
Latrophilins as Downstream Effectors of Androgen Receptors including a Splice Variant, AR-V7, Induce Prostate Cancer Progression
 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
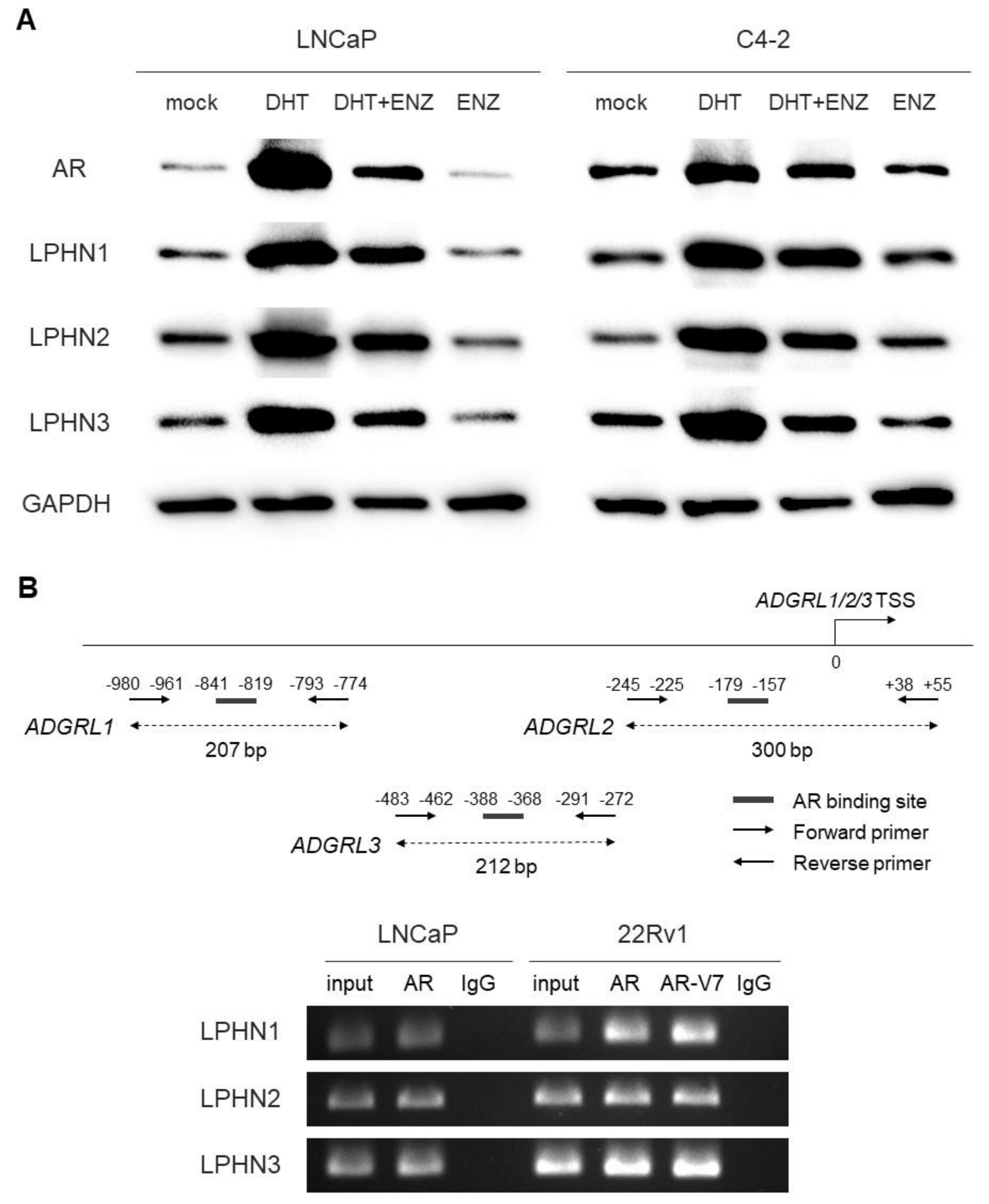
2.1. Effects of Androgen on the Expression of LPHNs
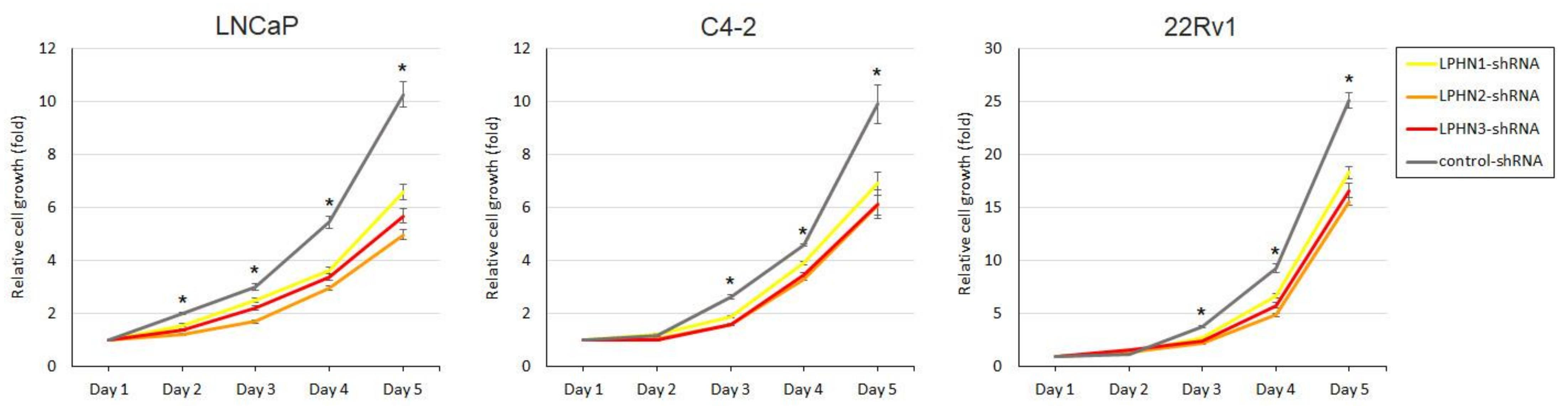
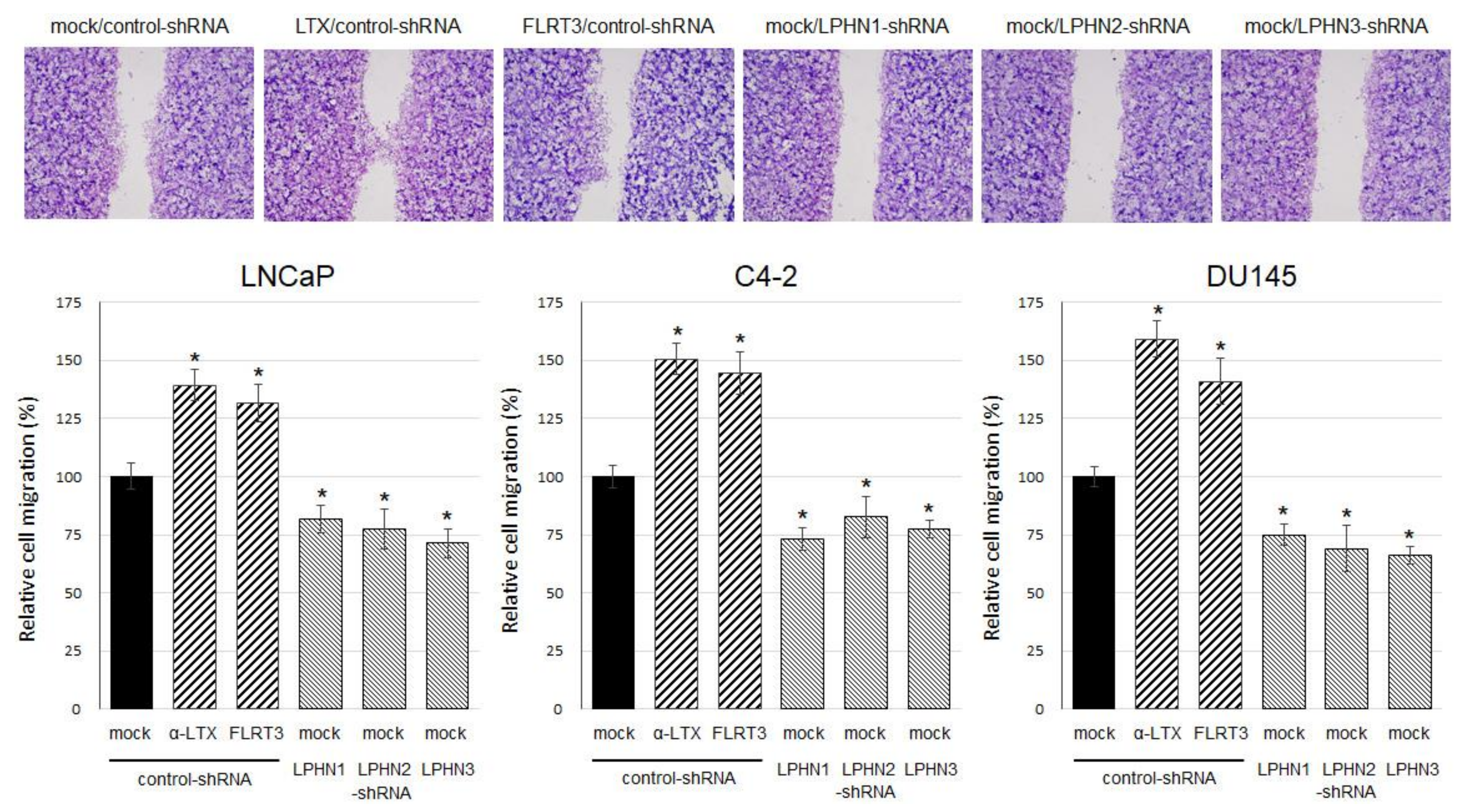
2.2. Effects of LPHN Ligand Treatment or Knockdown on Cell Growth
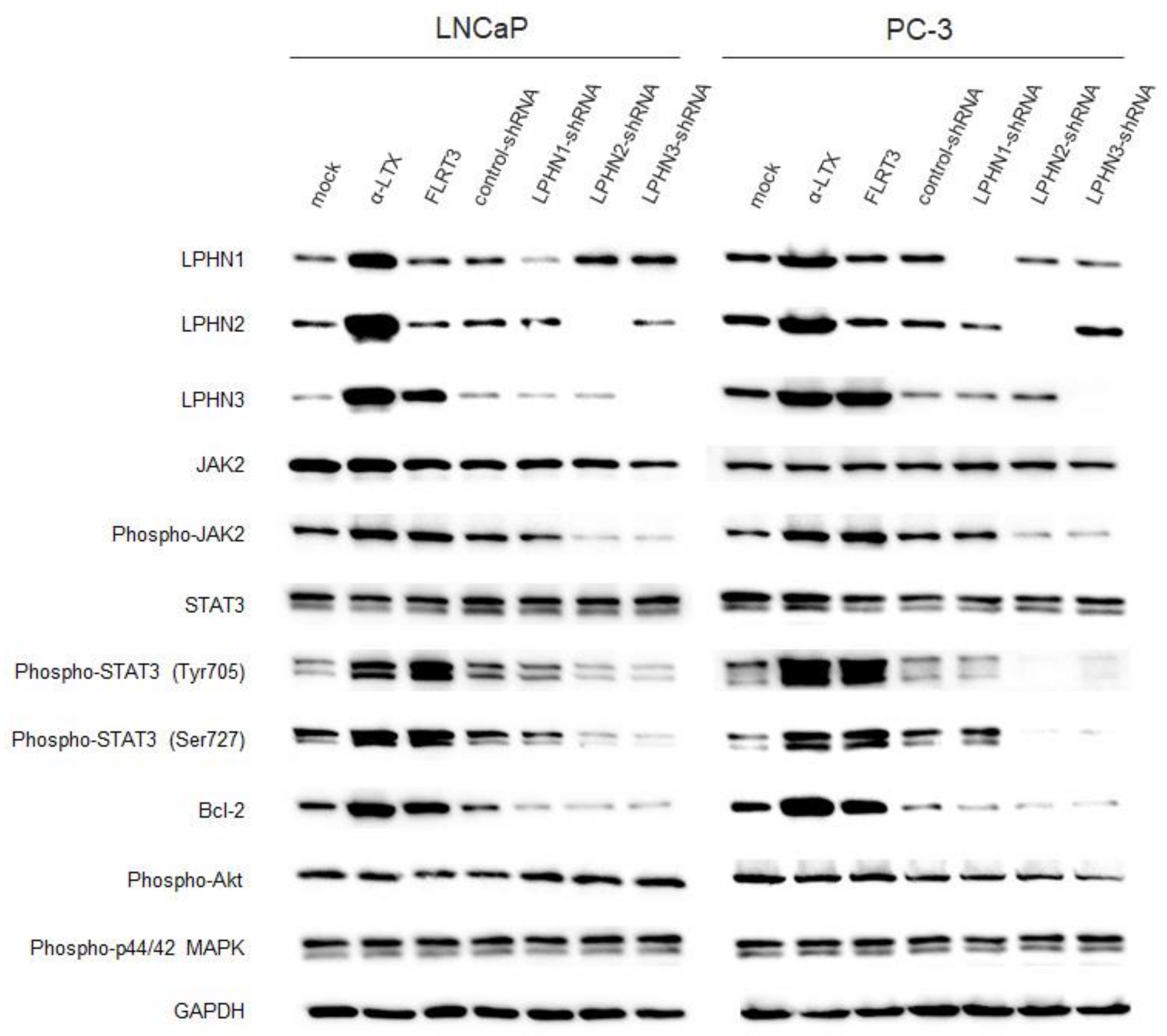
2.3. Involvement of LPHNs in JAK-STAT Signaling
2.4. Expression of LPHNs in Prostate Cancer Specimens
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Chemicals and Antibodies
4.3. Western Blotting
4.4. ChIP Assay
4.5. MTT (3-(4,5-Dimethylthiazol-2-Yl)-2,5Diphenyltetrazolium Bromide) Assay
4.6. Wound-Healing Assay
4.7. Prostate TMA and Immunohistochemistry
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Lancee, M.; Tikkinen, K.A.O.; de Reijke, T.M.; Kataja, V.V.; Aben, K.K.H.; Vernooij, R.W.M. Guideline of guidelines: Primary monotherapies for localised or locally advanced prostate cancer. BJU Int. 2018, 122, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Van den Broeck, T.; van den Bergh, R.C.N.; Briers, E.; Cornford, P.; Cumberbatch, M.; Tilki, D.; De Santis, M.; Fanti, S.; Fossati, N.; Gillessen, S.; et al. Biochemical recurrence in prostate cancer: The European Association of Urology prostate cancer guidelines panel recommendations. Eur. Urol. Focus 2020, 6, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, M.; Lu, C.; Luo, J.; Antonarakis, E.S. AR splicing variants and resistance to AR targeting agents. Cancers 2021, 13, 2563. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ben-David, R.; Mehrazin, R.; Yang, W.; Tewari, A.K.; Kyprianou, N. Novel signatures of prostate cancer progression and therapeutic resistance. Expert Opin. Ther. Targets 2023, 27, 1195–1206. [Google Scholar] [CrossRef]
- Daniels, V.A.; Luo, J.; Paller, C.J.; Kanayama, M. Therapeutic approaches to targeting androgen receptor splice variants. Cells 2024, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Davletov, B.A.; Shamotienko, O.G.; Lelianova, V.G.; Grishin, E.V.; Ushkaryov, Y.A. Isolation and biochemical characterization of a Ca2+-independent α-latrotoxin-binding protein. J. Biol. Chem. 1996, 271, 23239–23245. [Google Scholar] [CrossRef] [PubMed]
- Krasnoperov, V.G.; Beavis, R.; Chepurny, O.G.; Little, A.R.; Plotnikov, A.N.; Petrenko, A.G. The calcium-independent receptor of α-latrotoxin is not a neurexin. Biochem. Biophys. Res. Commun. 1996, 227, 868–875. [Google Scholar] [CrossRef]
- Meza-Aguilar, D.G.; Boucard, A.A. Latrophilins updated. BioMol. Concepts 2014, 5, 457–478. [Google Scholar] [CrossRef]
- Sugita, S.; Ichtchenko, K.; Khvotchev, M.; Südhof, T.C. α-Latrotoxin receptor CIRL/latrophilin 1 (CL1) defined an unusual family of ubiquitous G-protein-linked receptors: G-protein coupling not required for triggering exocytosis. J. Biol. Chem. 1998, 273, 32715–32724. [Google Scholar] [CrossRef] [PubMed]
- Boucard, A.A.; Maxeiner, S.; Südhol, T.C. Latrophilins function as heterophilic cell-adhesion molecules by binding to teneurins. J. Biol. Chem. 2014, 289, 387–402. [Google Scholar] [CrossRef]
- Regan, S.L.; Williams, M.T.; Vorhees, C.V. Review of rodent models of attention Deficit hyperactivity disorder. Neurosci. Biobehav. Rev. 2022, 132, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.V.; Ashwin, E.; Auyeung, B.; Chakrabarti, B.; Lai, M.C.; Taylor, K.; Hackett, G.; Bullmore, E.T.; Baron-Cohen, S. Fetal programming effects of testosterone on the reward system and behavioral approach tendencies in humans. Biol. Psychiatry 2012, 72, 839–847. [Google Scholar] [CrossRef]
- Kosidou, K.; Dalman, C.; Widman, L.; Arver, S.; Lee, B.K.; Magnusson, C.; Gardner, R.M. Maternal polycystic ovary syndrome and risk for attention-deficit/hyperactivity disorder in the offspring. Biol. Psychiatry 2017, 82, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Crowley, F.; Sterpi, M.; Buckley, C.; Margetich, L.; Handa, S.; Dovey, Z. A review of the pathophysiological mechanisms underlying castration-resistant prostate cancer. Res. Rep. Urol. 2021, 13, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat. Rev. Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Qin, K.; Dong, C.; Wu, G.; Lambert, N.A. Inactive-state preassembly of Gq-coupled receptors and Gq heterodimers. Nat. Chem. Biol. 2011, 7, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.S.; Song, S.H.; Yun, J.; Kang, J.Y.; Kim, H.P.; Han, S.W.; Kim, T.Y. Aberrant epigenetic modification of LPHN2 function as a potential cisplatin-specific biomarker for human gastrointestinal cancer. Cancer Res. Treat. 2016, 48, 676–686. [Google Scholar] [CrossRef]
- Kocibalova, Z.; Guzyova, M.; Imrichova, D.; Sulova, Z.; Breier, A. Overexpression of the ABCB1 drug transporter in acute myeloid leukemia cells is associated with downregulation of latrophilin-1. Gen. Physiol. Biophys. 2018, 37, 353–357. [Google Scholar] [CrossRef]
- Yasinska, I.M.; Sakhnevych, S.S.; Pavlova, L.; Teo Hansen Selnø, A.; Teuscher Abeleira, A.M.; Benlaouer, O.; Gonçalves Silva, I.; Mosimann, M.; Varani, L.; Bardelli, M.; et al. The tim-3-galectin-9 pathway and its regulatory mechanisms in human breast cancer. Front. Immunol. 2019, 10, 1594. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xi, S.; Zhao, Q.; Xia, Y.; Yang, Q.; Cai, H.; Wang, F.; Zhao, Y.Y.; Hu, H.J.; Yu, Z.H.; et al. Driver mutations in ADGRL3 are involved in the evolution of ependymoma. Lab. Invest. 2022, 102, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, T.; Jiang, G.; Kawahara, T.; Li, P.; Han, B.; Inoue, S.; Ide, H.; Kato, I.; Jalalizadeh, M.; Miyagi, E.; et al. Androgen receptor signaling reduces the efficacy of bacillus Calmette-Guérin therapy for bladder cancer via modulating Rab27b-induced exocytosis. Mol. Cancer Ther. 2020, 19, 1930–1942. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Yasui, M.; Teramoto, Y.; Nagata, Y.; Mizushima, T.; Miyamoto, H. Latrophilin-3 as a downstream effector of the androgen receptor induces urothelial tumorigenesis. Mol. Carcinogen. 2024, in press. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, M.L.; de Wit, J.; Savas, J.N.; Comoletti, D.; Otto-Hitt, S.; Yates, J.R., III; Ghosh, A. Postsynaptic FLRT proteins are endogenous ligands for the black widow spider venom receptor Latrophilin and regulate excitatory synapse development. Neuron 2012, 73, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Kan, Z.; Jaiswal, B.S.; Stinson, J.; Janakiraman, V.; Bhatt, D.; Stern, H.M.; Yue, P.; Haverty, P.M.; Bourgon, R.; Zheng, J.; et al. Diverse somatic mutation patterns and pathway alterations in human cancers. Nature 2010, 466, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Elahi Najafi, M.A.; Yasui, M.; Teramoto, Y.; Tatenuma, T.; Jiang, G.; Miyamoto, H. GABBR2 as a downstream effector of the androgen receptor induces cisplatin resistance in bladder cancer. Int. J. Mol. Sci. 2023, 24, 13733. [Google Scholar] [CrossRef]
- Canacci, A.M.; Izumi, K.; Zheng, Y.; Gordetsky, J.; Yao, J.L.; Miyamoto, H. Expression of semenogelins I and II and its prognostic significance in human prostate cancer. Prostate 2011, 71, 1108–1114. [Google Scholar] [CrossRef]
- Kawahara, T.; Teramoto, Y.; Li, Y.; Ishiguro, H.; Gordetsky, J.; Yang, Z.; Miyamoto, H. Impact of vasectomy on the development and progression of prostate cancer. Cancers 2020, 12, 2295. [Google Scholar] [CrossRef]
- Izumi, K.; Li, Y.; Zheng, Y.; Gordetsky, J.; Yao, J.L.; Miyamoto, H. Seminal plasma proteins in prostatic carcinoma: Increased nuclear semenogelin I expression is a predictor of biochemical recurrence after radical prostatectomy. Hum. Pathol. 2012, 43, 1991–2000. [Google Scholar] [CrossRef]
- McCarty, K.S., Jr.; Miller, L.S.; Cox, B.; Konrath, J.; McCarthy, K.S., Sr. Estrogen receptor analyses. Correlation of biochemical and immunohistochemical methods using monoclonal antireceptor antibodies. Arch. Pathol. Lab. Med. 1985, 109, 716–721. [Google Scholar] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teramoto, Y.; Elahi Najafi, M.A.; Matsukawa, T.; Sharma, A.; Goto, T.; Miyamoto, H. Latrophilins as Downstream Effectors of Androgen Receptors including a Splice Variant, AR-V7, Induce Prostate Cancer Progression. Int. J. Mol. Sci. 2024, 25, 7289. https://doi.org/10.3390/ijms25137289
Teramoto Y, Elahi Najafi MA, Matsukawa T, Sharma A, Goto T, Miyamoto H. Latrophilins as Downstream Effectors of Androgen Receptors including a Splice Variant, AR-V7, Induce Prostate Cancer Progression. International Journal of Molecular Sciences. 2024; 25(13):7289. https://doi.org/10.3390/ijms25137289
Chicago/Turabian StyleTeramoto, Yuki, Mohammad Amin Elahi Najafi, Takuo Matsukawa, Adhya Sharma, Takuro Goto, and Hiroshi Miyamoto. 2024. "Latrophilins as Downstream Effectors of Androgen Receptors including a Splice Variant, AR-V7, Induce Prostate Cancer Progression" International Journal of Molecular Sciences 25, no. 13: 7289. https://doi.org/10.3390/ijms25137289
APA StyleTeramoto, Y., Elahi Najafi, M. A., Matsukawa, T., Sharma, A., Goto, T., & Miyamoto, H. (2024). Latrophilins as Downstream Effectors of Androgen Receptors including a Splice Variant, AR-V7, Induce Prostate Cancer Progression. International Journal of Molecular Sciences, 25(13), 7289. https://doi.org/10.3390/ijms25137289