


Cosmic Ionizing Radiation: A DNA Damaging Agent That May Underly Excess Cancer in Flight Crews

 ,
,
Abstract

1. Introduction
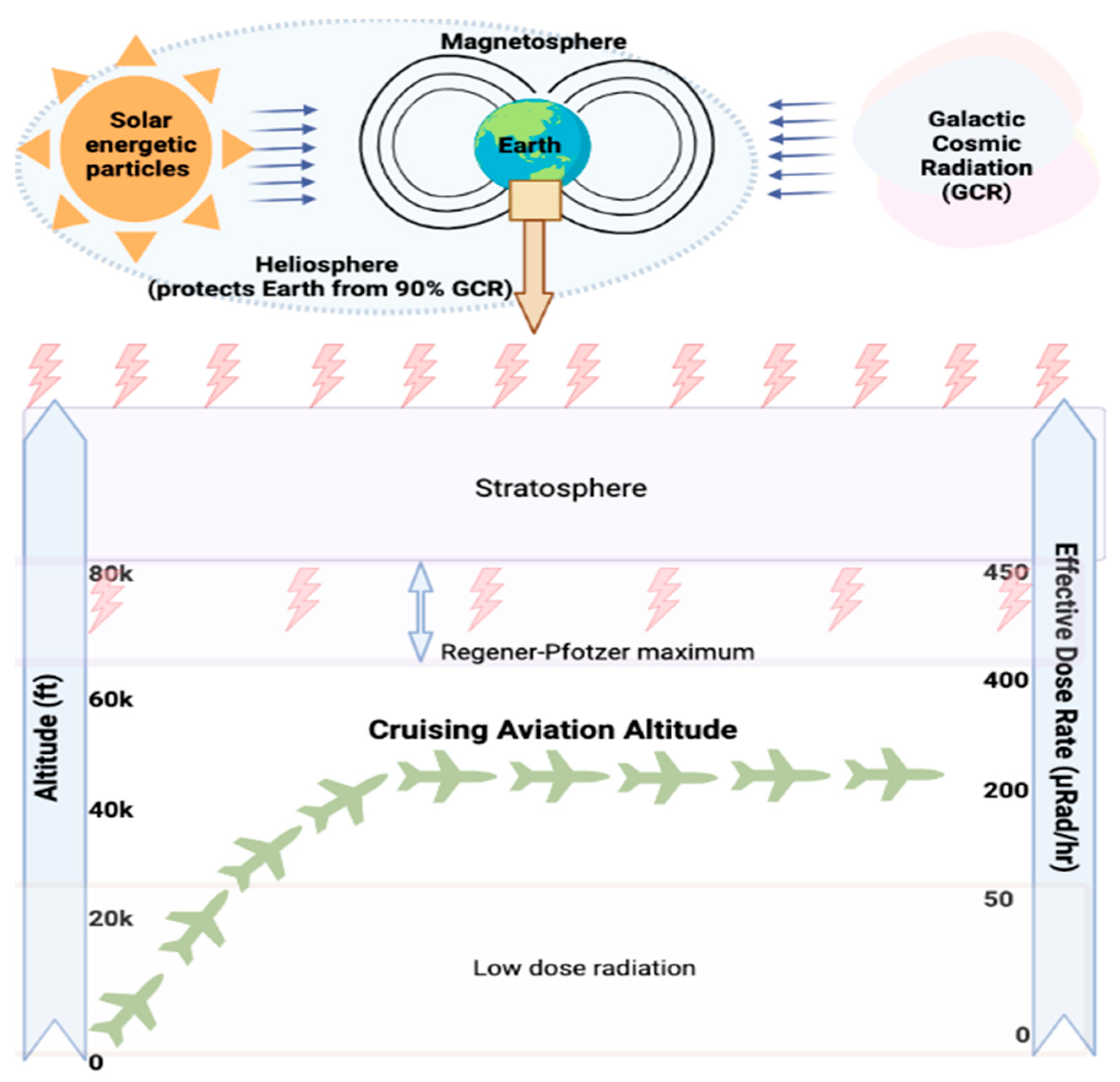
2. Current Understanding of Exposure to CIR during Flight Travel
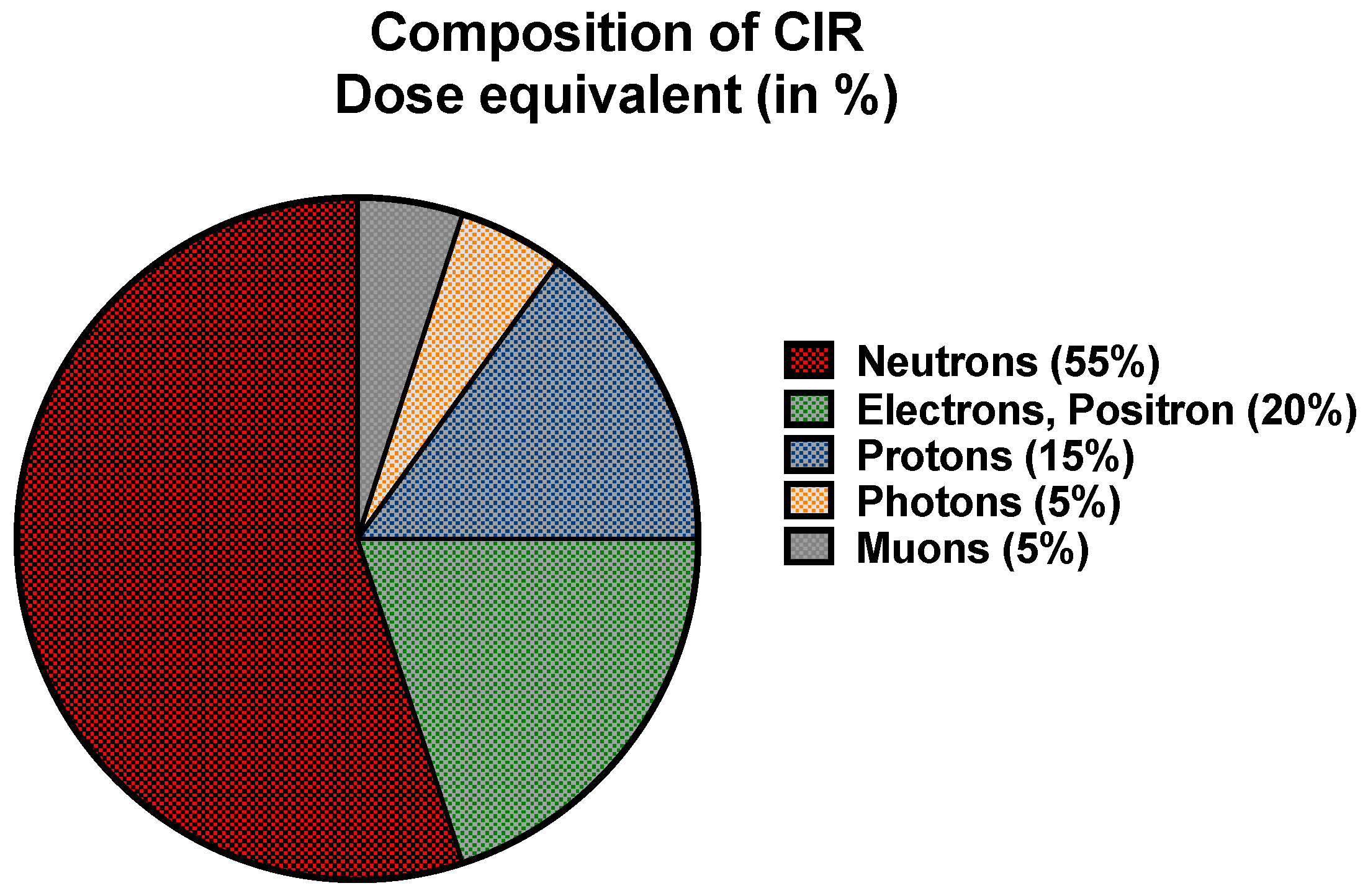
2.1. Exposure to CIR during Flight
2.2. Methods for Measuring or Estimating Exposure to CIR during Air Travel
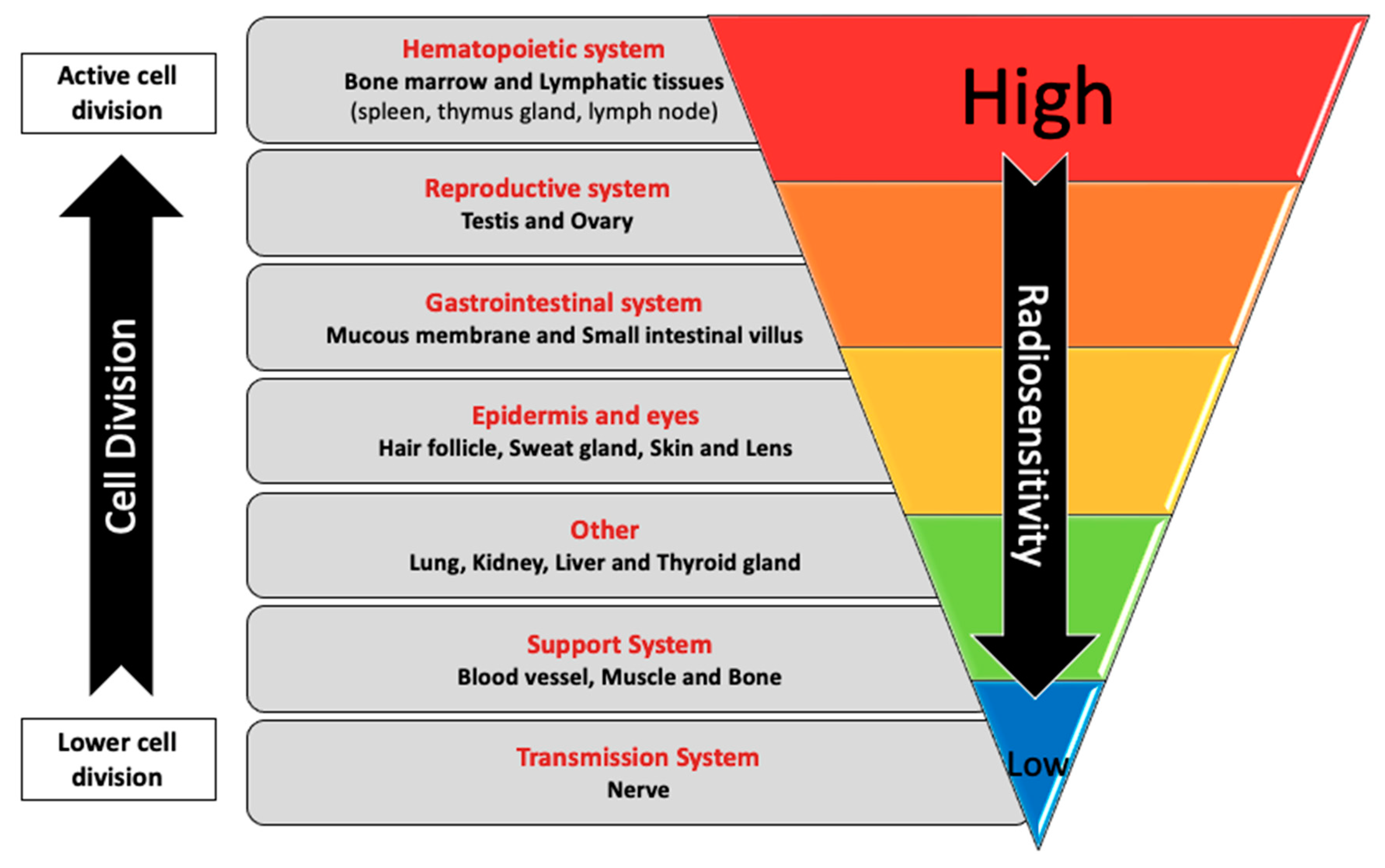
3. Interindividual Variation in Response to Exposure to Radiation
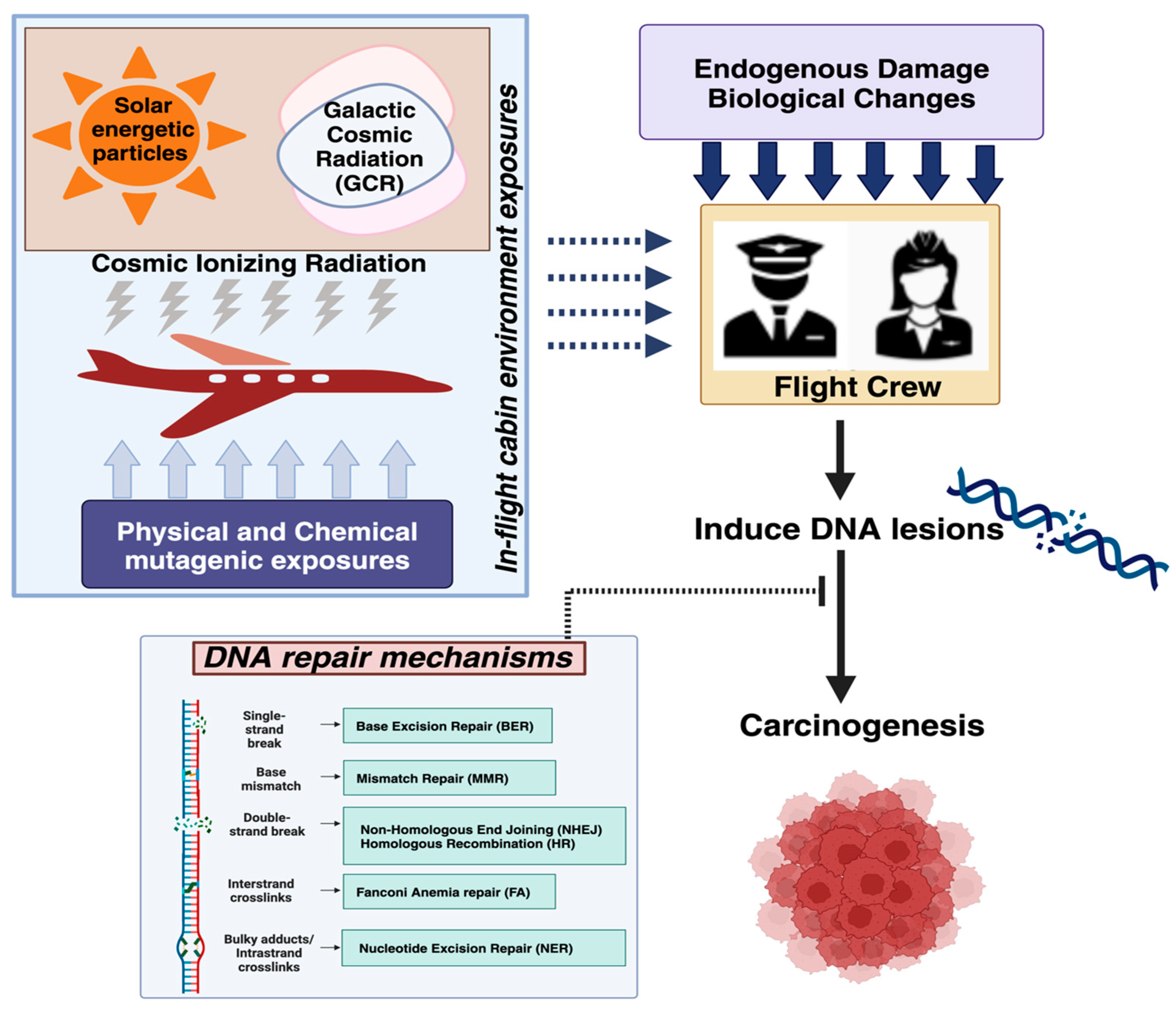
4. DNA Damage and Repair Mechanisms Associated with Exposure to CIR in FC
4.1. Markers of DNA Damage and Genomic Instability in FC That Are Consistent with the Expected Biological Effects of Exposure to CIR
4.2. DNA Repair Mechanisms Involved in the Cellular Response to the Damage Induced by Components of CIR
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IATA. IATA—World Air Transport Statistics (WATS). Available online: https://www.iata.org/en/services/data/market-data/world-air-transport-statistics/ (accessed on 14 June 2024).
- CAPA. The Global Pilot Shortage Is a Challenge to the World’s Airlines. 2023. Available online: https://centreforaviation.com/analysis/reports/the-global-pilot-shortage-is-a-challenge-to-the-worlds-airlines-658033 (accessed on 8 May 2024).
- DataUSA. IO Flight Attendants|Data USA. Available online: https://datausa.io/profile/soc/flight-attendants (accessed on 14 June 2024).
- National Council on Radiation Protection and Measurements. Ionizing Radiation Exposure of the Population of the United States; Report No. 160; Council on Radiation Protection and Measurements (NCRP): Bethesda, MD, USA, 2009. [Google Scholar]
- Lin, E.C. Radiation Risk From Medical Imaging. Mayo Clin. Proc. 2010, 85, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- UNSCEAR; Annex, C. Biological Mechanisms Relevant for the Inference of Cancer Risks from Low-Dose and Low-Dose-Rateradiation; UNSCEAR 2020/2021 Report; UNSCEAR: Vienna, Austria, 2021. [Google Scholar]
- Aw, J. Cosmic Radiation and Commercial Air Travel. J. Travel. Med. 2003, 10, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Goldhagen, P. Overview of Aircraft Radiation Exposure and Recent ER-2 Measurements. Health Phys. 2000, 79, 526–544. [Google Scholar] [CrossRef] [PubMed]
- Lantos, P.; Fuller, N. History of the Solar Particle Event Radiation Doses On-Board Aeroplanes Using a Semi-Empirical Model and Concorde Measurements. Radiat. Prot. Dosim. 2003, 104, 199–210. [Google Scholar] [CrossRef] [PubMed]
- McNeely, E.; Gale, S.; Tager, I.; Kincl, L.; Bradley, J.; Coull, B.; Hecker, S. The Self-Reported Health of U.S. Flight Attendants Compared to the General Population. Environ. Health 2014, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Scheibler, C.; Toprani, S.M.; Mordukhovich, I.; Schaefer, M.; Staffa, S.; Nagel, Z.D.; McNeely, E. Cancer Risks from Cosmic Radiation Exposure in Flight: A Review. Front. Public Health 2022, 10, 947068. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.F.; Powell, D.M.C. The Occupational Health and Safety of Flight Attendants. Aviat. Space Environ. Med. 2012, 83, 514–521. [Google Scholar] [CrossRef] [PubMed]
- McNeely, E.; Mordukhovich, I.; Tideman, S.; Gale, S.; Coull, B. Estimating the Health Consequences of Flight Attendant Work: Comparing Flight Attendant Health to the General Population in a Cross-Sectional Study. BMC Public Health 2018, 18, 346. [Google Scholar] [CrossRef]
- Bramlitt, E.T.; Shonka, J.J. Radiation Exposure of Aviation Crewmembers and Cancer. Health Phys. 2015, 108, 76–86. [Google Scholar] [CrossRef]
- Toprani, S.M.; Scheibler, C.; Nagel, Z.D. Interplay Between Air Travel, Genome Integrity, and COVID-19 Risk Vis-a-Vis Flight Crew. Front. Public Health 2020, 8, 590412. [Google Scholar] [CrossRef]
- McNeely, E.; Mordukhovich, I.; Staffa, S.; Tideman, S.; Coull, B. Legacy Health Effects among Never Smokers Exposed to Occupational Secondhand Smoke. PLoS ONE 2019, 14, e0215445. [Google Scholar] [CrossRef]
- Pinkerton, L.E.; Hein, M.J.; Anderson, J.L.; Little, M.P.; Sigurdson, A.J.; Schubauer-Berigan, M.K. Breast Cancer Incidence among Female Flight Attendants: Exposure–Response Analyses. Scand. J. Work Environ. Health 2016, 42, 538–546. [Google Scholar] [CrossRef] [PubMed]
- McNeely, E.; Mordukhovich, I.; Staffa, S.; Tideman, S.; Gale, S.; Coull, B. Cancer Prevalence among Flight Attendants Compared to the General Population. Environ. Health 2018, 17, 49. [Google Scholar] [CrossRef] [PubMed]
- Pukkala, E.; Helminen, M.; Haldorsen, T.; Hammar, N.; Kojo, K.; Linnersjö, A.; Rafnsson, V.; Tulinius, H.; Tveten, U.; Auvinen, A. Cancer Incidence among Nordic Airline Cabin Crew. Int. J. Cancer 2012, 131, 2886–2897. [Google Scholar] [CrossRef] [PubMed]
- Pinkerton, L.E.; Hein, M.J.; Anderson, J.L.; Christianson, A.; Little, M.P.; Sigurdson, A.J.; Schubauer-Berigan, M.K. Melanoma, Thyroid Cancer, and Gynecologic Cancers in a Cohort of Female Flight Attendants. Am. J. Ind. Med. 2018, 61, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Schüz, J. Airline Crew Cohorts: Is There More to Learn Regarding Their Cancer Risk? Occup. Environ. Med. 2014, 71, 307. [Google Scholar] [CrossRef] [PubMed]
- Nuclear Regulatory Commission (NRC). Biological Effects of Radiation. U.S. Nuclear Regulatory Commission Backgrounder; Nuclear Regulatory Commission: Rockville, MD, USA, 2017. [Google Scholar]
- ICRP International Commission on Radiological Protection (ICRP). Publication 103: Recommendations of the International Commission on Radiological Protection. Radiat. Prot. Dosim. 2008, 129, 500–507. [Google Scholar]
- Sridharan, D.M.; Asaithamby, A.; Blattnig, S.R.; Costes, S.V.; Doetsch, P.W.; Dynan, W.S.; Hahnfeldt, P.; Hlatky, L.; Kidane, Y.; Kronenberg, A.; et al. Evaluating Biomarkers to Model Cancer Risk Post Cosmic Ray Exposure. Life Sci. Space Res. 2016, 9, 19–47. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Barnes, D.E. Repair of Endogenous DNA Damage. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 127–134. [Google Scholar] [CrossRef]
- Hall, E.J.; Giaccia, A.J. Radiobiology for the Radiologist, 7th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012; ISBN 1608311937. [Google Scholar]
- Lomax, M.E.; Folkes, L.K.; O’Neill, P. Biological Consequences of Radiation-Induced DNA Damage: Relevance to Radiotherapy. Clin. Oncol. (R. Coll. Radiol.) 2013, 25, 578–585. [Google Scholar] [CrossRef]
- Jackson, S.P.; Bartek, J. The DNA-Damage Response in Human Biology and Disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Nagel, Z.D.; Chaim, I.A.; Samson, L.D. Inter-Individual Variation in DNA Repair Capacity: A Need for Multi-Pathway Functional Assays to Promote Translational DNA Repair Research. DNA Repair 2014, 19, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.C.; Dizdaroglu, M. Implications of DNA Damage and DNA Repair on Human Diseases. Mutagenesis 2020, 35, 1–3. [Google Scholar] [CrossRef]
- Yoo, S.S.; Jorgensen, T.J.; Kennedy, A.R.; Boice, J.D.; Shapiro, A.; Hu, T.C.C.; Moyer, B.R.; Grace, M.B.; Kelloff, G.J.; Fenech, M.; et al. Mitigating the Risk of Radiation-Induced Cancers: Limitations and Paradigms in Drug Development. J. Radiol. Prot. 2014, 34, R25. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.T. Radiation Protection Aspects of the Cosmic Radiation Exposure of Aircraft Crew. Radiat. Prot. Dosim. 2004, 109, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Vuković, B.; Radolić, V.; Miklavčić, I.; Poje, M.; Varga, M.; Planinić, J. Cosmic Radiation Dose in Aircraft--a Neutron Track Etch Detector. J. Environ. Radioact. 2007, 98, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Kim, S.H. Fabrication of a Gd2O3-Based Film to Shield from Space Radiation inside Aircraft and Its Effectiveness. Aerospace 2023, 10, 968. [Google Scholar] [CrossRef]
- Lochard, J.; Bartlett, D.T.; Rühm, W.; Yasuda, H.; Bottollier-Depois, J.F. ICRP Publication 132: Radiological Protection from Cosmic Radiation in Aviation. Ann. ICRP 2016, 45, 5–48. [Google Scholar] [CrossRef]
- Prado, A.C.M.; Federico, C.A.; Pereira Junior, E.C.F.; Goncalez, O.L. Effects of Cosmic Radiation on Devices and Embedded Systems in Aircrafts. In Proceedings of the 2013 International Nuclear Atlantic Conference-INAC, Recife, Brazil, 24–29 November 2013. [Google Scholar]
- Silva, R.; Folgosa, F.; Soares, P.; Pereira, A.S.; Garcia, R.; Gestal-Otero, J.J.; Tavares, P.; Gomes Da Silva, M.D.R. Occupational Cosmic Radiation Exposure in Portuguese Airline Pilots: Study of a Possible Correlation with Oxidative Biological Markers. Radiat. Environ. Biophys. 2013, 52, 211–220. [Google Scholar] [CrossRef]
- Mishev, A. Short- and Medium-Term Induced Ionization in the Earth Atmosphere by Galactic and Solar Cosmic Rays. Int. J. Atmos. Sci. 2013, 2013, 184508. [Google Scholar] [CrossRef]
- Usoskin, I.G.; Desorgher, L.; Velinov, P.; Storini, M.; Flückiger, E.O.; Bütikofer, R.; Kovaltsov, G.A. Ionization of the Earth’s Atmosphere by Solar and Galactic Cosmic Rays. Acta Geophys. 2008, 57, 88–101. [Google Scholar] [CrossRef]
- Phillips, T.; Johnson, S.; Koske-Phillips, A.; White, M.; Yarborough, A.; Lamb, A.; Herbst, A.; Molina, F.; Gilpin, J.; Grah, O.; et al. Space Weather Ballooning. Space Weather 2016, 14, 697–703. [Google Scholar] [CrossRef]
- Hands, A.D.P.; Ryden, K.A.; Mertens, C.J. The Disappearance of the Pfotzer-Regener Maximum in Dose Equivalent Measurements in the Stratosphere. Space Weather 2016, 14, 776–785. [Google Scholar] [CrossRef]
- Meier, M.M.; Copeland, K.; Klöble, K.E.J.; Matthiä, D.; Plettenberg, M.C.; Schennetten, K.; Wirtz, M.; Hellweg, C.E. Radiation in the Atmosphere—A Hazard to Aviation Safety? Atmosphere 2020, 11, 1358. [Google Scholar] [CrossRef]
- EURADOS. Comparison of Codes Assessing Radiation Exposure of Aircraft Crew Due to Galactic Cosmic Radiation; EURADOS Report 2012-03; EURADOS: Braunschweig, Germany, 2012. [Google Scholar]
- EURADOS. Exposure of Air Crew to Cosmic Radiation: A Report of EURADOS Working Group 11; EURADOS Report; EURADOS: Luxembourg, 1996. [Google Scholar]
- Federal Aviation Administration (FAA). In-Flight Radiation Exposure; FAA Advisory Circular 120–61B; US Department of Transportation: Washington, DC, USA, 2014. [Google Scholar]
- Desmaris, G. Cosmic Radiation in Aviation: Radiological Protection of Air France Aircraft Crew. Ann. ICRP 2016, 45, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Blettner, M.; Grosche, B.; Zeeb, H. Occupational Cancer Risk in Pilots and Flight Attendants: Current Epidemiological Knowledge. Radiat. Environ. Biophys. 1998, 37, 75–80. [Google Scholar] [CrossRef]
- Friedberg, W.; Faulkner, D.N.; Snyder, L.; Darden, E.B., Jr.; O’Brien, K. Galactic Cosmic Radiation Exposure and Associated Health Risks for Air Carrier Crewmembers. Aviat. Space Environ. Med. 1989, 60, 1104–1108. [Google Scholar]
- Dreger, S.; Wollschläger, D.; Schafft, T.; Hammer, G.P.; Blettner, M.; Zeeb, H. Cohort Study of Occupational Cosmic Radiation Dose and Cancer Mortality in German Aircrew, 1960–2014. Occup. Environ. Med. 2020, 77, 285–291. [Google Scholar] [CrossRef]
- Bottollier-Depois, J.F.; Chau, Q.; Bouisset, P.; Kerlau, G.; Plawinski, L.; Lebaron-Jacobs, L. Assessing Exposure to Cosmic Radiation on Board Aircraft. Adv. Space Res. 2003, 32, 59–66. [Google Scholar] [CrossRef]
- Oksanen, P.J. Estimated Individual Annual Cosmic Radiation Doses for Flight Crews. Aviat. Space Environ. Med. 1998, 69, 621–625. [Google Scholar]
- Grajewski, B.; Yong, L.C.; Bertke, S.J.; Bhatti, P.; Little, M.P.; Ramsey, M.J.; Tucker, J.D.; Ward, E.M.; Whelan, E.A.; Sigurdson, A.J.; et al. Chromosome Translocations and Cosmic Radiation Dose in Male U.S. Commercial Airline Pilots. Aerosp. Med. Hum. Perform. 2018, 89, 616–625. [Google Scholar] [CrossRef]
- Toprani, S.M.S.M.; Das, B. Radio-Adaptive Response of Base Excision Repair Genes and Proteins in Human Peripheral Blood Mononuclear Cells Exposed to Gamma Radiation. Mutagenesis 2015, 30, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.; Koliada, A.; Zabuga, O.; Socol, Y. Health Impacts of Low-Dose Ionizing Radiation: Current Scientific Debates and Regulatory Issues. Dose Response 2018, 16, 1559325818796331. [Google Scholar] [CrossRef]
- Toprani, S.M.; Das, B. Role of Base Excision Repair Genes and Proteins in Gamma-Irradiated Resting Human Peripheral Blood Mononuclear Cells. Mutagenesis 2015, 30, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J. The Additive to Background Assumption in Cancer Risk Assessment: A Reappraisal. Environ. Res. 2018, 166, 175–204. [Google Scholar] [CrossRef]
- Toprani, S.M.; Das, B. Radio-Adaptive Response, Individual Radio-Sensitivity and Correlation of Base Excision Repair Gene Polymorphism (HOGG1, APE1, XRCC1, and LIGASE1) in Human Peripheral Blood Mononuclear Cells Exposed to Gamma Radiation. Environ. Mol. Mutagen. 2020, 61, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Komov, O.; Krasavin, E.; Nasonov, E.; Mel’nikov, L.; Shmakov, N.; Cunh, M.; Testa, E.; Beuve, M. Relationship between Radioadaptive Response and Individual Radiosensitivity to Low Doses of Gamma Radiation: An Extended Study of Chromosome Damage in Blood Lymphocytes of Three Donors. Int. J. Radiat. Biol. 2018, 94, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Nenoi, M.; Wang, B.; Vares, G. In Vivo Radioadaptive Response: A Review of Studies Relevant to Radiation-Induced Cancer Risk. Hum. Exp. Toxicol. 2015, 34, 272. [Google Scholar] [CrossRef]
- Bolzán, A.D.; Bianchi, M.S.; Giménez, E.M.; Flaqué, M.C.D.; Ciancio, V.R. Analysis of Spontaneous and Bleomycin-Induced Chromosome Damage in Peripheral Lymphocytes of Long-Haul Aircrew Members from Argentina. Mutat. Res. 2008, 639, 64–79. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Radiation Emergencies Factsheet—Radiation and Pregnancy: A Fact Sheet for the Public Department of Health and Human Services; Updated 15 November 2011; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2011. [Google Scholar]
- Williams, P.; Fletcher, S. Health Effects of Prenatal Radiation Exposure. Am. Fam. Physician 2011, 82, 488–493. [Google Scholar]
- United Nations; Scientific Committee on the Effects of Atomic Radiation. General Assembly. Genetic and Somatic Effects of Ionizing Radiation: United Nations Scientific Committee on the Effects of Atomic Radiation: 1986 Report to the General Assembly, with Annexes; United Nations: New York, NY, USA, 1986; p. 366. [Google Scholar]
- Valentin, J.; Cox, R.; Streffer, C. Biological Effects after Prenatal Irradiation (Embryo and Fetus): ICRP Publication 90 Approved by the Commission in October 2002. Ann. ICRP 2003, 33, 1–206. [Google Scholar] [CrossRef] [PubMed]
- Saada, M.; Sanchez-Jimenez, E.; Roguin, A. Risk of Ionizing Radiation in Pregnancy: Just a Myth or a Real Concern? Europace 2023, 25, 270. [Google Scholar] [CrossRef]
- Grajewski, B.; Whelan, E.A.; Lawson, C.C.; Hein, M.J.; Waters, M.A.; Anderson, J.L.; Macdonald, L.A.; Mertens, C.J.; Tseng, C.Y.; Cassinelli, R.T.; et al. Miscarriage among Flight Attendants. Epidemiology 2015, 26, 192–203. [Google Scholar] [CrossRef]
- Cone, J.E.; Vaughan, L.M.; Huete, A.; Samuels, S.J. Reproductive Health Outcomes among Female Flight Attendants: An Exploratory Study. J. Occup. Environ. Med. 1998, 40, 210–216. [Google Scholar] [CrossRef]
- Lauria, L.; Ballard, T.J.; Caldora, M.; Mazzanti, C.; Verdecchia, A. Reproductive Disorders and Pregnancy Outcomes among Female Flight Attendants. Aviat. Space Environ. Med. 2006, 77, 533–539. [Google Scholar] [PubMed]
- Irgens, Å.; Irgens, L.M.; Reitan, J.B.; Haldorsen, T.; Tveten, U. Pregnancy Outcome among Offspring of Airline Pilots and Cabin Attendants. Scand. J. Work Environ. Health 2003, 29, 94–99. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, W.; Chan, A.; Li, C.; He, X.; Cui, L.; Lv, Y.; Liu, J.; Guo, X. An Epidemiological Study of Reproductive Health in Female Civil Aviation Employees. Aviat. Space Environ. Med. 2013, 84, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, A.R.; Barnes, J.; Ejiogu, N.; Foster, K.; Brant, L.J.; Zonderman, A.B.; Evans, M.K. Age, Sex, and Race Influence Single-Strand Break Repair Capacity in a Human Population. Free Radic. Biol. Med. 2008, 45, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Garm, C.; Moreno-Villanueva, M.; Bürkle, A.; Petersen, I.; Bohr, V.A.; Christensen, K.; Stevnsner, T. Age and Gender Effects on DNA Strand Break Repair in Peripheral Blood Mononuclear Cells. Aging Cell 2013, 12, 58–66. [Google Scholar] [CrossRef]
- Scott, D.; Barber, J.B.P.; Levine, E.L.; Burrill, W.; Roberts, S.A. Radiation-Induced Micronucleus Induction in Lymphocytes Identifies a High Frequency of Radiosensitive Cases among Breast Cancer Patients: A Test for Predisposition? Br. J. Cancer 1998, 77, 614–620. [Google Scholar] [CrossRef]
- Distel, L.V.R.; Neubauer, S.; Keller, U.; Sprung, C.N.; Sauer, R.; Grabenbauer, G.G. Individual Differences in Chromosomal Aberrations after in Vitro Irradiation of Cells from Healthy Individuals, Cancer and Cancer Susceptibility Syndrome Patients. Radiother. Oncol. 2006, 81, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Köberle, B.; Koch, B.; Fischer, B.M.; Hartwig, A. Single Nucleotide Polymorphisms in DNA Repair Genes and Putative Cancer Risk. Arch. Toxicol. 2016, 90, 2369–2388. [Google Scholar] [CrossRef] [PubMed]
- Heimers, A. Chromosome Aberration Analysis in Concorde Pilots. Mutat. Res. 2000, 467, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Ferrucci, L.; Nicolai, F.; Derme, V.; De Stefano, G.F. Increase of Chromosomal Aberrations Induced by Ionising Radiation in Peripheral Blood Lymphocytes of Civil Aviation Pilots and Crew Members. Mutat. Res. 1997, 377, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, J.S.; Butler, G.C.; Davis, S.; Bryant, E.; Hoel, D.G.; Mohr, L.C. Stable Chromosome Aberrations and Ionizing Radiation in Airline Pilots. Aviat. Space Environ. Med. 2003, 74, 953–956. [Google Scholar] [PubMed]
- Cavallo, D.; Marinaccio, A.; Perniconi, B.; Tomao, P.; Pecoriello, V.; Moccaldi, R.; Iavicoli, S. Chromosomal Aberrations in Long-Haul Air Crew Members. Mutat. Res. 2002, 513, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Yong, L.C.; Sigurdson, A.J.; Ward, E.M.; Waters, M.A.; Whelan, E.A.; Petersen, M.R.; Bhatti, P.; Ramsey, M.J.; Ron, E.; Tucker, J.D. Increased Frequency of Chromosome Translocations in Airline Pilots with Long-Term Flying Experience. Occup. Environ. Med. 2009, 66, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, Y.; Liu, J.; Zhang, H.; Guo, Y.; Zhang, Y.; Duan, S.; Peng, X.; Sun, T.; Jia, B.; et al. Influence of Cosmic Radiation on Lymphocyte Micronucleus, Serum Lipid Peroxide and Antioxidation Capacity in Aircrew Members. Chin. Sci. Bull. 2002, 47, 647–653. [Google Scholar] [CrossRef]
- Zwingmann, I.; Welle, I.; van Herwijen, M.; Engelen, J.; Schilderman, P.; Smid, T.; Kleinjans, J. Oxidative DNA Damage and Cytogenetic Effects in Flight Engineers Exposed to Cosmic Radiation. Environ. Mol. Mutagen. 1998, 32, 121–129. [Google Scholar] [CrossRef]
- Nickoloff, J.A.; Sharma, N.; Taylor, L. Clustered DNA Double-Strand Breaks: Biological Effects and Relevance to Cancer Radiotherapy. Genes 2020, 11, 99. [Google Scholar] [CrossRef]
- Novo, N.; Romero-Tamayo, S.; Marcuello, C.; Boneta, S.; Blasco-Machin, I.; Velázquez-Campoy, A.; Villanueva, R.; Moreno-Loshuertos, R.; Lostao, A.; Medina, M.; et al. Beyond a Platform Protein for the Degradosome Assembly: The Apoptosis-Inducing Factor as an Efficient Nuclease Involved in Chromatinolysis. PNAS Nexus 2023, 2, pgac312. [Google Scholar] [CrossRef]
- Siklos, M.; Kubicek, S. Therapeutic Targeting of Chromatin: Status and Opportunities. FEBS J. 2022, 289, 1276–1301. [Google Scholar] [CrossRef] [PubMed]
- Nuszkiewicz, J.; Woźniak, A.; Szewczyk-Golec, K. Ionizing Radiation as a Source of Oxidative Stress—The Protective Role of Melatonin and Vitamin D. Int. J. Mol. Sci. 2020, 21, 5804. [Google Scholar] [CrossRef] [PubMed]
- Hada, M.; Georgakilas, A.G. Formation of Clustered DNA Damage after High-LET Irradiation: A Review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jella, K.K.; Jaafar, L.; Li, S.; Park, S.; Story, M.D.; Wang, H.; Wang, Y.; Dynan, W.S. Exposure to Galactic Cosmic Radiation Compromises DNA Repair and Increases the Potential for Oncogenic Chromosomal Rearrangement in Bronchial Epithelial Cells. Sci. Rep. 2018, 8, 11038. [Google Scholar] [CrossRef] [PubMed]
- Asaithamby, A.; Chen, D.J. Mechanism of Cluster DNA Damage Repair in Response to High-Atomic Number and Energy Particles Radiation. Mutat. Res. 2011, 711, 87. [Google Scholar] [CrossRef] [PubMed]
- Campa, A.; Alloni, D.; Antonelli, F.; Ballarini, F.; Belli, M.; Dini, V.; Esposito, G.; Facoetti, A.; Friedland, W.; Furusawa, Y.; et al. DNA Fragmentation Induced in Human Fibroblasts by 56Fe Ions: Experimental Data and Monte Carlo Simulations. Radiat. Res. 2009, 171, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Nagamatsu, A.; Nenoi, M.; Fujimori, A.; Kakinuma, S.; Katsube, T.; Wang, B.; Tsuruoka, C.; Shirai, T.; Nakamura, A.J.; et al. Space Radiation Biology for “Living in Space”. Biomed. Res. Int. 2020, 2020, 4703286. [Google Scholar] [CrossRef] [PubMed]
- Juerb, D.; Zwar, M.; Giesen, U.; Nolte, R.; Kriesen, S.; Baiocco, G.; Puchalska, M.; van Goethem, M.J.; Manda, K.; Hildebrandt, G. Comparative Study of the Effects of Different Radiation Qualities on Normal Human Breast Cells. Radiat. Oncol. 2017, 12, 159. [Google Scholar] [CrossRef]
- Ma, A.; Dai, X. The Relationship between DNA Single-Stranded Damage Response and Double-Stranded Damage Response. Cell Cycle 2018, 17, 73–79. [Google Scholar] [CrossRef]
- Ward, J.F. Some Biochemical Consequences of the Spatial Distribution of Ionizing Radiation-Produced Free Radicals. Radiat. Res. 1981, 86, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, C.; Sun, F.; Wei, W.; Hu, B.; Wang, J. Both Complexity and Location of DNA Damage Contribute to Cellular Senescence Induced by Ionizing Radiation. PLoS ONE 2016, 11, e0155725. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, P.G.S.; Escalada, N.D.; Blakely, W.F. Induction of Premature Chromosome Condensation by a Phosphatase Inhibitor and a Protein Kinase in Unstimulated Human Peripheral Blood Lymphocytes: A Simple and Rapid Technique to Study Chromosome Aberrations Using Specific Whole-Chromosome DNA Hybridization Probes for Biological Dosimetry. Mutat. Res. 2000, 466, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Pujol-Canadell, M.; Perrier, J.R.; Cunha, L.; Shuryak, I.; Harken, A.; Garty, G.; Brenner, D.J. Cytogenetically-Based Biodosimetry after High Doses of Radiation. PLoS ONE 2020, 15, e0228350. [Google Scholar] [CrossRef] [PubMed]
- Leatherbarrow, E.L.; Harper, J.V.; Cucinotta, F.A.; O’Neill, P. Induction and Quantification of Gamma-H2AX Foci Following Low and High LET-Irradiation. Int. J. Radiat. Biol. 2006, 82, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Rose Li, Y.; Halliwill, K.D.; Adams, C.J.; Iyer, V.; Riva, L.; Mamunur, R.; Jen, K.Y.; del Rosario, R.; Fredlund, E.; Hirst, G.; et al. Mutational Signatures in Tumours Induced by High and Low Energy Radiation in Trp53 Deficient Mice. Nat. Commun. 2020, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, A.; Nasonova, E.; Czerski, K.; Kutsalo, P.; Pereira, W.; Krasavin, E. Production and Distribution of Chromosome Aberrations in Human Lymphocytes by Particle Beams with Different LET. Radiat. Environ. Biophys. 2019, 58, 99. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Engelbrecht, M.; Miles, X.; Ndimba, R.; Fisher, R.; du Plessis, P.; Bolcaen, J.; Nieto-Camero, J.; de Kock, E.; Vandevoorde, C. The Impact of Dose Rate on DNA Double-Strand Break Formation and Repair in Human Lymphocytes Exposed to Fast Neutron Irradiation. Int. J. Mol. Sci. 2019, 20, 5350. [Google Scholar] [CrossRef] [PubMed]
- Grayson, J.; Lyons, T. Brain Cancer, Flying, and Socioeconomic Status: A Nested Case-Control Study of USAF Aircrew. Aviat. Space Environ. Med. 1996, 67, 1152–1154. [Google Scholar]
- Mortazavi, S. The Safety Issues of Onboard Wi-Fi: Possible Interactions of Oxidative Stress-Causing High Altitude, Cosmic Radiation, and Wi-Fi Radiation. React. Oxyg. Species 2017, 4, 441–444. [Google Scholar] [CrossRef]
- Akdag, M.Z.; Dasdag, S.; Canturk, F.; Karabulut, D.; Caner, Y.; Adalier, N. Does Prolonged Radiofrequency Radiation Emitted from Wi-Fi Devices Induce DNA Damage in Various Tissues of Rats? J. Chem. Neuroanat. 2016, 75, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Bodewein, L.; Schmiedchen, K.; Dechent, D.; Stunder, D.; Graefrath, D.; Winter, L.; Kraus, T.; Driessen, S. Systematic Review on the Biological Effects of Electric, Magnetic and Electromagnetic Fields in the Intermediate Frequency Range (300 Hz to 1 MHz). Environ. Res. 2019, 171, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, D.; Tomao, P.; Marinaccio, A.; Perniconi, B.; Setini, A.; Palmi, S.; Iavicoli, S. Evaluation of DNA Damage in Flight Personnel by Comet Assay. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2002, 516, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Minoretti, P.; Riera, M.L.; Sáez, A.S.; Serrano, M.G.; Martín, Á.G. Increased Peripheral Blood DNA Damage and Elevated Serum Levels of Melanoma Inhibitory Activity Protein: Clues to Excess Skin Cancer Risk in Airline Pilots? Cureus 2023, 15, e51077. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G. Fundamental Space Radiobiology. Gravit. Space Biol. Bull. 2003, 16, 29–36. [Google Scholar] [PubMed]
- Seth, I.; Schwartz, J.L.; Stewart, R.D.; Emery, R.; Joiner, M.C.; Tucker, J.D. Neutron Exposures in Human Cells: Bystander Effect and Relative Biological Effectiveness. PLoS ONE 2014, 9, e0098947. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Wang, H.; Perrault, A.R.; Boecker, W.; Rosidi, B.; Windhofer, F.; Wu, W.; Guan, J.; Terzoudi, G.; Panteliasc, G. Mechanisms of DNA Double Strand Break Repair and Chromosome Aberration Formation. Cytogenet. Genome Res. 2004, 104, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Bauchinger, M. Quantification of Low-Level Radiation Exposure by Conventional Chromosome Aberration Analysis. Mutat. Res. Genet. Toxicol. 1995, 339, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Cologne, J.; Sugiyama, H.; Hamasaki, K.; Tatsukawa, Y.; French, B.; Sakata, R.; Misumi, M. Chromosome Aberrations among Atomic-Bomb Survivors Exposed in Utero: Updated Analysis Accounting for Revised Radiation Doses and Smoking. Radiat. Environ. Biophys. 2022, 61, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Ainsbury, E.A.; Livingston, G.K.; Abbott, M.G.; Moquet, J.E.; Hone, P.A.; Jenkins, M.S.; Christensen, D.M.; Lloyd, D.C.; Rothkamm, K. Interlaboratory Variation in Scoring Dicentric Chromosomes in a Case of Partial-Body x-Ray Exposure: Implications for Biodosimetry Networking and Cytogenetic “Triage Mode” Scoring. Radiat. Res. 2009, 172, 746–752. [Google Scholar] [CrossRef]
- CA, K.; Heintz, P.; Sandoval, D.; Chambers, G.; Adolphi, N.; Paffett, K. Radiation Effects on Tissues and Organs; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014. [Google Scholar]
- Ministry of the Environment Government of Japan (JCN1000012110001). Radiosensitivity of Organs and Tissues [MOE]. In Booklet to Provide Basic Information Regarding Health Effects of Radiation; Ministry of the Environment Government of Japan: Tokyo, Japan, 2013. [Google Scholar]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of Apoptosis in Health and Disease: The Balancing Act of BCL-2 Family Proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Powell, D.R.; Li, Z.; Bell, J.S.K.; Barwick, B.G.; Feng, H.; McCrary, M.R.; Dwivedi, B.; Kowalski, J.; Dynan, W.S.; et al. Galactic Cosmic Radiation Induces Persistent Epigenome Alterations Relevant to Human Lung Cancer. Sci. Rep. 2018, 8, 6709. [Google Scholar] [CrossRef] [PubMed]
- Gage, F.H. Mammalian Neural Stem Cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the Adult Human Hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A. The Mystery and Magic of Glia: A Perspective on Their Roles in Health and Disease. Neuron 2008, 60, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.A.; Hudspeth, A.J.M.S. (Eds.) Principles of Neural Science, 5th ed.; McGraw-Hill Education: New York, NY, USA, 2014. [Google Scholar]
- Tiwari, V.; Wilson, D.M. DNA Damage and Associated DNA Repair Defects in Disease and Premature Aging. Am. J. Hum. Genet. 2019, 105, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Limoli, C.L.; Ponnaiya, B.; Corcoran, J.J.; Giedzinski, E.; Kaplan, M.I.; Hartmann, A.; Morgan, W.F. Genomic Instability Induced by High and Low LET Ionizing Radiation. Adv. Space Res. 2000, 25, 2107–2117. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Dai, W. Genomic Instability and Cancer. J. Carcinog. Mutagen. 2015, 5, 1000165. [Google Scholar]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef]
- IARC (International Agency for Research on Cancer); IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Tobacco Smoke and Involuntary Smoking; International Agency for Research on Cancer: Lyon, France, 2004. [Google Scholar]
- IARC (International Agency for Research on Cancer); IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Painting, Firefighting, and Shiftwork; International Agency for Research on Cancer: Lyon, France, 2010. [Google Scholar]
- da Silva, J. DNA Damage Induced by Occupational and Environmental Exposure to Miscellaneous Chemicals. Mutat. Res. Rev. Mutat. Res. 2016, 770, 170–182. [Google Scholar] [CrossRef]
- Cheng, L.; Brzozowska, B.; Sollazzo, A.; Lundholm, L.; Lisowska, H.; Haghdoost, S.; Wojcik, A. Simultaneous Induction of Dispersed and Clustered DNA Lesions Compromises DNA Damage Response in Human Peripheral Blood Lymphocytes. PLoS ONE 2018, 13, e0204068. [Google Scholar] [CrossRef] [PubMed]
- Soren, D.C.; Toprani, S.M.; Jain, V.; Saini, D.; Das, B. Quantitation of Genome Damage and Transcriptional Profile of DNA Damage Response Genes in Human Peripheral Blood Mononuclear Cells Exposed in Vitro to Low Doses of Neutron Radiation. Int. J. Radiat. Res. 2019, 17, 1–14. [Google Scholar] [CrossRef]
- Roobol, S.J.; van den Bent, I.; van Cappellen, W.A.; Abraham, T.E.; Paul, M.W.; Kanaar, R.; Houtsmuller, A.B.; van Gent, D.C.; Essers, J. Comparison of High- and Low-LET Radiation-Induced DNA Double-Strand Break Processing in Living Cells. Int. J. Mol. Sci. 2020, 21, 6602. [Google Scholar] [CrossRef] [PubMed]
- Slatter, M.A.; Gennery, A.R. Update on DNA-Double Strand Break Repair Defects in Combined Primary Immunodeficiency. Curr. Allergy Asthma Rep. 2020, 20, 57. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Lewis, S.; Wlodarski, M.W. DNA Repair Syndromes and Cancer: Insights Into Genetics and Phenotype Patterns. Front. Pediatr. 2020, 8, 570084. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Chen, B.P.C.; Chen, D.J. DNA-PK: A Dynamic Enzyme in a Versatile DSB Repair Pathway. DNA Repair 2014, 17, 21–29. [Google Scholar] [CrossRef]
- Pang, D.; Winters, T.A.; Jung, M.; Purkayastha, S.; Cavalli, L.R.; Chasovkikh, S.; Haddad, B.R.; Dritschilo, A. Radiation-Generated Short DNA Fragments May Perturb Non-Homologous End-Joining and Induce Genomic Instability. J. Radiat. Res. 2011, 52, 309–319. [Google Scholar] [CrossRef]
- Yuan, Y.; Britton, S.; Delteil, C.; Coates, J.; Jackson, S.P.; Barboule, N.; Frit, P.; Calsou, P. Single-Stranded DNA Oligomers Stimulate Error-Prone Alternative Repair of DNA Double-Strand Breaks through Hijacking Ku Protein. Nucleic Acids Res. 2015, 43, 10264–10276. [Google Scholar] [CrossRef]
- Schmid, T.E.; Dollinger, G.; Beisker, W.; Hable, V.; Greubel, C.; Auer, S.; Mittag, A.; Tarnok, A.; Friedl, A.A.; Molls, M.; et al. Differences in the Kinetics of Gamma-H2AX Fluorescence Decay after Exposure to Low and High LET Radiation. Int. J. Radiat. Biol. 2010, 86, 682–691. [Google Scholar] [CrossRef]
- Allen, C.P.; Hirakawa, H.; Nakajima, N.I.; Moore, S.; Nie, J.; Sharma, N.; Sugiura, M.; Hoki, Y.; Araki, R.; Abe, M.; et al. Low- and High-LET Ionizing Radiation Induces Delayed Homologous Recombination That Persists for Two Weeks before Resolving. Radiat. Res. 2017, 188, 82–93. [Google Scholar] [CrossRef]
- Huang, L.; Grim, S.; Smith, L.E.; Kim, P.M.; Nickoloff, J.A.; Goloubeva, O.G.; Morgan, W.F. Ionizing Radiation Induces Delayed Hyperrecombination in Mammalian Cells. Mol. Cell. Biol. 2004, 24, 5060–5068. [Google Scholar] [CrossRef] [PubMed]
- Grosse, N.; Fontana, A.O.; Hug, E.B.; Lomax, A.; Coray, A.; Augsburger, M.; Paganetti, H.; Sartori, A.A.; Pruschy, M. Deficiency in Homologous Recombination Renders Mammalian Cells More Sensitive to Proton versus Photon Irradiation. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Seol, J.H.; Shim, E.Y.; Lee, S.E. Microhomology-Mediated End Joining: Good, Bad and Ugly. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.D.; Doublié, S. DNA Polymerase θ (POLQ), Double-Strand Break Repair, and Cancer. DNA Repair 2016, 44, 22–32. [Google Scholar] [CrossRef]
- Feng, W.; Smith, C.M.; Simpson, D.A.; Gupta, G.P. Targeting Non-Homologous and Alternative End Joining Repair to Enhance Cancer Radiosensitivity. Semin. Radiat. Oncol. 2022, 32, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Krokan, H.E.; Bjørås, M. Base Excision Repair. Cold Spring Harb. Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Galick, H.; Wallace, S.S. Attempted Base Excision Repair of Ionizing Radiation Damage in Human Lymphoblastoid Cells Produces Lethal and Mutagenic Double Strand Breaks. DNA Repair 2004, 3, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.; Demple, B. Distinct Roles of Ape1 Protein in the Repair of DNA Damage Induced by Ionizing Radiation or Bleomycin. J. Biol. Chem. 2011, 286, 4968–4977. [Google Scholar] [CrossRef]
- Eccles, L.J.; O’Neill, P.; Lomax, M.E. Delayed Repair of Radiation Induced Clustered DNA Damage: Friend or Foe? Mutat. Res. 2011, 711, 134–141. [Google Scholar] [CrossRef]
- Rajaraman, P.; Bhatti, P.; Doody, M.M.; Simon, S.L.; Weinstock, R.M.; Linet, M.S.; Rosenstein, M.; Stovall, M.; Alexander, B.H.; Preston, D.L.; et al. Nucleotide Excision Repair Polymorphisms May Modify Ionizing Radiation-Related Breast Cancer Risk in US Radiologic Technologists. Int. J. Cancer 2008, 123, 2713–2716. [Google Scholar] [CrossRef]
- Terzidis, M.A.; Ferreri, C.; Chatgilialoglu, C. Radiation-Induced Formation of Purine Lesions in Single and Double Stranded DNA: Revised Quantification. Front. Chem. 2015, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Raja, S.; Van Houten, B. The Involvement of Nucleotide Excision Repair Proteins in the Removal of Oxidative DNA Damage. Nucleic Acids Res. 2020, 48, 11227–11243. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Mattson, M.P. How Does Hormesis Impact Biology, Toxicology, and Medicine? NPJ Aging Mech. Dis. 2017, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Latimer, J.J.; Alhamed, A.; Sveiven, S.; Almutairy, A.; Klimas, N.G.; Abreu, M.; Sullivan, K.; Grant, S.G. Preliminary Evidence for a Hormetic Effect on DNA Nucleotide Excision Repair in Veterans with Gulf War Illness. Mil. Med. 2020, 185, E47–E52. [Google Scholar] [CrossRef]
- Kottemann, M.C.; Smogorzewska, A. Fanconi Anaemia and the Repair of Watson and Crick DNA Crosslinks. Nature 2013, 493, 356–363. [Google Scholar] [CrossRef]
- Sanlorenzo, M.; Wehner, M.R.; Linos, E.; Kornak, J.; Kainz, W.; Posch, C.; Vujic, I.; Johnston, K.; Gho, D.; Monico, G.; et al. The Risk of Melanoma in Airline Pilots and Cabin Crew: A Meta-Analysis. JAMA Dermatol. 2015, 151, 51–58. [Google Scholar] [CrossRef]
- Institute, P. Women Pilot Statistics: Female Representation in Aviation-Pilot Institute. Available online: https://pilotinstitute.com/women-aviation-statistics/#:~:text=This is a massive 151,compared to 6.03%25 in 2021 (accessed on 3 January 2023).
- National Institute for Occupational Safety and Health (NIOSH). Hierarchy of Controls. NIOSH Workplace Safety and Health Topics; National Institute for Occupational Safety and Health: Washington, DC, USA, 2015. [Google Scholar]
- Radon, K.; Goldberg, M.; Becklake, M. Healthy Worker Effect in Cohort Studies on Chronic Bronchitis. Scand. J. Work Environ. Health 2002, 28, 328–332. [Google Scholar] [CrossRef]
- Ge, J.; Prasongtanakij, S.; Wood, D.K.; Weingeist, D.M.; Fessler, J.; Navasummrit, P.; Ruchirawat, M.; Engelward, B.P. Cometchip: A High-Throughput 96-Well Platform for Measuring DNA Damage in Microarrayed Human Cells. J. Vis. Exp. 2014, 18, e50607. [Google Scholar] [CrossRef]
- Garty, G.; Chen, Y.; Salerno, A.; Turner, H.; Zhang, J.; Lyulko, O.; Bertucci, A.; Xu, Y.; Wang, H.; Simaan, N.; et al. The RABIT: A Rapid Automated Biodosimetry Tool for Radiological Triage. Health Phys. 2010, 98, 209–217. [Google Scholar] [CrossRef]
- Nagel, Z.D.; Margulies, C.M.; Chaim, I.A.; McRee, S.K.; Mazzucato, P.; Ahmad, A.; Abo, R.P.; Butty, V.L.; Forget, A.L.; Samson, L.D. Multiplexed DNA Repair Assays for Multiple Lesions and Multiple Doses via Transcription Inhibition and Transcriptional Mutagenesis. Proc. Natl. Acad. Sci. USA 2014, 111, E1823–E1832. [Google Scholar] [CrossRef]
- Nagel, Z.D.; Beharry, A.A.; Mazzucato, P.; Kitange, G.J.; Sarkaria, J.N.; Kool, E.T.; Samson, L.D. Fluorescent Reporter Assays Provide Direct, Accurate, Quantitative Measurements of MGMT Status in Human Cells. PLoS ONE 2019, 14, e0208341. [Google Scholar] [CrossRef]
- Nagel, Z.D.; Engelward, B.P.; Brenner, D.J.; Begley, T.J.; Sobol, R.W.; Bielas, J.H.; Stambrook, P.J.; Wei, Q.; Hu, J.J.; Terry, M.B.; et al. Towards Precision Prevention: Technologies for Identifying Healthy Individuals with High Risk of Disease. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2017, 800–802, 14–28. [Google Scholar] [CrossRef]
- Cheong, A.; Nagel, Z.D. Human Variation in DNA Repair, Immune Function, and Cancer Risk. Front. Immunol. 2022, 13, 899574. [Google Scholar] [CrossRef]
- Hornhardt, S.; Rößler, U.; Sauter, W.; Rosenberger, A.; Illig, T.; Bickeböller, H.; Wichmann, H.E.; Gomolka, M. Genetic Factors in Individual Radiation Sensitivity. DNA Repair 2014, 16, 54–65. [Google Scholar] [CrossRef]
- Wu, H.C.; Kehm, R.; Santella, R.M.; Brenner, D.J.; Terry, M.B. DNA Repair Phenotype and Cancer Risk: A Systematic Review and Meta-Analysis of 55 Case–Control Studies. Sci. Rep. 2022, 12, 3405. [Google Scholar] [CrossRef]
- Luxton, J.J.; McKenna, M.J.; Lewis, A.; Taylor, L.E.; George, K.A.; Dixit, S.M.; Moniz, M.; Benegas, W.; Mackay, M.J.; Mozsary, C.; et al. Telomere Length Dynamics and DNA Damage Responses Associated with Long-Duration Spaceflight. Cell Rep. 2020, 33, 108457. [Google Scholar] [CrossRef]
- Marnell, C.S.; Bick, A.; Natarajan, P. Clonal Hematopoiesis of Indeterminate Potential (CHIP): Linking Somatic Mutations, Hematopoiesis, Chronic Inflammation and Cardiovascular Disease. J. Mol. Cell. Cardiol. 2021, 161, 98–105. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
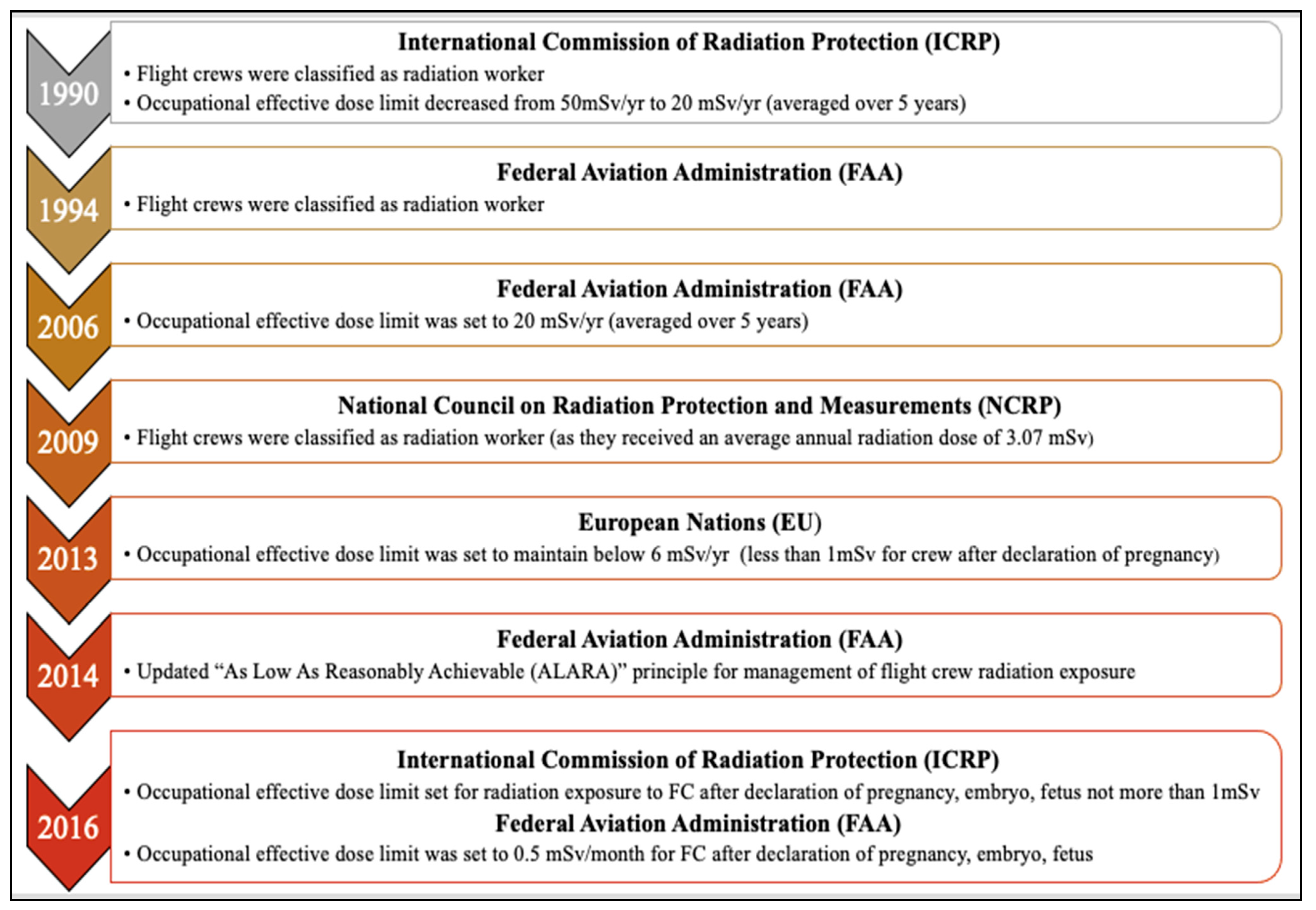
| Exposure limits | International Commission of Radiation Protection (ICRP) [23] | European Nations (EU) [43,44] | US National Council on Radiation Protection and Measurements (NCRP) [4] | US Nuclear Regulatory Commission, (NRC) [22] | US Federal Aviation Administration (FAA) [45] |
|---|---|---|---|---|---|
| General public | 1 mSv/y | 1 mSv/y | 1 mSv/y | 1 mSv/y | 1 mSv/y |
| Pregnant women and unborn fetuses | 1 mSv after declaration of pregnancy | 1 mSv after declaration of pregnancy | 5 mSv and no more than 0.5 mSv in any month | 5 mSv and no more than 0.5 mSv in any month | 1 mSv and no more than 0.5 mSv in any month |
| Occupational exposure | 20 mSv/y * | 20 mSv/y * employer required monitoring and administrative controls to maintain <6 mSv/y | 50 mSv/y | 50 mSv/y | 20 mSv/y * recommendation for FC to self-monitor without requirements for the employer |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toprani, S.M.; Scheibler, C.; Mordukhovich, I.; McNeely, E.; Nagel, Z.D. Cosmic Ionizing Radiation: A DNA Damaging Agent That May Underly Excess Cancer in Flight Crews. Int. J. Mol. Sci. 2024, 25, 7670. https://doi.org/10.3390/ijms25147670
Toprani SM, Scheibler C, Mordukhovich I, McNeely E, Nagel ZD. Cosmic Ionizing Radiation: A DNA Damaging Agent That May Underly Excess Cancer in Flight Crews. International Journal of Molecular Sciences. 2024; 25(14):7670. https://doi.org/10.3390/ijms25147670
Chicago/Turabian StyleToprani, Sneh M., Christopher Scheibler, Irina Mordukhovich, Eileen McNeely, and Zachary D. Nagel. 2024. "Cosmic Ionizing Radiation: A DNA Damaging Agent That May Underly Excess Cancer in Flight Crews" International Journal of Molecular Sciences 25, no. 14: 7670. https://doi.org/10.3390/ijms25147670
APA StyleToprani, S. M., Scheibler, C., Mordukhovich, I., McNeely, E., & Nagel, Z. D. (2024). Cosmic Ionizing Radiation: A DNA Damaging Agent That May Underly Excess Cancer in Flight Crews. International Journal of Molecular Sciences, 25(14), 7670. https://doi.org/10.3390/ijms25147670