
Preparation and Epitope Identification of Monoclonal Antibodies against the NS6 Protein of Porcine Deltacoronavirus (PDCoV)

Abstract
1. Introduction
2. Results
2.1. Plasmid Construction and Protein Expression
2.2. IFA and Western Blot Identification of mAb 5A-11 against PDCoV NS6 Protein
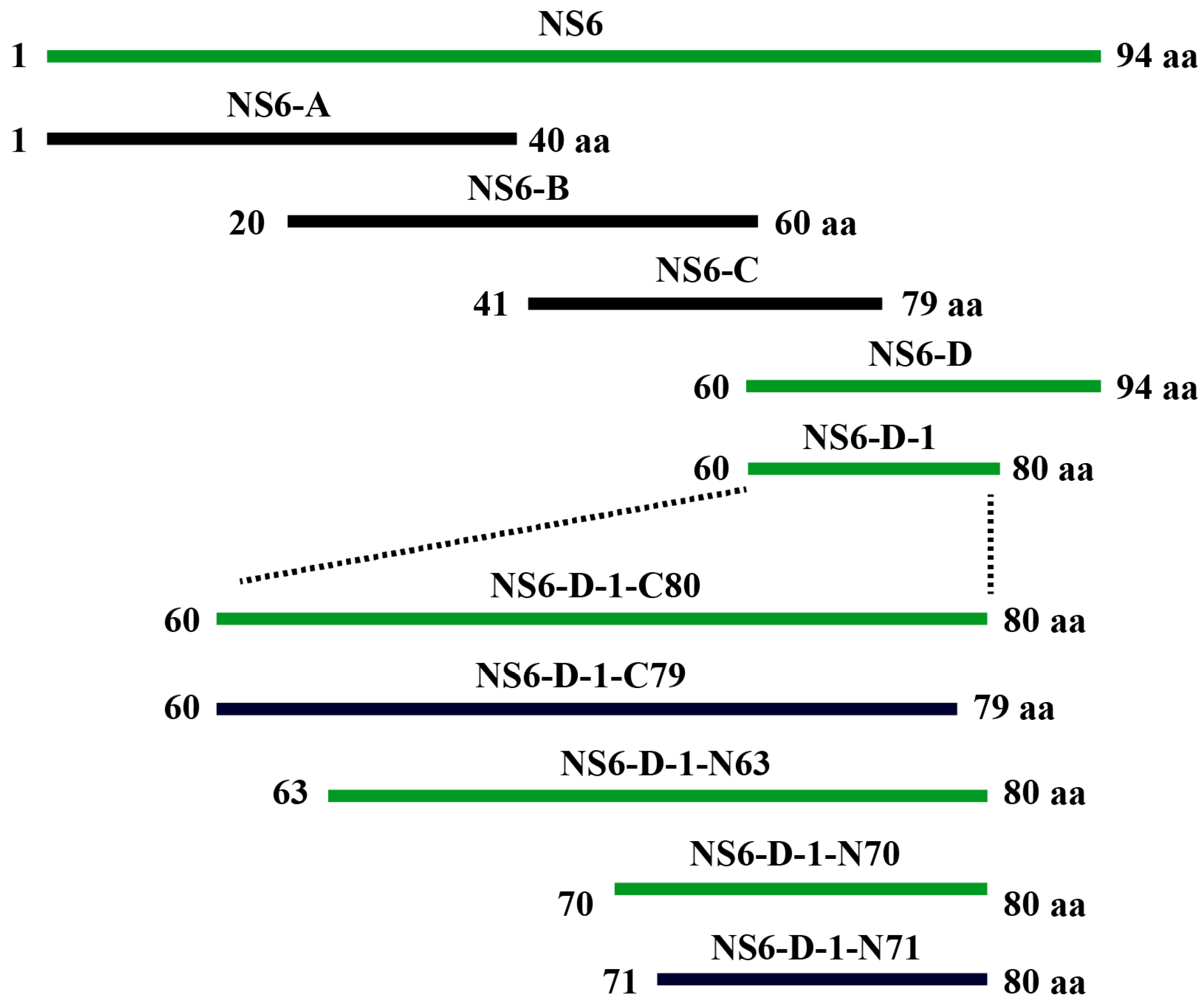
2.3. Epitope Mapping of PDCoV NS6 Protein
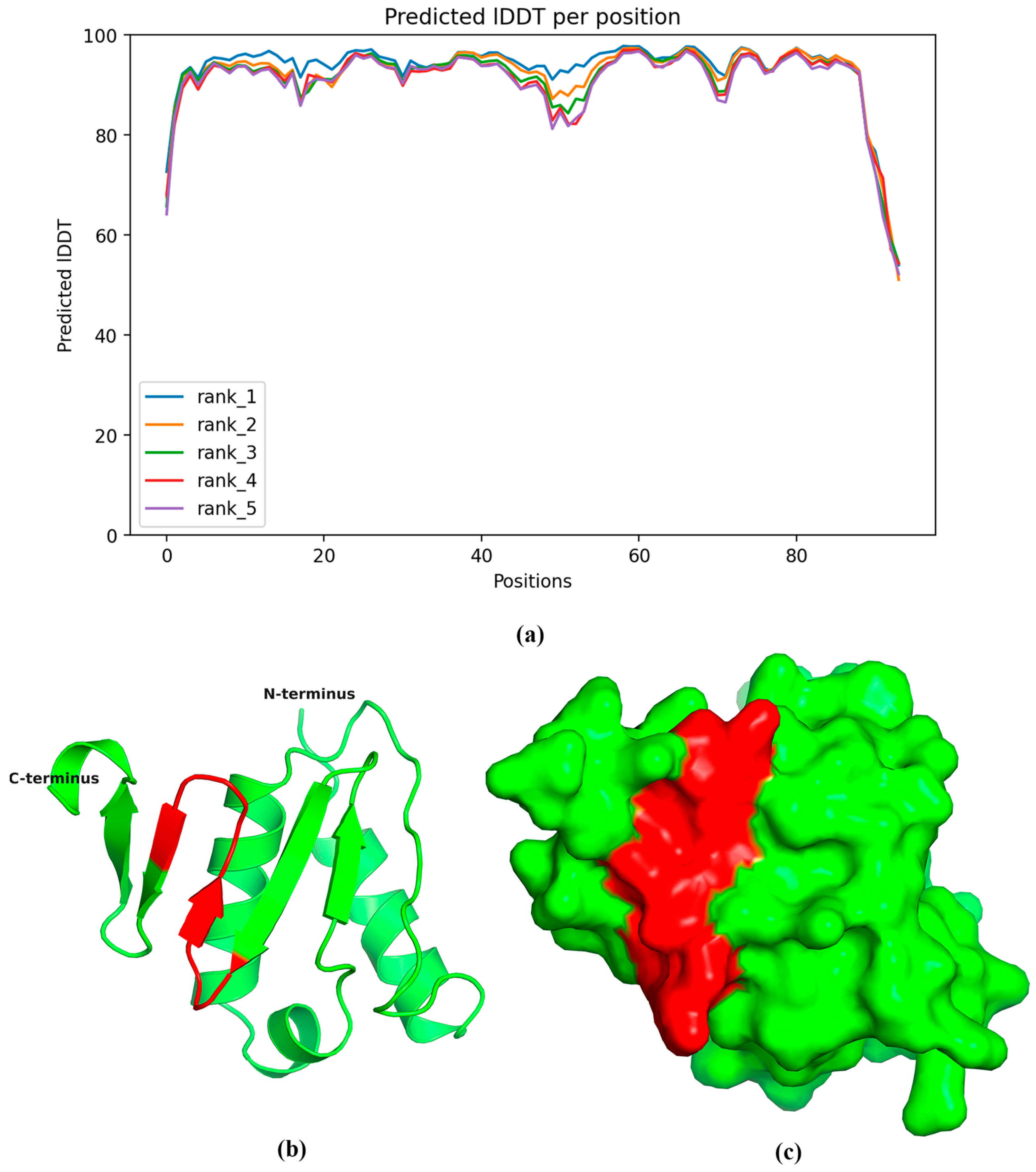
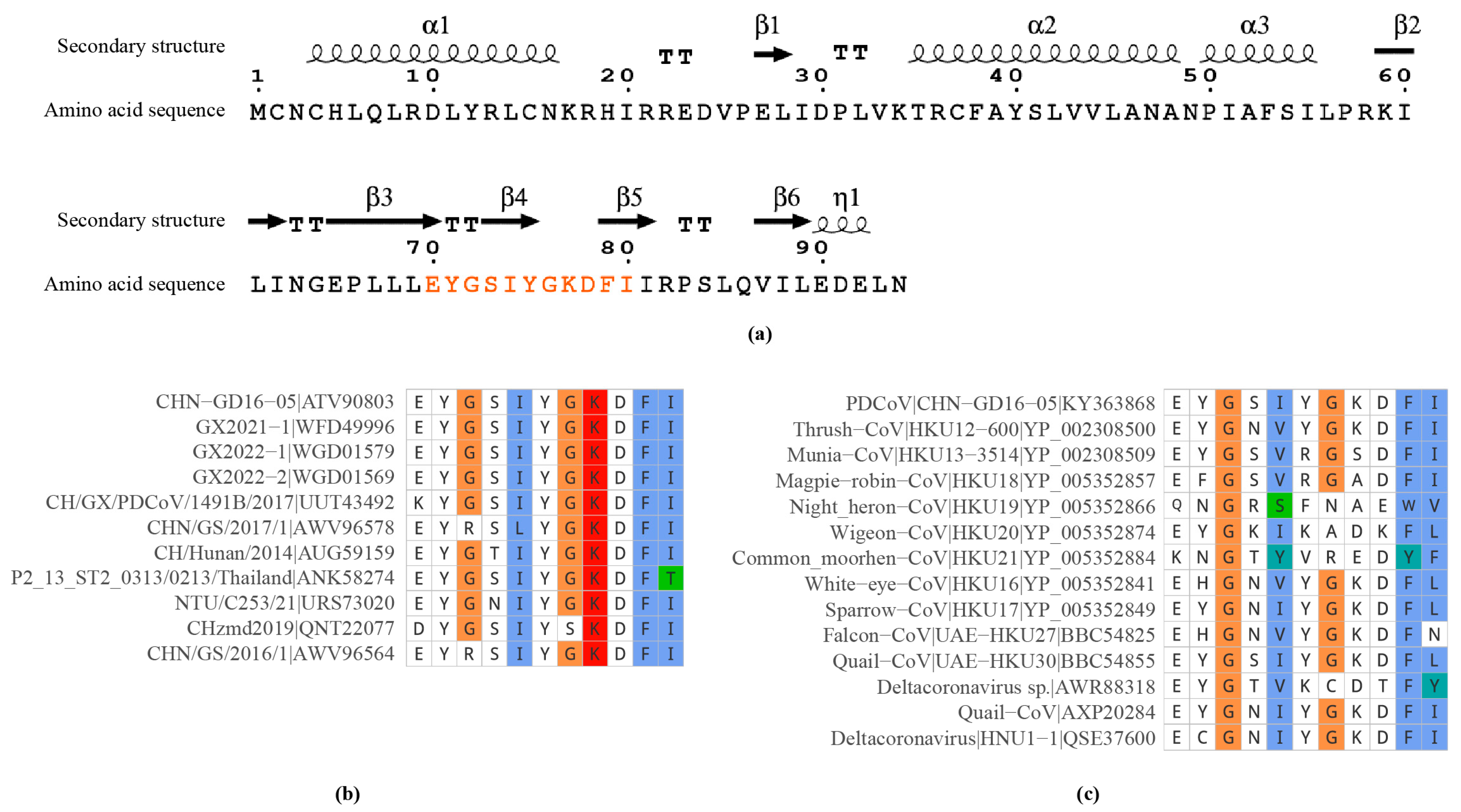
2.4. Spatial Localization and Conservation of the Epitope
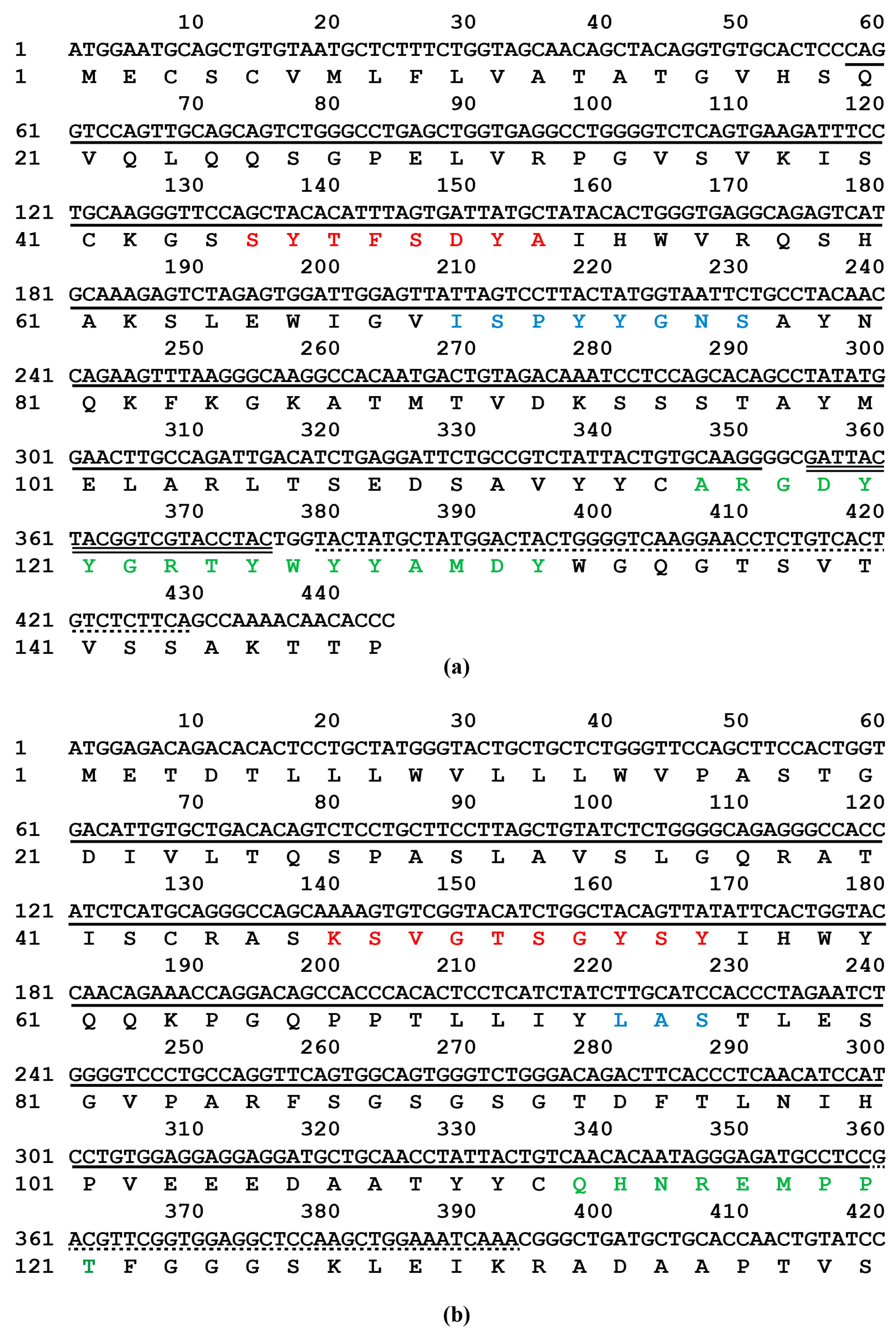
2.5. Sequence Analysis of mAb 5-A11 Light and Heavy Chains
3. Discussion
4. Materials and Methods
4.1. Viruses, Cells, and Animals
4.2. Expression and Purification of PDCoV NS6 Protein
4.3. Generation of mAbs against PDCoV NS6 Protein
4.4. Screening of PDCoV NS6 Protein Antigen
4.5. Identification of Monoclonal Antibodies by Western Blot Analysis
4.6. Mapping the Precise Epitope of PDCoV NS6 Protein
4.7. Prediction of NS6 Protein Structure and Epitope Distribution
4.8. Epitope Sequence Similarity Analysis
4.9. Cloning of the Variable Regions of mAb 5-A11 Heavy and Light Chains
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [PubMed]
- Wang, L.; Byrum, B.; Zhang, Y. Detection and Genetic Characterization of Deltacoronavirus in Pigs, Ohio, USA, 2014. Emerg. Infect. Dis. 2014, 20, 1227. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Raymond, L.; Jiang, Y.; Collins, J.; Rossow, K.; Rovira, A. Rapid Detection, Complete Genome Sequencing, and Phylogenetic Analysis of Porcine Deltacoronavirus. Emerg. Infect. Dis. 2014, 20, 1347. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rivera, C.; Ramírez-Mendoza, H.; Mendoza-Elvira, S.; Segura-Velázquez, R.; Sánchez-Betancourt, J.I. First Report and Phylogenetic Analysis of Porcine Deltacoronavirus in Mexico. Transbound. Emerg. Dis. 2019, 66, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- More-Bayona, J.A.; Ramirez-Velasquez, M.; Hause, B.; Nelson, E.; Rivera-Geronimo, H. First Isolation and Whole Genome Characterization of Porcine Deltacoronavirus from Pigs in Peru. Transbound. Emerg. Dis. 2022, 69, e1561–e1573. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, C. Complete Genome Characterization of Korean Porcine Deltacoronavirus Strain KOR/KNU14-04/2014. Genome Announc. 2014, 2, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Zhou, X.; Peng, Q.; Chen, Y.; Zhang, F.; Huang, T.; Zhang, T.; Li, A.; Huang, D.; Wu, Q. Newly Emerged Porcine Deltacoronavirus Associated with Diarrhoea in Swine in China: Identification, Prevalence and Full-length Genome Sequence Analysis. Transbound. Emerg. Dis. 2015, 62, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Shibahara, T.; Imai, N.; Yamamoto, T.; Ohashi, S. Genetic Characterization and Pathogenicity of Japanese Porcine Deltacoronavirus. Infect. Genet. Evol. 2018, 61, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Janetanakit, T.; Lumyai, M.; Bunpapong, N.; Boonyapisitsopa, S.; Chaiyawong, S.; Nonthabenjawan, N.; Kesdaengsakonwut, S.; Amonsin, A. Porcine Deltacoronavirus, Thailand, 2015. Emerg. Infect. Dis. 2016, 22, 757. [Google Scholar] [CrossRef] [PubMed]
- Lorsirigool, A.; Saeng-Chuto, K.; Temeeyasen, G.; Madapong, A.; Tripipat, T.; Wegner, M.; Tuntituvanont, A.; Intrakamhaeng, M.; Nilubol, D. The First Detection and Full-Length Genome Sequence of Porcine Deltacoronavirus Isolated in Lao PDR. Arch. Virol. 2016, 161, 2909–2911. [Google Scholar] [CrossRef] [PubMed]
- Saeng-Chuto, K.; Lorsirigool, A.; Temeeyasen, G.; Vui, D.; Stott, C.; Madapong, A.; Tripipat, T.; Wegner, M.; Intrakamhaeng, M.; Chongcharoen, W. Different Lineage of Porcine Deltacoronavirus in Thailand, Vietnam and Lao PDR in 2015. Transbound. Emerg. Dis. 2017, 64, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Saif, L.J. Porcine Deltacoronavirus Infection: Etiology, Cell Culture for Virus Isolation and Propagation, Molecular Epidemiology and Pathogenesis. Virus Res. 2016, 226, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Vitosh-Sillman, S.; Loy, J.D.; Brodersen, B.; Kelling, C.; Doster, A.; Topliff, C.; Nelson, E.; Bai, J.; Schirtzinger, E.; Poulsen, E. Experimental Infection of Conventional Nursing Pigs and Their Dams with Porcine Deltacoronavirus. J. Vet. Diagn. Investig. 2016, 28, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Zhang, H.; Li, B.; Ding, Q.; Wang, Y.; Gao, W.; Guo, D.; Wei, Z.; Hu, H. Susceptibility of Chickens to Porcine Deltacoronavirus Infection. Viruses 2019, 11, 573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ding, Q.; Yuan, J.; Han, F.; Wei, Z.; Hu, H. Susceptibility to Mice and Potential Evolutionary Characteristics of Porcine Deltacoronavirus. J. Med. Virol. 2022, 94, 5723–5738. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Saif, L.J. Calves Are Susceptible to Infection with the Newly Emerged Porcine Deltacoronavirus, but Not with the Swine Enteric Alphacoronavirus, Porcine Epidemic Diarrhea Virus. Arch. Virol. 2017, 162, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Hulswit, R.J.; Kenney, S.P.; Widjaja, I.; Jung, K.; Alhamo, M.A.; van Dieren, B.; van Kuppeveld, F.J.; Saif, L.J.; Bosch, B.-J. Broad Receptor Engagement of an Emerging Global Coronavirus May Potentiate Its Diverse Cross-Species Transmissibility. Proc. Natl. Acad. Sci. USA 2018, 115, E5135–E5143. [Google Scholar] [CrossRef] [PubMed]
- Boley, P.A.; Alhamo, M.A.; Lossie, G.; Yadav, K.K.; Vasquez-Lee, M.; Saif, L.J.; Kenney, S.P. Porcine Deltacoronavirus Infection and Transmission in Poultry, United States. Emerg. Infect. Dis. 2020, 26, 255. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, Y.; Ji, C.-M.; Yang, Y.-L.; Liang, Q.-Z.; Zhao, P.; Xu, L.-D.; Lei, X.-M.; Luo, W.-T.; Qin, P. Porcine Deltacoronavirus Engages the Transmissible Gastroenteritis Virus Functional Receptor Porcine Aminopeptidase N for Infectious Cellular Entry. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Stoian, A.; Rowland, R.R.; Petrovan, V.; Sheahan, M.; Samuel, M.S.; Whitworth, K.M.; Wells, K.D.; Zhang, J.; Beaton, B.; Cigan, M. The Use of Cells from ANPEP Knockout Pigs to Evaluate the Role of Aminopeptidase N (APN) as a Receptor for Porcine Deltacoronavirus (PDCoV). Virology 2020, 541, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-L.; Liu, J.; Wang, T.-Y.; Chen, M.; Wang, G.; Yang, Y.-B.; Geng, X.; Sun, M.-X.; Meng, F.; Tang, Y.-D. Aminopeptidase N Is an Entry Co-Factor Triggering Porcine Deltacoronavirus Entry via an Endocytotic Pathway. J. Virol. 2021, 95, 10–1128. [Google Scholar] [CrossRef]
- Xia, S.; Xiao, W.; Zhu, X.; Liao, S.; Guo, J.; Zhou, J.; Xiao, S.; Fang, P.; Fang, L. Porcine Deltacoronavirus Resists Antibody Neutralization through Cell-to-Cell Transmission. Emerg. Microbes Infect. 2023, 12, 2207688. [Google Scholar] [CrossRef] [PubMed]
- Lednicky, J.A.; Tagliamonte, M.S.; White, S.K.; Elbadry, M.A.; Alam, M.M.; Stephenson, C.J.; Bonny, T.S.; Loeb, J.C.; Telisma, T.; Chavannes, S. Independent Infections of Porcine Deltacoronavirus among Haitian Children. Nature 2021, 600, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Liu, X.; Hong, Y.; Wang, Y.; Dong, N.; Ma, P.; Bi, J.; Wang, D.; Xiao, S. Identification and Subcellular Localization of Porcine Deltacoronavirus Accessory Protein NS6. Virology 2016, 499, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Ren, J.; Hong, Y.; Liu, X.; Zhao, Y.; Wang, D.; Peng, G.; Xiao, S. Porcine Deltacoronavirus Accessory Protein NS6 Antagonizes Interferon Beta Production by Interfering with the Binding of RIG-I/MDA5 to Double-Stranded RNA. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Zhang, H.; Cheng, T.; Ding, T.; Xia, S.; Xiao, W.; Li, Z.; Xiao, S.; Fang, L. Porcine Deltacoronavirus Accessory Protein NS6 Harnesses VPS35-Mediated Retrograde Trafficking to Facilitate Efficient Viral Infection. J. Virol. 2023, 97, e00957-23. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Luo, W.-T.; Su, Q.; Zhao, P.; Zhang, Y.; Wang, B.; Yang, Y.-L.; Huang, Y.-W. The Porcine Deltacoronavirus Accessory Protein NS6 Is Expressed in Vivo and Incorporated into Virions. Virology 2021, 556, 1–8. [Google Scholar] [CrossRef]
- Wu, H.; Li, C.; Sun, X.; Cheng, Y.; Chen, Z. Identification of a Monoclonal Antibody against Porcine Deltacoronavirus Membrane Protein. Int. J. Mol. Sci. 2023, 24, 13934. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, J.; Fan, B.; Zhang, X.; Guo, R.; Zhao, Y.; Zhou, J.; Zhou, J.; Sun, D.; Li, B. Development of a Novel Double Antibody Sandwich ELISA for Quantitative Detection of Porcine Deltacoronavirus Antigen. Viruses 2021, 13, 2403. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, L.; Zhang, P.; Ge, X.; Guo, X.; Han, J.; Zhang, Y.; Yang, H. A Strain of Porcine Deltacoronavirus: Genomic Characterization, Pathogenicity and Its Full-length cDNA Infectious Clone. Transbound. Emerg. Dis. 2021, 68, 2130–2146. [Google Scholar] [CrossRef] [PubMed]
- Okda, F.; Lawson, S.; Liu, X.; Singrey, A.; Clement, T.; Hain, K.; Nelson, J.; Christopher-Hennings, J.; Nelson, E.A. Development of Monoclonal Antibodies and Serological Assays Including Indirect ELISA and Fluorescent Microsphere Immunoassays for Diagnosis of Porcine Deltacoronavirus. BMC Vet. Res. 2016, 12, 95. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Shi, D.; Wu, H.; Sun, H.; Chen, J.; Feng, L.; Su, M.; Sun, D. Identification of a Novel B Cell Epitope on the Nucleocapsid Protein of Porcine Deltacoronavirus. Virus Res. 2021, 302, 198497. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Shi, X.; Guan, H.; Zou, Y.; Zhang, S.; Jiang, Z.; Su, S. Identification of a Novel Linear B-Cell Epitope in Porcine Deltacoronavirus Nucleocapsid Protein. Appl. Microbiol. Biotechnol. 2023, 107, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Chen, R.; Hu, J.; Qu, H.; Zhao, Y.; Cao, S.; Wen, X.; Wen, Y.; Wu, R.; Zhao, Q. Identification of a Novel Linear B-Cell Epitope on the Nucleocapsid Protein of Porcine Deltacoronavirus. Int. J. Mol. Sci. 2020, 21, 648. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Yan, X.; Liu, L.; Zhang, Y.; Li, Q.; Li, X.; Hu, H. Identification of Two Novel B-Cell Epitopes on the Nucleocapsid Protein of Porcine Deltacoronavirus. Virol. Sin. 2022, 37, 303. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Luo, S.; Gong, W.; Wang, L.; Ding, N.; Chen, J.; Chen, J.; Wang, T.; Ye, Y.; Song, D. Subcellular Localization of the Porcine Deltacoronavirus Nucleocapsid Protein. Virus Genes 2020, 56, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wen, Y.; Yu, E.; Yang, J.; Liang, Y.; Song, D.; Wen, Y.; Wu, R.; Zhao, Q.; Du, S. Identification of an Immunodominant Neutralizing Epitope of Porcine Deltacoronavirus Spike Protein. Int. J. Biol. Macromol. 2023, 242, 125190. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, J.; Gao, N.; Zhang, X.; Gao, M.; Zhang, X.; Guo, L.; Wu, Y.; Shi, D.; Shi, H. A Novel Antigenic Epitope Identified on the Accessory Protein NS6 of Porcine Deltacoronavirus. Virus Res. 2024, 341, 199329. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Fang, L.; Hong, Y.; Liu, X.; Dong, N.; Ma, P.; Bi, J.; Wang, D.; Xiao, S. Discovery of a Novel Accessory Protein NS7a Encoded by Porcine Deltacoronavirus. J. Gen. Virol. 2017, 98, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, LLC. The PyMOL Molecular Graphics System; Open Source Version 2.5.0; Schrödinger, LLC.: New York, NY, USA, 2021; Available online: https://github.com/schrodinger/pymol-open-source (accessed on 10 May 2021).
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A. The Conserved Domain Database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zhou, L.; Feng, T.; Xu, S.; Gao, F.; Lam, T.T.; Wang, Q.; Wu, T.; Huang, H.; Zhan, L.; Li, L. Ggmsa: A Visual Exploration Tool for Multiple Sequence Alignment and Associated Data. Brief. Bioinform. 2022, 23, bbac222. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 24 April 2024).
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Primer Sequence 1 (5′–3′) | Size (bp) | Purpose of Primers | |
|---|---|---|---|---|
| Sense | Negative Sense | |||
| NS6 protein | TGGGATCCATGTGCAACTGCCATCTGCA | GAGTCGACTTAATTTAATTCATCTTCAAGAATG | 285 | NS6 gene cloning |
| NS6-A | CACCTCGAGATGTGCAACTGCCATCTGCAG | CACAAGCTTGTAAGCAAAACAGCGAGTTTT | 120 | Epitope analysis |
| NS6-B | CACCTCGAGATCAGAAGAGAGGATGTTCCA | CACAAGCTTAATTTTCCGAGGTAGTATGCT | 123 | Epitope analysis |
| NS6-C | CACCTCGAGAGTCTCGTGGTTCTTGCTAAT | CACAAGCTTAAAGTCTTTACCATATATGCT | 117 | Epitope analysis |
| NS6-D | CACCTCGAGATTCTTATCAATGGTGAGCCT | CACAAGCTTTTAATTTAATTCATCTTCAAG | 108 | Epitope analysis |
| NS6-D-1 (NS6-D-1-C80) | CACCTCGAGATTCTTATCAATGGTGAGCCT | CACAAGCTTGATAAAGTCTTTACCATATAT | 63 | Epitope analysis |
| NS6-D-1-C79 | CACCTCGAGATTCTTATCAATGGTGAGCCT | CACAAGCTTAAAGTCTTTACCATATATGCT | 60 | Epitope analysis |
| NS6-D-1-N63 | CACCTCGAGAATGGTGAGCCTTTACTGCTT | CACAAGCTTGATAAAGTCTTTACCATATAT | 54 | Epitope analysis |
| NS6-D-1-N70 | CACCTCGAGGAATATGGTAGCATATATGGT | CACAAGCTTGATAAAGTCTTTACCATATAT | 33 | Epitope analysis |
| NS6-D-1-N71 | CCCTCGAGTATGGTAGCATATATGGTAAAGA | CACAAGCTTGATAAAGTCTTTACCATATAT | 30 | Epitope analysis |
| pEGFP | ATCCGCTAGCGCTACCGGTCGCCA | CAATTTACGCGTTAAGATACATTGATG | Plasmid Verification | |
| VL | ATGGAGACAGACACACTCCTGCTAT | GGATACAGTTGGTGCAGCATCAGCCCGTTT | 420 | Heavy chain cloning |
| VH | ATGGRATGSAGCTGKGTMATSCTCT | GGGSTGTYGTTTTGGCTGMRGAGACRGTGA | 444 | Light chain cloning |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Sun, X.; Li, C.; Xie, S.; Chen, Z. Preparation and Epitope Identification of Monoclonal Antibodies against the NS6 Protein of Porcine Deltacoronavirus (PDCoV). Int. J. Mol. Sci. 2024, 25, 7645. https://doi.org/10.3390/ijms25147645
Wu H, Sun X, Li C, Xie S, Chen Z. Preparation and Epitope Identification of Monoclonal Antibodies against the NS6 Protein of Porcine Deltacoronavirus (PDCoV). International Journal of Molecular Sciences. 2024; 25(14):7645. https://doi.org/10.3390/ijms25147645
Chicago/Turabian StyleWu, Huiguang, Xian Sun, Chen Li, Sihan Xie, and Zhenhai Chen. 2024. "Preparation and Epitope Identification of Monoclonal Antibodies against the NS6 Protein of Porcine Deltacoronavirus (PDCoV)" International Journal of Molecular Sciences 25, no. 14: 7645. https://doi.org/10.3390/ijms25147645
APA StyleWu, H., Sun, X., Li, C., Xie, S., & Chen, Z. (2024). Preparation and Epitope Identification of Monoclonal Antibodies against the NS6 Protein of Porcine Deltacoronavirus (PDCoV). International Journal of Molecular Sciences, 25(14), 7645. https://doi.org/10.3390/ijms25147645