
Sex-Specific Changes to Brain Fatty Acids, Plasmalogen, and Plasma Endocannabinoids in Offspring Exposed to Maternal and Postnatal High-Linoleic-Acid Diets
 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
2.1. Maternal and Postnatal Consumption of HLA Diet Affects the Brain’s Saturated Fatty Acid, Monounsaturated Fatty Acid, and Polyunsaturated Fatty Acid Composition in Adult Offspring in a Sex-Dependent Manner
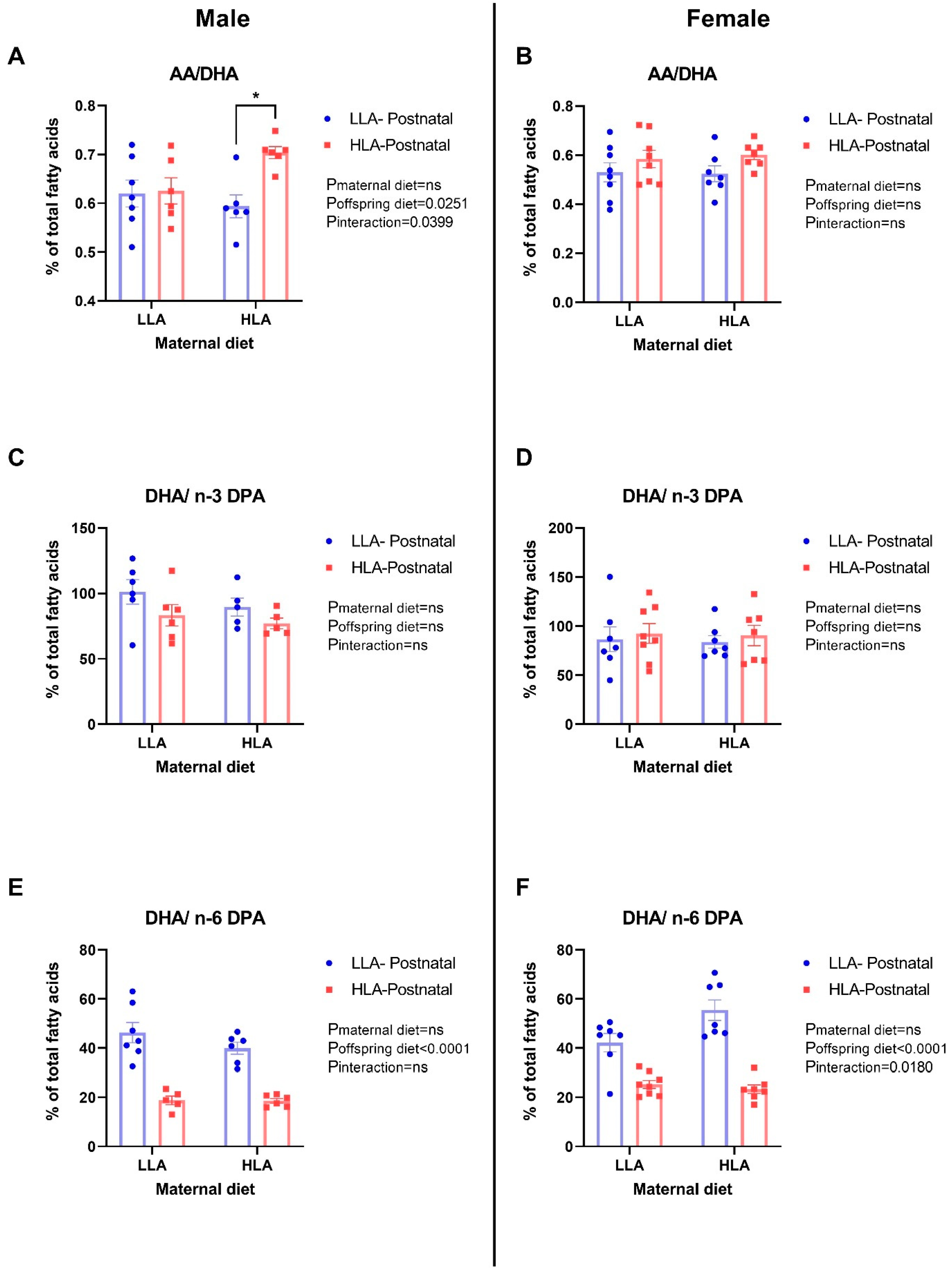
2.2. Maternal and Postnatal Consumption of an HLA Diet Affects Brain AA/DHA, DHA/n-3 DPA, and DHA/n-6 DPA in a Sex-Dependent Manner
2.3. Maternal HLA Diet Increases Brain Plasmalogen (C16:0 DMA/C16:0 and C18:0 DMA/C18:0) in Male Offspring
2.4. Maternal and Postnatal HLA Diets Alter Plasma Endocannabinoids in Adult Offspring, in a Sex-Specific Manner
2.5. Effects of Maternal and Postnatal HLA Diets on Plasma N-acyl Ethanolamide in Adult Offspring
2.6. Effects of Maternal and Postnatal HLA Diets on Plasma Steroid Hormones in Adult Offspring
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Model and Diet
4.2. Rodent Brain Fatty Acid Analysis
4.3. Plasma Endocannabinoid Analysis
4.4. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taha, A.Y. Linoleic acid-good or bad for the brain? NPJ Sci. Food 2020, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Linoleic acid and the pathogenesis of obesity. Prostaglandins Other Lipid Mediat. 2016, 125, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Marchix, J.; Choque, B.; Kouba, M.; Fautrel, A.; Catheline, D.; Legrand, P. Excessive dietary linoleic acid induces proinflammatory markers in rats. J. Nutr. Biochem. 2015, 26, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Cuffe, J.S.M.; Holland, O.J.; Bulmer, A.C.; Hill, M.; Perkins, A.V.; Muhlhausler, B.S.; McAinch, A.J.; Hryciw, D.H. Elevated maternal linoleic acid reduces circulating leptin concentrations, cholesterol levels and male fetal survival in a rat model. J. Physiol. 2019, 597, 3349–3361. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Sleep, S.; Helman, T.; Holland, O.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Headrick, J.P.; Hryciw, D.H. Maternal diet high in linoleic acid alters offspring fatty acids and cardiovascular function in a rat model. Br. J. Nutr. 2022, 127, 540–553. [Google Scholar] [CrossRef]
- Shrestha, N.; Sleep, S.L.; Holland, O.J.; Vidimce, J.; Bulmer, A.C.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Maternal Diet High in Linoleic Acid Alters Offspring Lipids and Hepatic Regulators of Lipid Metabolism in an Adolescent Rat Model. Int. J. Mol. Sci. 2024, 25, 1129. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Vidimce, J.; Holland, O.J.; Cuffe, J.S.M.; Beck, B.R.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Maternal and Postnatal High Linoleic Acid Diet Impacts Lipid Metabolism in Adult Rat Offspring in a Sex-Specific Manner. Int. J. Mol. Sci. 2021, 22, 2946. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.; Álvarez, D.; Muñoz, Y.; Crisosto, N.; Valenzuela, R.; Maliqueo, M. Linoleic and arachidonic fatty acids and their potential relationship with inflammation, pregnancy, and fetal development. Curr. Med. Chem. 2023, 31, 5046–5060. [Google Scholar] [CrossRef] [PubMed]
- Zengeler, K.E.; Lukens, J.R. Inflammation stresses out brain development. Nat. Neurosci. 2021, 24, 155–157. [Google Scholar] [CrossRef]
- Yau, S.Y.; Yip, Y.S.L.; Formolo, D.A.; He, S.; Lee, T.H.Y.; Wen, C.; Hryciw, D.H. Chronic consumption of a high linoleic acid diet during pregnancy, lactation and post-weaning period increases depression-like behavior in male, but not female offspring. Behav. Brain Res. 2022, 416, 113538. [Google Scholar] [CrossRef]
- Giles, C.; Takechi, R.; Mellett, N.A.; Meikle, P.J.; Dhaliwal, S.; Mamo, J.C. The Effects of Long-Term Saturated Fat Enriched Diets on the Brain Lipidome. PLoS ONE 2016, 11, e0166964. [Google Scholar] [CrossRef]
- Brites, P.; Waterham, H.R.; Wanders, R.J. Functions and biosynthesis of plasmalogens in health and disease. Biochim. Biophys. Acta. 2004, 1636, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Cunnane, S.C.; Schneider, J.A.; Tangney, C.; Tremblay-Mercier, J.; Fortier, M.; Bennett, D.A.; Morris, M.C. Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer’s disease. J. Alzheimers Dis. 2012, 29, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, C.; Saitoh, M.; Itoh, M.; Yamashita, S.; Miyagishi, M.; Takashima, S.; Moser, A.B.; Iwamori, M.; Mizuguchi, M. Altered phospholipid molecular species and glycolipid composition in brain, liver and fibroblasts of Zellweger syndrome. Neurosci. Lett. 2013, 552, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Vanier, M.T. Lipid changes in Niemann-Pick disease type C brain: Personal experience and review of the literature. Neurochem. Res. 1999, 24, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, C.E.; Domenichiello, A.F.; Yuan, Z.X.; Sapio, M.R.; Keyes, G.S.; Mishra, S.K.; Gross, J.R.; Majchrzak-Hong, S.; Zamora, D.; Horowitz, M.S.; et al. A systems approach for discovering linoleic acid derivatives that potentially mediate pain and itch. Sci. Signal. 2017, 10, eaal5241. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, A.M.; Scotland, P.E.; Akopian, A.N.; Hargreaves, K.M. Activation of TRPV1 in the spinal cord by oxidized linoleic acid metabolites contributes to inflammatory hyperalgesia. Proc. Natl. Acad. Sci. USA 2009, 106, 18820–18824. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Cinquina, V.; Keimpema, E.; Pollak, D.D.; Harkany, T. Adverse effects of gestational ω-3 and ω-6 polyunsaturated fatty acid imbalance on the programming of fetal brain development. J. Neuroendocrinol. 2023, 35, e13320. [Google Scholar] [CrossRef]
- Honsho, M.; Fujiki, Y. Regulation of plasmalogen biosynthesis in mammalian cells and tissues. Brain Res. Bull. 2023, 194, 118–123. [Google Scholar] [CrossRef]
- Honsho, M.; Fujiki, Y. Plasmalogen homeostasis—Regulation of plasmalogen biosynthesis and its physiological consequence in mammals. FEBS. Lett. 2017, 591, 2720–2729. [Google Scholar] [CrossRef] [PubMed]
- Brites, P.; Mooyer, P.A.; El Mrabet, L.; Waterham, H.R.; Wanders, R.J. Plasmalogens participate in very-long-chain fatty acid-induced pathology. Brain 2009, 132, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, V.; Goodenowe, D.B. Plasmalogen deficiency and neuropathology in Alzheimer’s disease: Causation or coincidence? Alzheimers Dement. 2019, 5, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Moraitou, M.; Dimitriou, E.; Zafeiriou, D.; Reppa, C.; Marinakis, T.; Sarafidou, J.; Michelakakis, H. Plasmalogen levels in Gaucher disease. Blood Cells Mol. Dis. 2008, 41, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From Phytocannabinoids to Cannabinoid Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Roles Through Complex Pharmacology. Physiol. Rev. 2016, 96, 1593–1659. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Sun, J.; Zhang, W.; Guo, Z.; Ma, Q. Arachidonic acid metabolism in health and disease. MedComm 2023, 4, e363. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-mediated control of synaptic transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Amir Hamzah, K.; Toms, L.M.; Kucharski, N.; Orr, J.; Turner, N.P.; Hobson, P.; Nichols, D.S.; Ney, L.J. Sex-dimorphism in human serum endocannabinoid and n-acyl ethanolamine concentrations across the lifespan. Sci. Rep. 2023, 13, 23059. [Google Scholar] [CrossRef]
- Ney, L.J.; Akhurst, J.; Bruno, R.; Laing, P.A.F.; Matthews, A.; Felmingham, K.L. Dopamine, endocannabinoids and their interaction in fear extinction and negative affect in PTSD. Prog Neuro-Psychopharmacol. Biol. Psychiatry 2021, 105, 110118. [Google Scholar] [CrossRef] [PubMed]
- Bjerregaard, A.A.; Halldorsson, T.I.; Tetens, I.; Olsen, S.F. Mother’s dietary quality during pregnancy and offspring’s dietary quality in adolescence: Follow-up from a national birth cohort study of 19,582 mother-offspring pairs. PLoS Med. 2019, 16, e1002911. [Google Scholar]
- Raipuria, M.; Bahari, H.; Morris, M.J. Effects of maternal diet and exercise during pregnancy on glucose metabolism in skeletal muscle and fat of weanling rats. PLoS ONE 2015, 10, e0120980. [Google Scholar] [CrossRef] [PubMed]
- Campodonico-Burnett, W.; Hetrick, B.; Wesolowski, S.R.; Schenk, S.; Takahashi, D.L.; Dean, T.A.; Sullivan, E.L.; Kievit, P.; Gannon, M.; Aagaard, K.; et al. Maternal Obesity and Western-Style Diet Impair Fetal and Juvenile Offspring Skeletal Muscle Insulin-Stimulated Glucose Transport in Nonhuman Primates. Diabetes 2020, 69, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Winett, L.; Wallack, L.; Richardson, D.; Boone-Heinonen, J.; Messer, L. A Framework to Address Challenges in Communicating the Developmental Origins of Health and Disease. Curr. Environ. Health Rep. 2016, 3, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.F.; Mutch, D.M.; Leri, F. The Relationship between Fatty Acids and Different Depression-Related Brain Regions, and Their Potential Role as Biomarkers of Response to Antidepressants. Nutrients 2017, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Ghazali, R.; Mehta, K.J.; Bligh, S.A.; Tewfik, I.; Clemens, D.; Patel, V.B. High omega arachidonic acid/docosahexaenoic acid ratio induces mitochondrial dysfunction and altered lipid metabolism in human hepatoma cells. World J. Hepatol. 2020, 12, 84–98. [Google Scholar] [CrossRef]
- Tanaka, T.; Shen, J.; Abecasis, G.R.; Kisialiou, A.; Ordovas, J.M.; Guralnik, J.M.; Singleton, A.; Bandinelli, S.; Cherubini, A.; Arnett, D.; et al. Genome-wide association study of plasma polyunsaturated fatty acids in the InCHIANTI Study. PLoS Genet. 2009, 5, e1000338. [Google Scholar] [CrossRef]
- Ma, Q.L.; Zhu, C.; Morselli, M.; Su, T.; Pelligrini, M.; Lu, Z.; Jones, M.; Denver, P.; Castro, D.; Gu, X.; et al. The Novel Omega-6 Fatty Acid Docosapentaenoic Acid Positively Modulates Brain Innate Immune Response for Resolving Neuroinflammation at Early and Late Stages of Humanized APOE-Based Alzheimer’s Disease Models. Front. Immunol. 2020, 11, 558036. [Google Scholar] [CrossRef]
- Otto, S.J.; de Groot, R.H.; Hornstra, G. Increased risk of postpartum depressive symptoms is associated with slower normalization after pregnancy of the functional docosahexaenoic acid status. Prostaglandins Leukot. Essent. Fat. Acids 2003, 69, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Hoshiba, J.; Salem, N., Jr. An extraordinary degree of structural specificity is required in neural phospholipids for optimal brain function: N-6 docosapentaenoic acid substitution for docosahexaenoic acid leads to a loss in spatial task performance. J. Neurochem. 2005, 95, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Lee-Okada, H.C.; Ikeda, M.; Nakamura, T.; Saito, T.; Takata, A.; Yokomizo, T.; Iwata, N.; Kato, T.; Kasahara, T. GWAS-identified bipolar disorder risk allele in the FADS1/2 gene region links mood episodes and unsaturated fatty acid metabolism in mutant mice. Mol. Psychiatry 2023, 28, 2848–2856. [Google Scholar] [CrossRef] [PubMed]
- Helland, I.B.; Smith, L.; Saarem, K.; Saugstad, O.D.; Drevon, C.A. Maternal supplementation with very-long-chain n-3 fatty acids during pregnancy and lactation augments children’s IQ at 4 years of age. Pediatrics 2003, 111, e39–e44. [Google Scholar] [CrossRef] [PubMed]
- van Dorp, D.A. Essential fatty acid metabolism. Proc. Nutr. Soc. 1975, 34, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Sherzai, D.; Moness, R.; Sherzai, S.; Sherzai, A. A Systematic Review of Omega-3 Fatty Acid Consumption and Cognitive Outcomes in Neurodevelopment. Am. J. Lifestyle Med. 2023, 17, 649–685. [Google Scholar] [CrossRef] [PubMed]
- Román, G.C.; Jackson, R.E.; Gadhia, R.; Román, A.N.; Reis, J. Mediterranean diet: The role of long-chain ω-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Helland, I.B.; Smith, L.; Blomén, B.; Saarem, K.; Saugstad, O.D.; Drevon, C.A. Effect of supplementing pregnant and lactating mothers with n-3 very-long-chain fatty acids on children’s IQ and body mass index at 7 years of age. Pediatrics 2008, 122, e472–e479. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Liang, Y.; Tian, X.; Peng, C.; Ma, K.Y.; Liu, J.; Huang, Y.; Chen, Z.Y. DPA n-3, DPA n-6 and DHA improve lipoprotein profiles and aortic function in hamsters fed a high cholesterol diet. Atherosclerosis 2012, 221, 397–404. [Google Scholar] [CrossRef]
- Bourre, J.M. Diet, Brain Lipids, and Brain Functions: Polyunsaturated Fatty Acids, Mainly Omega-3 Fatty Acids. In Handbook of Neurochemistry and Molecular Neurobiology: Neural Lipids; Lajtha, A., Tettamanti, G., Goracci, G., Eds.; Springer: Boston, MA, USA, 2009; pp. 409–441. [Google Scholar]
- Hibbeln, J.R. Seafood consumption, the DHA content of mothers’ milk and prevalence rates of postpartum depression: A cross-national, ecological analysis. J. Affect Disord. 2002, 69, 15–29. [Google Scholar] [CrossRef]
- Levant, B. N-3 (omega-3) polyunsaturated Fatty acids in the pathophysiology and treatment of depression: Pre-clinical evidence. CNS. Neurol. Disord. Drug Targets 2013, 12, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Levant, B.; Ozias, M.K.; Carlson, S.E. Diet (n-3) polyunsaturated fatty acid content and parity interact to alter maternal rat brain phospholipid fatty acid composition. J. Nutr. 2006, 136, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Albouery, M.; Buteau, B.; Grégoire, S.; Martine, L.; Gambert, S.; Bron, A.M.; Acar, N.; Chassaing, B.; Bringer, M.A. Impact of a high-fat diet on the fatty acid composition of the retina. Exp. Eye Res. 2020, 196, 108059. [Google Scholar] [CrossRef]
- Kitson, A.P.; Stroud, C.K.; Stark, K.D. Elevated production of docosahexaenoic acid in females: Potential molecular mechanisms. Lipids 2010, 45, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Fride, E.; Gobshtis, N.; Dahan, H.; Weller, A.; Giuffrida, A.; Ben-Shabat, S. The endocannabinoid system during development: Emphasis on perinatal events and delayed effects. Vitam. Horm. 2009, 81, 139–158. [Google Scholar] [PubMed]
- Ney, L.J.; Cooper, J.; Lam, G.N.; Moffitt, K.; Nichols, D.S.; Mayo, L.M.; Lipp, O.V. Hair endocannabinoids predict physiological fear conditioning and salivary endocannabinoids predict subjective stress reactivity in humans. Psychoneuroendocrinology 2023, 154, 106296. [Google Scholar] [CrossRef]
- Thompson, Z.; Argueta, D.; Garland, T., Jr.; DiPatrizio, N. Circulating levels of endocannabinoids respond acutely to voluntary exercise, are altered in mice selectively bred for high voluntary wheel running, and differ between the sexes. Physiol. Behav. 2017, 170, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Salemme, B.W.; Raymundi, A.M.; Sohn, J.M.B.; Stern, C.A. The Estrous Cycle Influences the Effects of Fatty Acid Amide Hydrolase and Monoacylglycerol Lipase Inhibition in the Anxiety-Like Behavior in Rats. Cannabis Cannabinoid Res. 2023. [Google Scholar] [CrossRef]
- Ney, L.J.; Matthews, A.; Bruno, R.; Felmingham, K.L. Modulation of the endocannabinoid system by sex hormones: Implications for posttraumatic stress disorder. Neurosci. Biobehav. Rev. 2018, 94, 302–320. [Google Scholar] [CrossRef]
- Henricks, A.M.; Berger, A.L.; Lugo, J.M.; Baxter-Potter, L.N.; Bieniasz, K.V.; Petrie, G.; Sticht, M.A.; Hill, M.N.; McLaughlin, R.J. Sex- and hormone-dependent alterations in alcohol withdrawal-induced anxiety and corticolimbic endocannabinoid signaling. Neuropharmacology 2017, 124, 121–133. [Google Scholar] [CrossRef]
- Krebs-Kraft, D.L.; Hill, M.N.; Hillard, C.J.; McCarthy, M.M. Sex difference in cell proliferation in developing rat amygdala mediated by endocannabinoids has implications for social behavior. Proc. Natl. Acad. Sci. USA 2010, 107, 20535–20540. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; McCarron, A.; Rout-Pitt, N.; Donnelley, M.; Parsons, D.W.; Hryciw, D.H. Essential Fatty Acid Deficiency in Cystic Fibrosis Disease Progression: Role of Genotype and Sex. Nutrients 2022, 14, 4666. [Google Scholar] [CrossRef] [PubMed]
- Ney, L.J.; Felmingham, K.L.; Bruno, R.; Matthews, A.; Nichols, D.S. Simultaneous quantification of endocannabinoids, oleoylethanolamide and steroid hormones in human plasma and saliva. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1152, 122252. [Google Scholar] [CrossRef] [PubMed]
- Ney, L.J.; Felmingham, K.L.; Bruno, R.; Matthews, A.; Nichols, D.S. Chloroform-based liquid-liquid extraction and LC-MS/MS quantification of endocannabinoids, cortisol and progesterone in human hair. J. Pharm. Biomed. Anal. 2021, 201, 114103. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LLA Maternal Diet | HLA Maternal Diet | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|
| LLA PN Diet | HLA PN Diet | LLA PN Diet | HLA PN Diet | Pmaternal | Ppostnatal | Pint | |
| C14:0 Myristate | 0.077 ± 0.005 | 0.106 ± 0.010 | 0.121 ± 0.021 | 0.083 ± 0.005 | ns | ns | 0.0095 |
| C15:0 Pentadecanoate | 0.023 ± 0.004 | 0.026 ± 0.005 | 0.032 ± 0.005 | 0.025 ± 0.003 | ns | ns | ns |
| C16:0 Dimethylacetal | 5.570 ± 0.507 | 5.468 ± 0.155 | 3.824 ± 0.110 | 4.478 ± 0.252 | 0.001 | ns | ns |
| C16:0 Palmitate | 21.69 ± 0.927 | 23.18 ± 0.927 | 23.47 ± 0.217 | 23.63 ± 0.153 | ns | ns | ns |
| C16:1n7 Palmitoleate | 0.171 ± 0.007 | 0.209 ± 0.013 | 0.208 ± 0.013 | 0.184 ± 0.017 | ns | ns | 0.0239 |
| C17:0 Heptadecanoate | 0.047 ± 0.005 | 0.060 ± 0.007 | 0.065 ± 0.009 | 0.056 ± 0.006 | ns | ns | ns |
| C18:0 Dimethylacetal | 7.840 ± 0.848 | 8.705 ± 1.238 | 4.611 ± 0.536 | 6.193 ± 0.318 | 0.0022 | ns | ns |
| C18:0 Stearate | 20.51 ± 0.336 | 20.24 ± 0.323 | 20.72 ± 0.373 | 21.34 ± 0.282 | ns | ns | ns |
| C18:1n9 Oleate | 14.26 ± 0.443 | 12.57 ± 0.087 | 14.90 ± 0.170 | 14.02 ± 0.431 | 0.0042 | 0.0008 | ns |
| C18:1 Vaccenate | 3.169 ± 0.065 | 2.977 ± 0.046 | 3.447 ± 0.103 | 3.282 ± 0.093 | 0.001 | 0.0251 | ns |
| C18:2n6 Linoleate | 0.211 ± 0.013 | 0.254 ± 0.017 | 0.303 ± 0.034 | 0.330 ± 0.026 | 0.0017 | ns | ns |
| C20:0 Arachidate | 0.078 ± 0.005 | 0.105 ± 0.015 | 0.105 ± 0.013 | 0.113 ± 0.014 | ns | ns | ns |
| C18:3n6 Gamma Linolenate | 0.016 ± 0.005 | 0.016 ± 0.004 | 0.015 ± 0.002 | 0.013 ± 0.003 | ns | ns | ns |
| C20:1n9 Eicosenoate | 0.436 ± 0.018 | 0.408 ± 0.020 | 0.597 ± 0.056 | 0.472 ± 0.041 | 0.0068 | ns | ns |
| C20: 2n6 Eicosadienoate | 0.046 ± 0.008 | 0.059 ± 0.005 | 0.069 ± 0.011 | 0.059 ± 0.006 | ns | ns | ns |
| C22:0 Behenate | 0.126 ± 0.028 | 0.074 ± 0.010 | 0.122 ± 0.011 | 0.109 ± 0.011 | ns | ns | ns |
| C20:3 Homogamma Linolenate | 0.142 ± 0.009 | 0.157 ± 0.012 | 0.183 ± 0.016 | 0.165 ± 0.018 | ns | ns | ns |
| C22:1 Erucate | 0.053 ± 0.009 | 0.045 ± 0.003 | 0.069 ± 0.008 | 0.056 ± 0.010 | ns | ns | ns |
| C20:4n6 Arachidonate | 7.985 ± 0.079 | 8.063 ± 0.082 | 8.610 ± 0.177 | 8.883 ± 0.178 | <0.0001 | ns | ns |
| C24:0 Lignocerate | 0.193 ± 0.024 | 0.156 ± 0.010 | 0.242 ± 0.037 | 0.216 ± 0.018 | 0.0371 | ns | ns |
| C20:5n3 Eicosapentaenoate | 0.010 ± 0.002 | 0.021 ± 0.007 | 0.019 ± 0.004 | 0.006 ± 0.001 | ns | ns | 0.0095 |
| C24:1n9 Nervoniate | 0.094 ± 0.023 | 0.069 ± 0.008 | 0.139 ± 0.009 | 0.097 ± 0.017 | 0.0334 | ns | ns |
| C22:4n6 Adrenate | 2.705 ± 0.063 | 2.877 ± 0.186 | 3.157 ± 0.317 | 2.988 ± 0.196 | ns | ns | ns |
| C22:5n6 Docosapentaenoate | 0.281 ± 0.022 | 0.650 ± 0.107 | 0.372 ± 0.025 | 0.687 ± 0.033 | ns | <0.0001 | ns |
| C22:5n3 Docosapentaenoate | 0.116 ± 0.012 | 0.165 ± 0.019 | 0.154 ± 0.023 | 0.164 ± 0.005 | ns | ns | ns |
| C22:6n3 Docosahexaenoate | 12.64 ± 0.807 | 12.99 ± 0.475 | 13.95 ± 0.284 | 12.62 ± 0.216 | ns | ns | ns |
| LLA Maternal Diet | HLA Maternal Diet | Two-Way ANOVA | |||||
|---|---|---|---|---|---|---|---|
| LLA PN Diet | HLA PN Diet | LLA PN Diet | HLA PN Diet | Pmaternal | Ppostnatal | Pint | |
| C14:0 Myristate | 0.144 ± 0.022 | 0.114 ± 0.013 | 0.144 ± 0.008 | 0.123 ± 0.012 | ns | ns | ns |
| C15:0 Pentadecanoate | 0.034 ± 0.004 | 0.033 ± 0.005 | 0.041 ± 0.007 | 0.029 ± 0.005 | ns | ns | ns |
| C16:0 Dimethylacetal | 3.668 ± 0.491 | 4.778 ± 0.233 | 3.993 ± 0.394 | 4.346 ± 0.338 | ns | ns | ns |
| C16:0 Palmitate | 22.91 ± 0.508 | 23.57 ± 0.710 | 23.57 ± 0.402 | 22.69 ± 0.311 | ns | ns | ns |
| C16:1n7 Palmitoleate | 0.247 ± 0.026 | 0.213 ± 0.013 | 0.231 ± 0.012 | 0.222 ± 0.012 | ns | ns | ns |
| C17:0 Heptadecanoate | 0.073 ± 0.008 | 0.071 ± 0.007 | 0.072 ± 0.005 | 0.064 ± 0.006 | ns | ns | ns |
| C18:0 Dimethylacetal | 4.494 ± 0.746 | 5.887 ± 0.697 | 4.956 ± 0.219 | 5.043 ± 0.463 | ns | ns | ns |
| C18:0 Stearate | 20.10 ± 0.467 | 20.78 ± 0.201 | 20.54 ± 0.252 | 21.19 ± 0.190 | ns | 0.0409 | ns |
| C18:1n9 Oleate | 15.65 ± 0.805 | 13.36 ± 0.661 | 14.02 ± 0.320 | 14.80 ± 0.158 | ns | ns | 0.0221 |
| C18:1 Vaccenate | 3.479 ± 0.112 | 3.235 ± 0.099 | 3.319 ± 0.086 | 3.418 ± 0.051 | ns | ns | ns |
| C18:2n6 Linoleate | 0.295 ± 0.044 | 0.292 ± 0.009 | 0.264 ± 0.009 | 0.292 ± 0.014 | ns | ns | ns |
| C20:0 Arachidate | 0.138 ± 0.014 | 0.087 ± 0.005 | 0.103 ± 0.006 | 0.116 ± 0.007 | ns | ns | 0.003 |
| C18:3n6 Gamma Linolenate | 0.013 ± 0.002 | 0.013 ± 0.003 | 0.016 ± 0.003 | 0.009 ± 0.001 | ns | ns | ns |
| C20:1n9 Eicosenoate | 0.532 ± 0.043 | 0.506 ± 0.047 | 0.483 ± 0.050 | 0.592 ± 0.027 | ns | ns | ns |
| C20: 2n6 Eicosadienoate | 0.079 ± 0.013 | 0.064 ± 0.007 | 0.037 ± 0.005 | 0.056 ± 0.004 | 0.0083 | ns | ns |
| C22:0 Behenate | 0.127 ± 0.013 | 0.114 ± 0.018 | 0.104 ± 0.012 | 0.119 ± 0.007 | ns | ns | ns |
| C20:3 Homogamma Linolenate | 0.212 ± 0.024 | 0.218 ± 0.019 | 0.237 ± 0.016 | 0.199 ± 0.010 | ns | ns | ns |
| C22:1 Erucate | 0.072 ± 0.012 | 0.068 ± 0.007 | 0.062 ± 0.008 | 0.077 ± 0.010 | ns | ns | ns |
| C20:4n6 Arachidonate | 8.135 ± 0.185 | 8.150 ± 0.117 | 8.374 ± 0.281 | 8.378 ± 0.187 | ns | ns | ns |
| C24:0 Lignocerate | 0.221 ± 0.017 | 0.205 ± 0.012 | 0.216 ± 0.024 | 0.235 ± 0.017 | ns | ns | ns |
| C20:5n3 Eicosapentaenoate | 0.019 ± 0.005 | 0.012 ± 0.002 | 0.013 ± 0.002 | 0.014 ± 0.001 | ns | ns | ns |
| C24:1n9 Nervoniate | 0.127 ± 0.021 | 0.096 ± 0.008 | 0.117 ± 0.012 | 0.122 ± 0.009 | ns | ns | ns |
| C22:4n6 Adrenate | 3.047 ± 0.207 | 3.094 ± 0.126 | 3.287 ± 0.118 | 3.356 ± 0.156 | ns | ns | ns |
| C22:5n6 Docosapentaenoate | 0.349 ± 0.046 | 0.568 ± 0.017 | 0.299 ± 0.020 | 0.621 ± 0.055 | ns | <0.0001 | ns |
| C22:5n3 Docosapentaenoate | 0.146 ± 0.021 | 0.173 ± 0.027 | 0.199 ± 0.015 | 0.168 ± 0.021 | ns | ns | ns |
| C22:6n3 Docosahexaenoate | 14.20 ± 0.601 | 14.23 ± 0.669 | 16.22 ± 0.823 | 13.99 ± 0.375 | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ezechukwu, H.C.; Ney, L.J.; Jarvis, M.A.; Shrestha, N.; Holland, O.J.; Cuffe, J.S.M.; Perkins, A.V.; Yau, S.-Y.; McAinch, A.J.; Hryciw, D.H. Sex-Specific Changes to Brain Fatty Acids, Plasmalogen, and Plasma Endocannabinoids in Offspring Exposed to Maternal and Postnatal High-Linoleic-Acid Diets. Int. J. Mol. Sci. 2024, 25, 7911. https://doi.org/10.3390/ijms25147911
Ezechukwu HC, Ney LJ, Jarvis MA, Shrestha N, Holland OJ, Cuffe JSM, Perkins AV, Yau S-Y, McAinch AJ, Hryciw DH. Sex-Specific Changes to Brain Fatty Acids, Plasmalogen, and Plasma Endocannabinoids in Offspring Exposed to Maternal and Postnatal High-Linoleic-Acid Diets. International Journal of Molecular Sciences. 2024; 25(14):7911. https://doi.org/10.3390/ijms25147911
Chicago/Turabian StyleEzechukwu, Henry C., Luke J. Ney, Madeline A. Jarvis, Nirajan Shrestha, Olivia J. Holland, James S. M. Cuffe, Anthony V. Perkins, Suk-Yu Yau, Andrew J. McAinch, and Deanne H. Hryciw. 2024. "Sex-Specific Changes to Brain Fatty Acids, Plasmalogen, and Plasma Endocannabinoids in Offspring Exposed to Maternal and Postnatal High-Linoleic-Acid Diets" International Journal of Molecular Sciences 25, no. 14: 7911. https://doi.org/10.3390/ijms25147911