10, 2199. [Google Scholar] [CrossRef] [PubMed]
- Nikodemova, M.; Kimyon, R.S.; De, I.; Small, A.L.; Collier, L.S.; Watters, J.J. Microglial numbers attain adult levels after undergoing a rapid decrease in cell number in the third postnatal week. J. Neuroimmunol. 2014, 278, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Barry-Carroll, L.; Greulich, P.; Marshall, A.R.; Riecken, K.; Fehse, B.; Askew, K.E.; Li, K.; Garaschuk, O.; Menassa, D.A.; Gomez-Nicola, D. Microglia colonize the developing brain by clonal expansion of highly proliferative progenitors, following allometric scaling. Cell Rep. 2023, 42, 112425. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.M.; Tsenkina, Y.; Arizanovska, D.; Mehlen, P.; Liebl, D.J. DCC/netrin-1 regulates cell death in oligodendrocytes after brain injury. Cell Death Differ. 2023, 30, 397–406. [Google Scholar] [CrossRef]
- Dominici, C.; Moreno-Bravo, J.A.; Puiggros, S.R.; Rappeneau, Q.; Rama, N.; Vieugue, P.; Bernet, A.; Mehlen, P.; Chédotal, A. Floor-plate-derived netrin-1 is dispensable for commissural axon guidance. Nature 2017, 545, 350–354. [Google Scholar] [CrossRef]
- Morcom, L.; Gobius, I.; Marsh, A.P.; Suárez, R.; Lim, J.W.C.; Bridges, C.; Ye, Y.; Fenlon, L.R.; Zagar, Y.; Douglass, A.M.; et al. DCC regulates astroglial development essential for telencephalic morphogenesis and corpus callosum formation. eLife 2021, 10, e61769. [Google Scholar] [CrossRef] [PubMed]
- Milligan, C.E.; Cunningham, T.J.; Levitt, P. Differential Immunochemical Markers Reveal the Normal Distribution of Brain Macrophages and Microglia in the Developing Rat Brain. J. Comp. Neurol. 1991, 314, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Barry-Carroll, L.; Gomez-Nicola, D. The molecular determinants of microglial developmental dynamics. Nat. Rev. Neurosci. 2024, 25, 414–427. [Google Scholar] [CrossRef]
- Ishikawa, T.; Wondimu, Z.; Oikawa, Y.; Gentilcore, G.; Kiessling, R.; Brage, S.E.; Hansson, J.; Patarroyo, M. Laminins 411 and 421 differentially promote tumor cell migration via α6β1 integrin and MCAM (CD146). Matrix Biol. 2014, 38, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Larrieu-Lahargue, F.; Welm, A.L.; Thomas, K.R.; Li, D.Y. Netrin-4 Activates Endothelial Integrin α6β1. Circ. Res. 2011, 109, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Jacques, T.S.; Relvas, J.B.; Nishimura, S.; Pytela, R.; Edwards, G.M.; Streuli, C.H.; Ffrench-Constant, C. Neural precursor cell chain migration and division are regulated through different β1 integrins. Development 1998, 125, 3167–3177. [Google Scholar] [CrossRef] [PubMed]
- Colantonio, L.; Iellem, A.; Clissi, B.; Pardi, R.; Rogge, L.; Sinigaglia, F.; D’Ambrosio, D. Upregulation of integrin α6/β1 and chemokine receptor CCR1 by interleukin-12 promotes the migration of human type 1 helper T cells. Blood 1999, 94, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- Górska, A.; Mazur, A.J. Integrin-linked kinase (ILK): The known vs. the unknown and perspectives. Cell. Mol. Life Sci. 2022, 79, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Di, L. Glycogen synthesis and beyond, a comprehensive review of GSK3 as a key regulator of metabolic pathways and a therapeutic target for treating metabolic diseases. Med. Res. Rev. 2021, 42, 946–982. [Google Scholar] [CrossRef]
- Hajka, D.; Budziak, B.; Pietras, Ł.; Duda, P.; McCubrey, J.A.; Gizak, A. GSK3 as a Regulator of Cytoskeleton Architecture: Consequences for Health and Disease. Cells 2021, 10, 2092. [Google Scholar] [CrossRef]
- Planas, A.M. Role of microglia in stroke. Glia 2024, 72, 1016–1053. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Huang, L.; Enkhjargal, B.; Reis, C.; Wan, W.; Tang, J.; Cheng, Y.; Zhang, J.H. Recombinant Netrin-1 binding UNC5B receptor attenuates neuroinflammation and brain injury via PPARγ/NFκB signaling pathway after subarachnoid hemorrhage in rats. Brain, Behav. Immun. 2018, 69, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Liu, X.-Y.; Zhao, L.; Hu, Z.-B.; Li, Z.-X.; Liu, W.-T.; Song, N.-N.; Hu, Y.-Q.; Jiang, L.-P.; Zhang, L.; et al. Ventricular Netrin-1 deficiency leads to defective pyramidal decussation and mirror movement in mice. Cell Death Dis. 2024, 15, 343. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-L.; Liu, X.; Yin, Y.; Liu, X.-N.; Feng, Y.-Y.; Tahir, M.M.; Miao, X.-Z.; He, X.-X.; He, Z.-X.; Zhu, X.-J. Netrin-1 Is an Important Mediator in Microglia Migration. Int. J. Mol. Sci. 2024, 25, 7079. https://doi.org/10.3390/ijms25137079
Yu H-L, Liu X, Yin Y, Liu X-N, Feng Y-Y, Tahir MM, Miao X-Z, He X-X, He Z-X, Zhu X-J. Netrin-1 Is an Important Mediator in Microglia Migration. International Journal of Molecular Sciences. 2024; 25(13):7079. https://doi.org/10.3390/ijms25137079
Chicago/Turabian StyleYu, Hua-Li, Xiu Liu, Yue Yin, Xiao-Nuo Liu, Yu-Yao Feng, Muhammad Mateen Tahir, Xin-Zhi Miao, Xiao-Xiao He, Zi-Xuan He, and Xiao-Juan Zhu. 2024. "Netrin-1 Is an Important Mediator in Microglia Migration" International Journal of Molecular Sciences 25, no. 13: 7079. https://doi.org/10.3390/ijms25137079