
Heteromers Formed by GPR55 and Either Cannabinoid CB1 or CB2 Receptors Are Upregulated in the Prefrontal Cortex of Multiple Sclerosis Patients

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
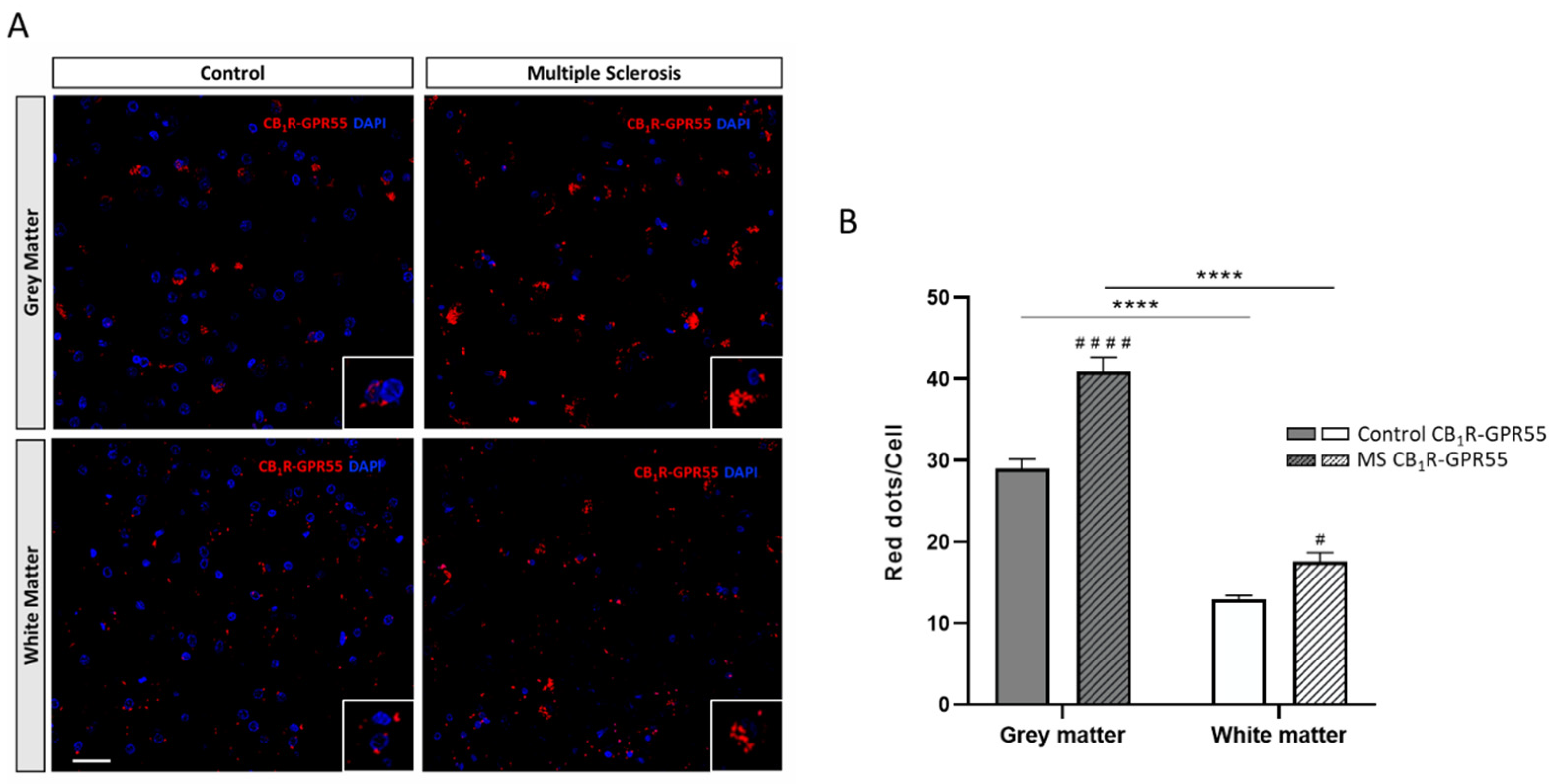
2.1. Expression of CB1R-GPR55 Heteromers in Controls and MS Patients
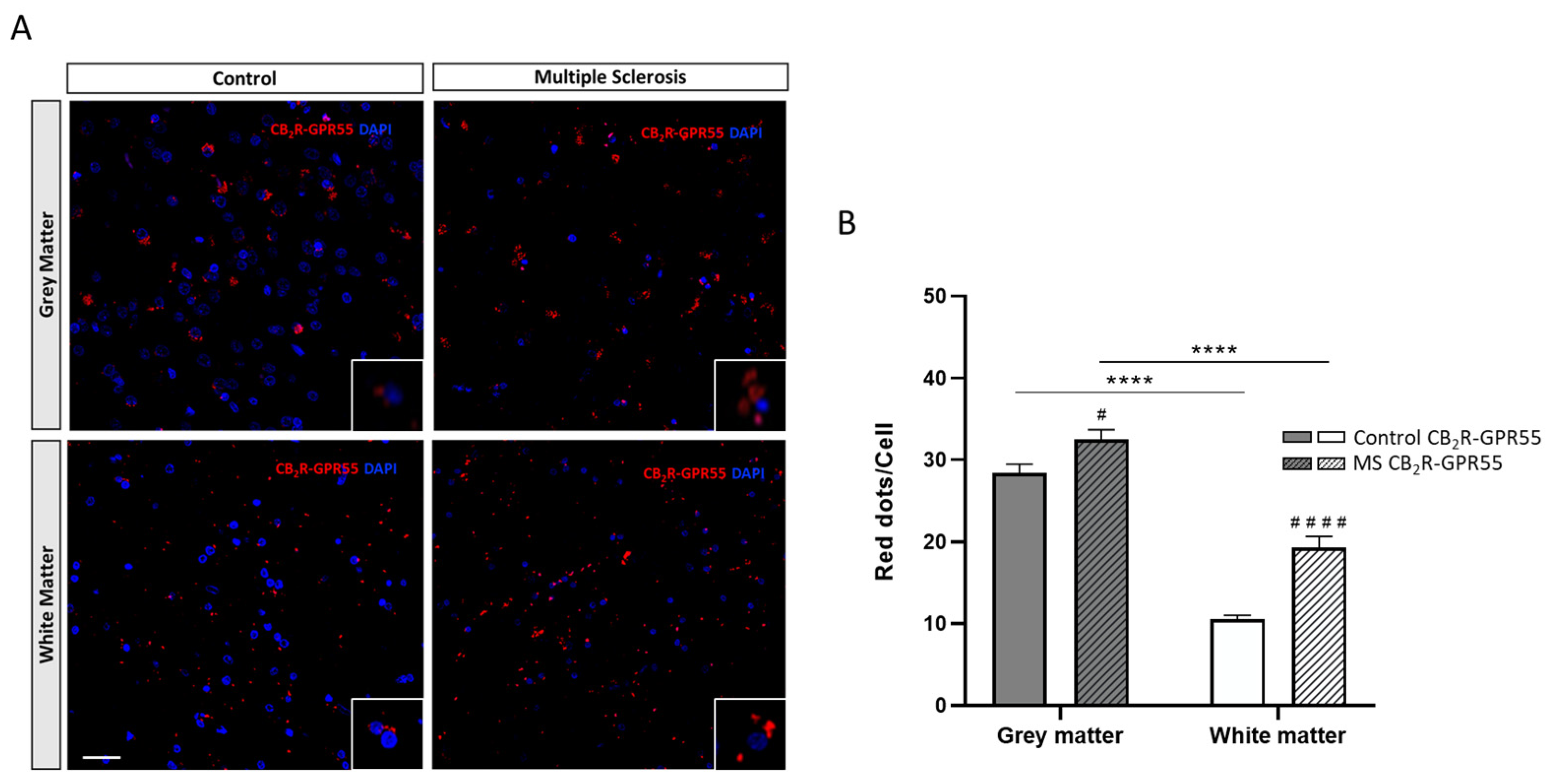
2.2. Expression of CB2R-GPR55 Heteromers in Controls and MS Patients
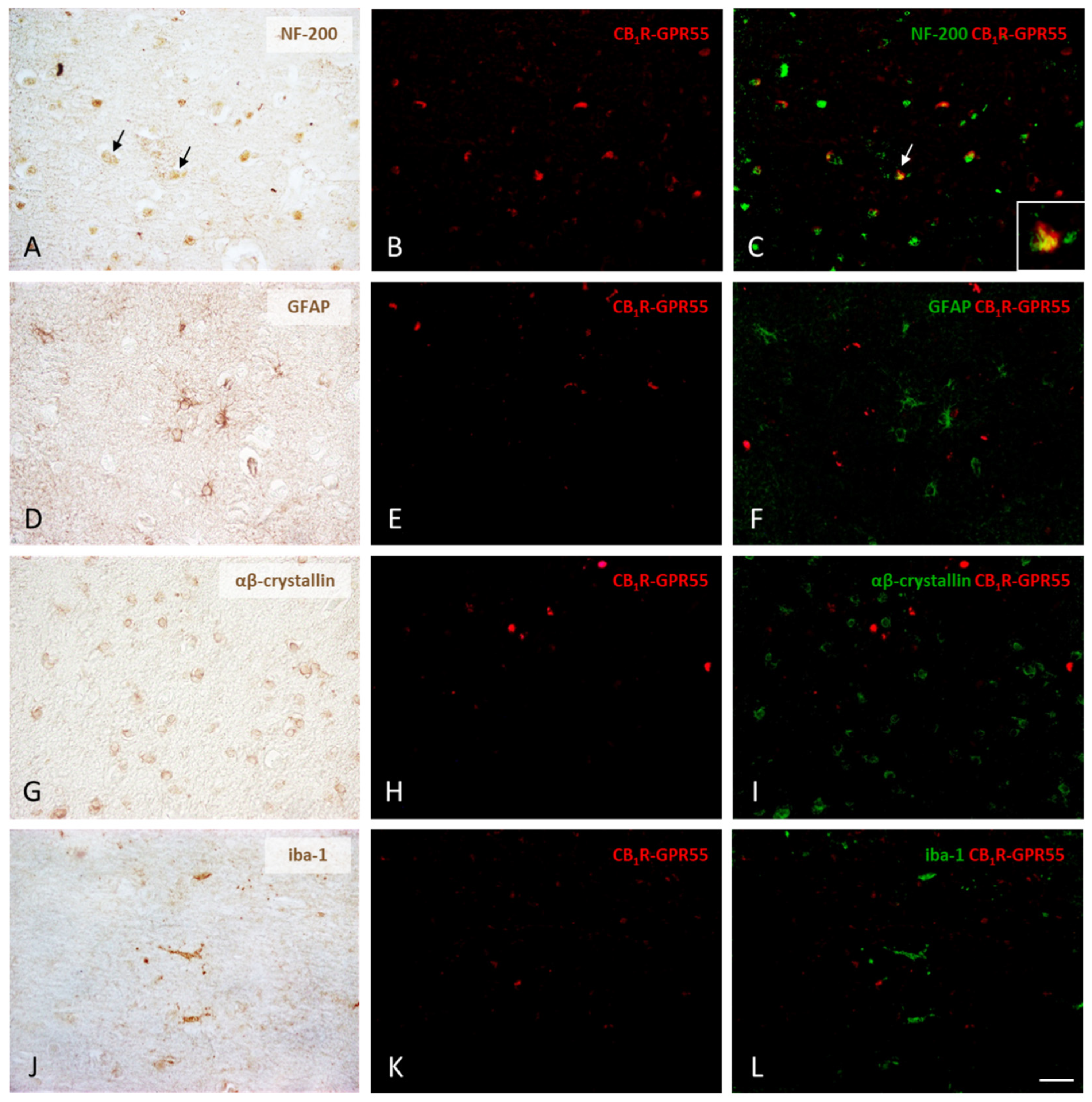
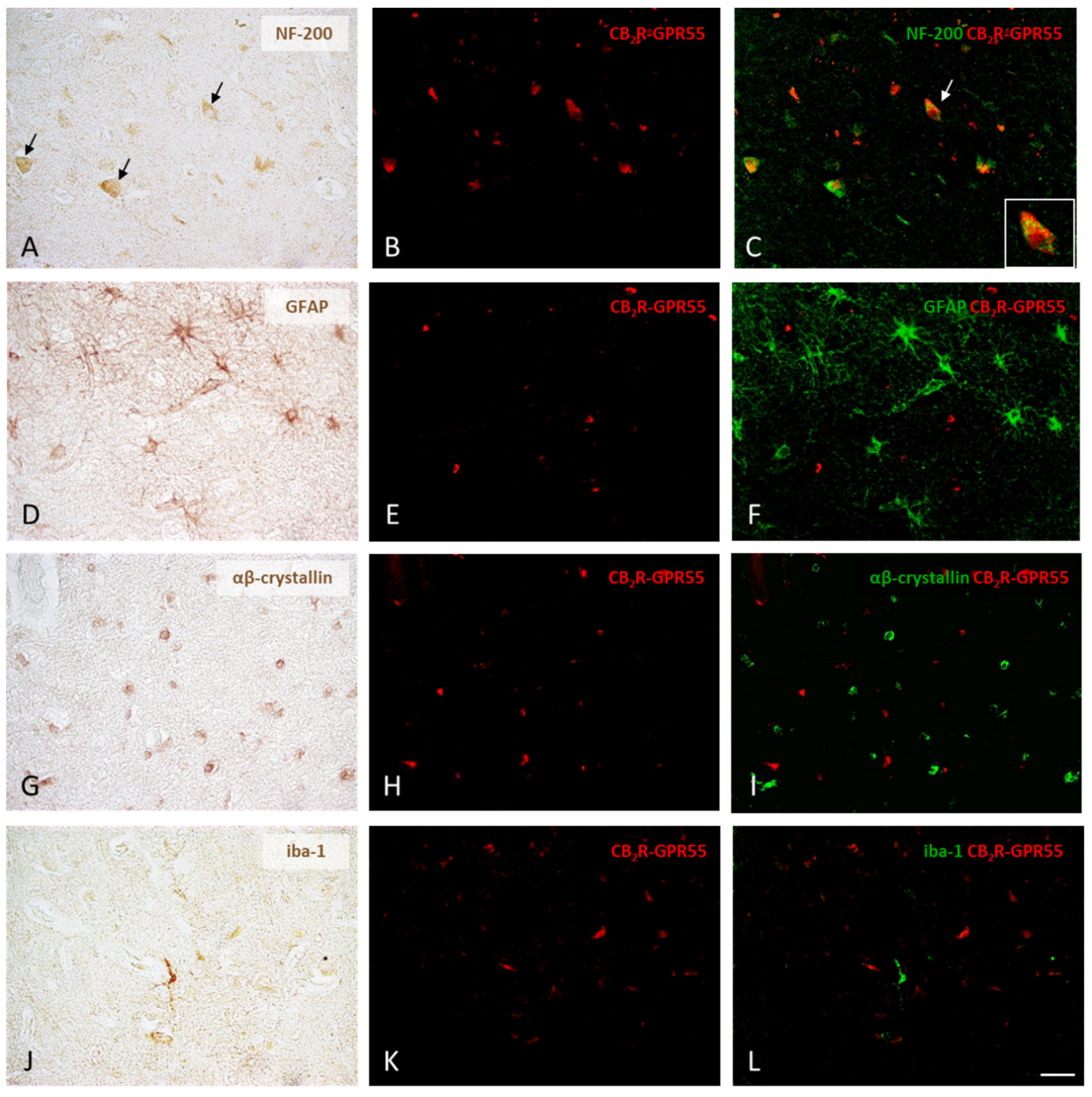
2.3. Expression of CB1R-GPR55 and CB2R-GPR55 Heteromers in Neurons and Different Types of Glial Cells
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. In Situ Proximity Ligation Assay (PLA)
4.3. Co-Staining Combining PLA and Immunohistochemistry
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walton, C.; King, R.; Rechtman, L.; Kaye, W.; Leray, E.; Marrie, R.A.; Robertson, N.; La Rocca, N.; Uitdehaag, B.; van der Mei, I.; et al. Rising prevalence of multiple sclerosis worldwide: Insights from the Atlas of MS, third edition. Mult. Scler. J. 2020, 26, 1816–1821. [Google Scholar] [CrossRef]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and Treatment of Multiple Sclerosis: A Review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef]
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef]
- Cao, Y.; Diao, W.; Tian, F.; Zhang, F.; He, L.; Long, X.; Zhou, F.; Jia, Z. Gray Matter Atrophy in the Cortico-Striatal-Thalamic Network and Sensorimotor Network in Relapsing–Remitting and Primary Progressive Multiple Sclerosis. Neuropsychol. Rev. 2021, 31, 703–720. [Google Scholar] [CrossRef]
- Calabrese, M.; Magliozzi, R.; Ciccarelli, O.; Geurts, J.J.G.; Reynolds, R.; Martin, R. Exploring the origins of grey matter damage in multiple sclerosis. Nat. Rev. Neurosci. 2015, 16, 147–158. [Google Scholar] [CrossRef]
- Oh, J.; Vidal-Jordana, A.; Montalban, X. Multiple sclerosis: Clinical aspects. Curr. Opin. Neurol. 2018, 31, 752–759. [Google Scholar] [CrossRef]
- Ford, H. Clinical presentation and diagnosis of multiple sclerosis. Clin. Med. J. R. Coll. Physicians Lond. 2020, 20, 380–383. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sørensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef]
- Hegen, H.; Berek, K.; Deisenhammer, F. Cerebrospinal fluid kappa free light chains as biomarker in multiple sclerosis—From diagnosis to prediction of disease activity. Wien. Med. Wochenschr. 2022, 172, 337–345. [Google Scholar] [CrossRef]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- International Multiple Sclerosis Genetics Consortium; Harroud, A.; Stridh, P.; McCauley, J.L.; Saarela, J.; Bosch, A.M.R.v.D.; Engelenburg, H.J.; Beecham, A.H.; Alfredsson, L.; Alikhani, K.; et al. Locus for severity implicates CNS resilience in progression of multiple sclerosis. Nature 2023, 619, 323–331. [Google Scholar] [CrossRef]
- Goodin, D.S. Glucocorticoid treatment of multiple sclerosis. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 122, pp. 455–464. [Google Scholar]
- Krajnc, N.; Bsteh, G.; Berger, T.; Mares, J.; Hartung, H.P. Monoclonal Antibodies in the Treatment of Relapsing Multiple Sclerosis: An Overview with Emphasis on Pregnancy, Vaccination, and Risk Management. Neurotherapeutics 2022, 19, 753–773. [Google Scholar] [CrossRef]
- McGinley, M.P.; Cohen, J.A. Sphingosine 1-phosphate receptor modulators in multiple sclerosis and other conditions. Lancet 2021, 398, 1184–1194. [Google Scholar] [CrossRef]
- Conte, A.; Silván, C.V. Review of Available Data for the Efficacy and Effectiveness of Nabiximols Oromucosal Spray (Sativex®) in Multiple Sclerosis Patients with Moderate to Severe Spasticity. Neurodegener. Dis. 2022, 21, 55–62. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Gómez-Ruiz, M.; García, C.; Hernández, M.; Ramos, J.A. Modeling Neurodegenerative Disorders for Developing Cannabinoid-Based Neuroprotective Therapies. Methods Enzymol. 2017, 593, 175–198. [Google Scholar]
- Mecha, M.; Carrillo-Salinas, F.J.; Feliú, A.; Mestre, L.; Guaza, C. Perspectives on Cannabis-Based Therapy of Multiple Sclerosis: A Mini-Review. Front. Cell. Neurosci. 2020, 14, 34. [Google Scholar] [CrossRef]
- Nouh, R.A.; Kamal, A.; Abdelnaser, A. Cannabinoids and Multiple Sclerosis: A Critical Analysis of Therapeutic Potentials and Safety Concerns. Pharmaceutics 2023, 15, 1151. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Giuffrida, A.; Seillier, A. New insights on endocannabinoid transmission in psychomotor disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 38, 51–58. [Google Scholar] [CrossRef]
- Aymerich, M.S.; Aso, E.; Abellanas, M.A.; Tolon, R.M.; Ramos, J.A.; Ferrer, I.; Romero, J.; Fernández-Ruiz, J. Cannabinoid pharmacology/therapeutics in chronic degenerative disorders affecting the central nervous system. Biochem. Pharmacol. 2018, 157, 67–84. [Google Scholar] [CrossRef]
- Shu-Jung Hu, S.; Mackie, K. Distribution of the endocannabinoid system in the central nervous system. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2015; Volume 231, pp. 59–93. [Google Scholar]
- Glass, M.; Dragunow, M.; Faull, R.L.M. Cannabinoid receptors in the human brain: A detailed anatomical and quantitative autoradiographic study in the fetal, neonatal and adult human brain. Neuroscience 1997, 77, 299–318. [Google Scholar] [CrossRef]
- Chou, S.; Ranganath, T.; Fish, K.N.; Lewis, D.A.; Sweet, R.A. Cell type specific cannabinoid CB1 receptor distribution across the human and non-human primate cortex. Sci. Rep. 2022, 12, 9605. [Google Scholar] [CrossRef]
- Martínez-Pinilla, E.; Rico, A.J.; Rivas-Santisteban, R.; Lillo, J.; Roda, E.; Navarro, G.; Franco, R.; Lanciego, J.L. Expression of cannabinoid CB1 R–GPR55 heteromers in neuronal subtypes of the Macaca fascicularis striatum. Ann. N. Y. Acad. Sci. 2020, 1475, 34–42. [Google Scholar] [CrossRef]
- Cabral, G.A.; Ferreira, G.A.; Jamerson, M.J. Endocannabinoids and the immune system in health and disease. In Endocannabinoids. Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2015; Volume 231, pp. 185–211. ISBN 9783319208251. [Google Scholar]
- Onaivi, E.S. Neuropsychobiological evidence for the functional presence and expression of cannabinoid CB2 receptors in the brain. Neuropsychobiology 2007, 54, 231–246. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the presence and functional expression of cannabinoid CB2 receptors in brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef]
- Lanciego, J.L.; Barroso-Chinea, P.; Rico, A.J.; Conte-Perales, L.; Callén, L.; Roda, E.; Gómez-Bautista, V.; López, I.P.; Lluis, C.; Labandeira-García, J.L.; et al. Expression of the mRNA coding the cannabinoid receptor 2 in the pallidal complex of Macaca fascicularis. J. Psychopharmacol. 2011, 25, 97–104. [Google Scholar] [CrossRef]
- Martínez-Pinilla, E.; Rico, A.J.; Rivas-Santisteban, R.; Lillo, J.; Roda, E.; Navarro, G.; Lanciego, J.L.; Franco, R. Expression of GPR55 and either cannabinoid CB1 or CB2 heteroreceptor complexes in the caudate, putamen, and accumbens nuclei of control, parkinsonian, and dyskinetic non-human primates. Brain Struct. Funct. 2020, 225, 2153–2164. [Google Scholar] [CrossRef]
- Liu, Q.R.; Pan, C.H.; Hishimoto, A.; Li, C.Y.; Xi, Z.X.; Llorente-Berzal, A.; Viveros, M.P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; et al. Species differences in cannabinoid receptor 2 (CNR2 gene): Identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar] [CrossRef]
- Brusco, A.; Tagliaferro, P.A.; Saez, T.; Onaivi, E.S. Ultrastructural localization of neuronal brain CB2 cannabinoid receptors. Ann. N. Y. Acad. Sci. 2008, 1139, 450–457. [Google Scholar] [CrossRef]
- Brusco, A.; Tagliaferro, P.; Saez, T.; Onaivi, E.S. Postsynaptic localization of CB2 cannabinoid receptors in the rat hippocampus. Synapse 2008, 62, 944–949. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Moro, M.A.; Martínez-Orgado, J. Cannabinoids in Neurodegenerative Disorders and Stroke/Brain Trauma: From Preclinical Models to Clinical Applications. Neurotherapeutics 2015, 12, 793–806. [Google Scholar] [CrossRef]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Baker, D.; Pryce, G.; Davies, W.L.; Hiley, C.R. In silico patent searching reveals a new cannabinoid receptor. Trends Pharmacol. Sci. 2006, 27, 1–4. [Google Scholar] [CrossRef]
- Oka, S.; Nakajima, K.; Yamashita, A.; Kishimoto, S.; Sugiura, T. Identification of GPR55 as a lysophosphatidylinositol receptor. Biochem. Biophys. Res. Commun. 2007, 362, 928–934. [Google Scholar] [CrossRef]
- Oyagawa, C.R.M.; Grimsey, N.L. Cannabinoid receptor CB1 and CB2 interacting proteins: Techniques, progress and perspectives. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 166, pp. 83–132. ISBN 9780128233511. [Google Scholar]
- Pietr, M.; Kozela, E.; Levy, R.; Rimmerman, N.; Lin, Y.H.; Stella, N.; Vogel, Z.; Juknat, A. Differential changes in GPR55 during microglial cell activation. FEBS Lett. 2009, 583, 2071–2076. [Google Scholar] [CrossRef]
- Wu, C.S.; Zhu, J.; Wager-Miller, J.; Wang, S.; O’Leary, D.; Monory, K.; Lutz, B.; MacKie, K.; Lu, H.C. Requirement of cannabinoid CB1 receptors in cortical pyramidal neurons for appropriate development of corticothalamic and thalamocortical projections. Eur. J. Neurosci. 2010, 32, 693–706. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Marrion, N.V.; Peters, J.A.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: G protein-coupled receptors. Br. J. Pharmacol. 2017, 174, S17–S129. [Google Scholar] [CrossRef]
- Krishna Kumar, K.; Shalev-Benami, M.; Robertson, M.J.; Hu, H.; Banister, S.D.; Hollingsworth, S.A.; Latorraca, N.R.; Kato, H.E.; Hilger, D.; Maeda, S.; et al. Structure of a Signaling Cannabinoid Receptor 1-G Protein Complex. Cell 2019, 176, 448–458.e12. [Google Scholar] [CrossRef]
- Lynch, D.L.; Hurst, D.P.; Reggio, P.H. The Nucleotide-Free State of the Cannabinoid CB2/Gi Complex. Cell 2020, 180, 603–604. [Google Scholar] [CrossRef]
- Henstridge, C.M.; Balenga, N.A.B.; Ford, L.A.; Ross, R.A.; Waldhoer, M.; Irving, A.J. The GPR55 ligand L-α-lysophosphatidylinositol promotes RhoA-dependent Ca2+ signaling and NFAT activation. FASEB J. 2009, 23, 183–193. [Google Scholar] [CrossRef]
- Henstridge, C.M.; Balenga, N.A.; Schröder, R.; Kargl, J.K.; Platzer, W.; Martini, L.; Arthur, S.; Penman, J.; Whistler, J.L.; Kostenis, E.; et al. GPR55 ligands promote receptor coupling to multiple signalling pathways. Br. J. Pharmacol. 2010, 160, 604–614. [Google Scholar] [CrossRef]
- Ferré, S.; Baler, R.; Bouvier, M.; Caron, M.G.; Devi, L.A.; Durroux, T.; Fuxe, K.; George, S.R.; Javitch, J.A.; Lohse, M.J.; et al. Building a new conceptual framework for receptor heteromers. Nat. Chem. Biol. 2009, 5, 131–134. [Google Scholar] [CrossRef]
- Franco, R.; Martínez-Pinilla, E.; Lanciego, J.L.; Navarro, G. Basic pharmacological and structural evidence for class A G-protein-coupled receptor heteromerization. Front. Pharmacol. 2016, 7, 76. [Google Scholar] [CrossRef]
- Franco, R.; Aguinaga, D.; Jiménez, J.; Lillo, J.; Martínez-Pinilla, E.; Navarro, G. Biased receptor functionality versus biased agonism in G-protein-coupled receptors. Biomol. Concepts 2018, 9, 143–154. [Google Scholar] [CrossRef]
- Balenga, N.A.; Martínez-Pinilla, E.; Kargl, J.; Schröder, R.; Peinhaupt, M.; Platzer, W.; Bálint, Z.; Zamarbide, M.; Dopeso-Reyes, I.G.; Ricobaraza, A.; et al. Heteromerization of GPR55 and cannabinoid CB2 receptors modulates signalling. Br. J. Pharmacol. 2014, 171, 5387–5406. [Google Scholar] [CrossRef]
- Martínez-Pinilla, E.; Aguinaga, D.; Navarro, G.; Rico, A.J.; Oyarzábal, J.; Sánchez-Arias, J.A.; Lanciego, J.L.; Franco, R. Targeting CB1 and GPR55 Endocannabinoid Receptors as a Potential Neuroprotective Approach for Parkinson’s Disease. Mol. Neurobiol. 2019, 56, 5900–5910. [Google Scholar] [CrossRef]
- Martínez-Pinilla, E.; Reyes-Resina, I.; Oñatibia-Astibia, A.; Zamarbide, M.; Ricobaraza, A.; Navarro, G.; Moreno, E.; Dopeso-Reyes, I.G.; Sierra, S.; Rico, A.J.; et al. CB1 and GPR55 receptors are co-expressed and form heteromers in rat and monkey striatum. Exp. Neurol. 2014, 261, 44–52. [Google Scholar] [CrossRef]
- Tomas-Roig, J.; Agbemenyah, H.Y.; Celarain, N.; Quintana, E.; Ramió-Torrentà, L.; Havemann-Reinecke, U. Dose-dependent effect of cannabinoid WIN-55,212-2 on myelin repair following a demyelinating insult. Sci. Rep. 2020, 10, 590. [Google Scholar] [CrossRef]
- Shao, B.Z.; Wei, W.; Ke, P.; Xu, Z.Q.; Zhou, J.X.; Liu, C. Activating cannabinoid receptor 2 alleviates pathogenesis of experimental autoimmune encephalomyelitis via activation of autophagy and inhibiting NLRP3 inflammasome. CNS Neurosci. Ther. 2014, 20, 1021–1028. [Google Scholar] [CrossRef]
- Paloczi, J.; Varga, Z.V.; Hasko, G.; Pacher, P. Neuroprotection in Oxidative Stress-Related Neurodegenerative Diseases: Role of Endocannabinoid System Modulation. Antioxid. Redox Signal. 2018, 29, 75–108. [Google Scholar] [CrossRef]
- Kozela, E.; Lev, N.; Kaushansky, N.; Eilam, R.; Rimmerman, N.; Levy, R.; Ben-Nun, A.; Juknat, A.; Vogel, Z. Cannabidiol inhibits pathogenic T cells, decreases spinal microglial activation and ameliorates multiple sclerosis-like disease in C57BL/6 mice. Br. J. Pharmacol. 2011, 163, 1507–1519. [Google Scholar] [CrossRef]
- Fujiwara, M.; Egashira, N. New perspectives in the studies on endocannabinoid and cannabis: Abnormal behaviors associate with CB1 cannabinoid receptor and development of therapeutic application. J. Pharmacol. Sci. 2004, 96, 362–366. [Google Scholar] [CrossRef]
- Pryce, G.; Ahmed, Z.; Hankey, D.J.R.; Jackson, S.J.; Croxford, J.L.; Pocock, J.M.; Ledent, C.; Petzold, A.; Thompson, A.J.; Giovannoni, G.; et al. Cannabinoids inhibit neurodegeneration in models of multiple sclerosis. Brain 2003, 126, 2191–2202. [Google Scholar] [CrossRef]
- Navarro, G.; Borroto-Escuela, D.; Angelats, E.; Etayo, I.; Reyes-Resina, I.; Pulido-Salgado, M.; Rodríguez-Pérez, A.; Canela, E.; Saura, J.; Lanciego, J.L.; et al. Receptor-heteromer mediated regulation of endocannabinoid signaling in activated microglia. Role of CB1 and CB2 receptors and relevance for Alzheimer’s disease and levodopa-induced dyskinesia. Brain. Behav. Immun. 2018, 67, 139–151. [Google Scholar] [CrossRef]
- McKenna, M.; McDougall, J.J. Cannabinoid control of neurogenic inflammation. Br. J. Pharmacol. 2020, 177, 4386–4399. [Google Scholar] [CrossRef]
- Tao, Y.; Li, L.; Jiang, B.; Feng, Z.; Yang, L.; Tang, J.; Chen, Q.; Zhang, J.; Tan, Q.; Feng, H.; et al. Cannabinoid receptor-2 stimulation suppresses neuroinflammation by regulating microglial M1/M2 polarization through the cAMP/PKA pathway in an experimental GMH rat model. Brain. Behav. Immun. 2016, 58, 118–129. [Google Scholar] [CrossRef]
- Martínez-pinilla, E.; Rubio-sardón, N.; Villar-conde, S.; Navarro, G.; Del Valle, E.; Tolivia, J.; Franco, R.; Navarro, A. Cuprizone-induced neurotoxicity in human neural cell lines is mediated by a reversible mitochondrial dysfunction: Relevance for demyelination models. Brain Sci. 2021, 11, 272. [Google Scholar] [CrossRef]
- Urbi, B.; Corbett, J.; Hughes, I.; Owusu, M.A.; Thorning, S.; Broadley, S.A.; Sabet, A.; Heshmat, S. Effects of Cannabis in Parkinson’s Disease: A Systematic Review and Meta-Analysis. J. Parkinsons. Dis. 2022, 12, 495–508. [Google Scholar] [CrossRef]
- Varshney, K.; Patel, A.; Ansari, S.; Shet, P.; Panag, S.S. Cannabinoids in Treating Parkinson’s Disease Symptoms: A Systematic Review of Clinical Studies. Cannabis Cannabinoid Res. 2023, 8, 716–730. [Google Scholar] [CrossRef]
- Christensen, D.B.; Roth, M.; Trygstad, T.; Byrd, J. Evaluation of a pilot medication therapy management project within the North Carolina State Health Plan. J. Am. Pharm. Assoc. 2007, 47, 471–483. [Google Scholar] [CrossRef]
- Tolivia, J.; Navarro, A.; Del Valle, E.; Perez, C.; Ordoñez, C.; Martínez, E. Application of photoshop and scion image analysis to quantification of signals in histochemistry, immunocytochemistry and hybridocytochemistry. Anal. Quant. Cytol. Histol. 2006, 28, 43–53. [Google Scholar]
- Navarro, A.; Ordóñez, C.; Martínez, E.; Pérez, C.; Astudillo, A.; Tolivia, J. Apolipoprotein d expression absence in degenerating neurons of human central nervous system. Histol. Histopathol. 2008, 23, 995–1001. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Sex | Age | Type of MS | Brain Area | Post-Mortem Time | Brain Bank |
|---|---|---|---|---|---|---|
| 1 | Male | 58 | MS | Prefrontal cortex | 6–12 h | Central Nervous Tissue Bank Madrid (CIEN Foundation) |
| 2 | Male | 38 | SPMS | Prefrontal cortex | 6–12 h | University Central Hospital of Asturias (HUCA) |
| 3 | Female | 47 | PPMS | Prefrontal cortex | 6–12 h | Center for Biomedical Research of Navarra (NAVARRABIOMED) |
| 4 | Female | 48 | MS | Prefrontal cortex | 6–12 h | Southern Galicia Health Research Institute (IISGS) |
| 5 | Male | 38 | Control | Prefrontal cortex | 6–12 h | University Central Hospital of Asturias (HUCA) |
| 6 | Male | 66 | Control | Prefrontal cortex | 6–12 h | Center for Biomedical Research of Navarra (NAVARRABIOMED) |
| 7 | Male | 62 | Control | Prefrontal cortex | 6–12 h | Center for Biomedical Research of Navarra (NAVARRABIOMED) |
| 8 | Female | 52 | Control | Prefrontal cortex | 6–12 h | University Central Hospital of Asturias (HUCA) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menéndez-Pérez, C.; Rivas-Santisteban, R.; del Valle, E.; Tolivia, J.; Navarro, A.; Franco, R.; Martínez-Pinilla, E. Heteromers Formed by GPR55 and Either Cannabinoid CB1 or CB2 Receptors Are Upregulated in the Prefrontal Cortex of Multiple Sclerosis Patients. Int. J. Mol. Sci. 2024, 25, 4176. https://doi.org/10.3390/ijms25084176
Menéndez-Pérez C, Rivas-Santisteban R, del Valle E, Tolivia J, Navarro A, Franco R, Martínez-Pinilla E. Heteromers Formed by GPR55 and Either Cannabinoid CB1 or CB2 Receptors Are Upregulated in the Prefrontal Cortex of Multiple Sclerosis Patients. International Journal of Molecular Sciences. 2024; 25(8):4176. https://doi.org/10.3390/ijms25084176
Chicago/Turabian StyleMenéndez-Pérez, Carlota, Rafael Rivas-Santisteban, Eva del Valle, Jorge Tolivia, Ana Navarro, Rafael Franco, and Eva Martínez-Pinilla. 2024. "Heteromers Formed by GPR55 and Either Cannabinoid CB1 or CB2 Receptors Are Upregulated in the Prefrontal Cortex of Multiple Sclerosis Patients" International Journal of Molecular Sciences 25, no. 8: 4176. https://doi.org/10.3390/ijms25084176