
Exogenous GA3 Enhances Nitrogen Uptake and Metabolism under Low Nitrate Conditions in ‘Duli’ (Pyrus betulifolia Bunge) Seedlings
Abstract
:1. Introduction
2. Results
2.1. Effects of GA3 and PAC on the Growth, Chlorophyll Content, and Root Length of ‘Duli’
2.2. Effects of GA3 on the Growth and Chlorophyll Content of ‘Duli’ Seedlings under NO3− Deficiency
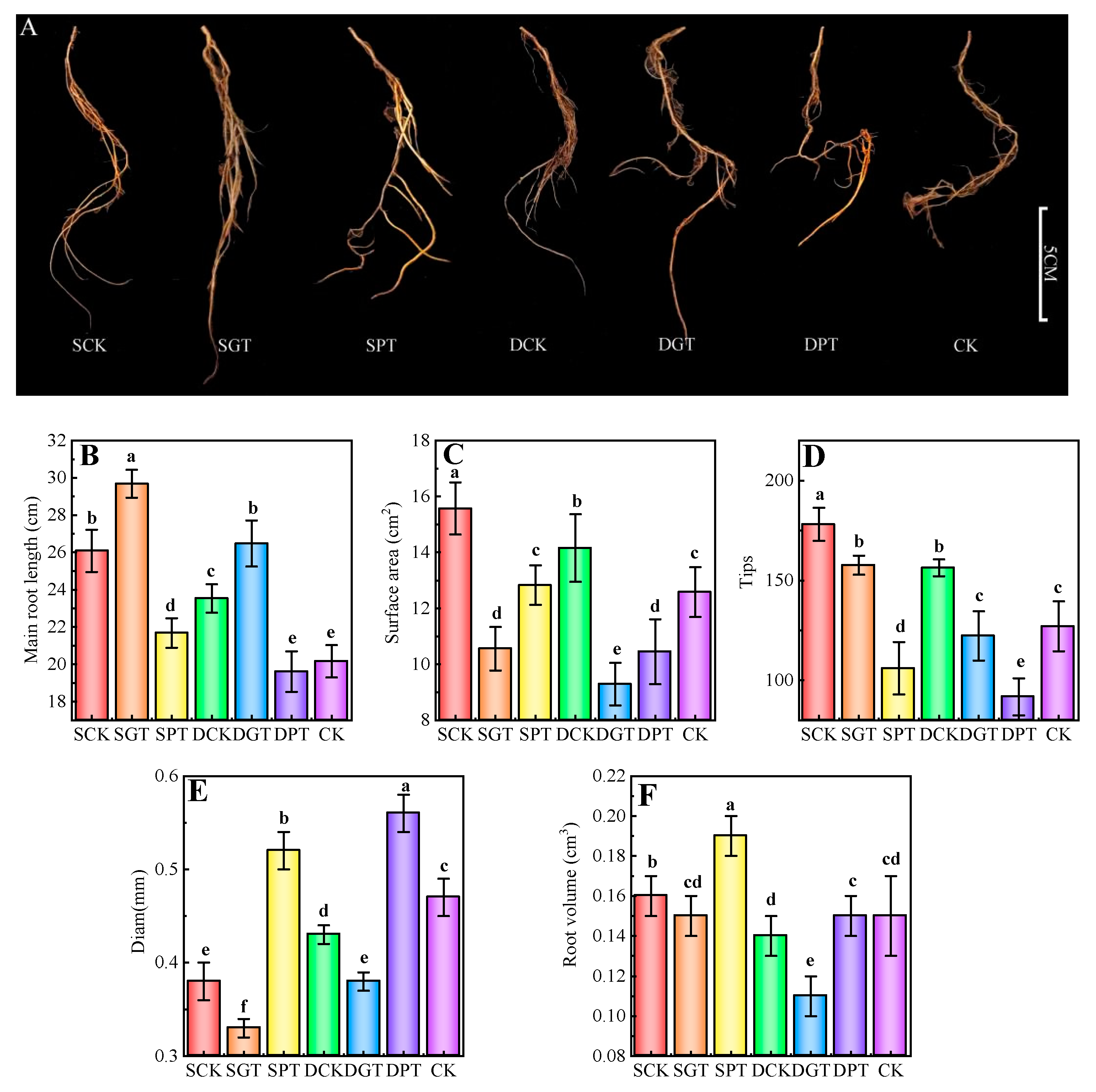
2.3. Effects of Exogenous GA3 Application on the Root Architecture of ‘Duli’ under NO3− Deficiency
2.4. Effects of Exogenous GA3 on the Photosynthetic Parameters of ‘Duli’ under NO3− Deficiency
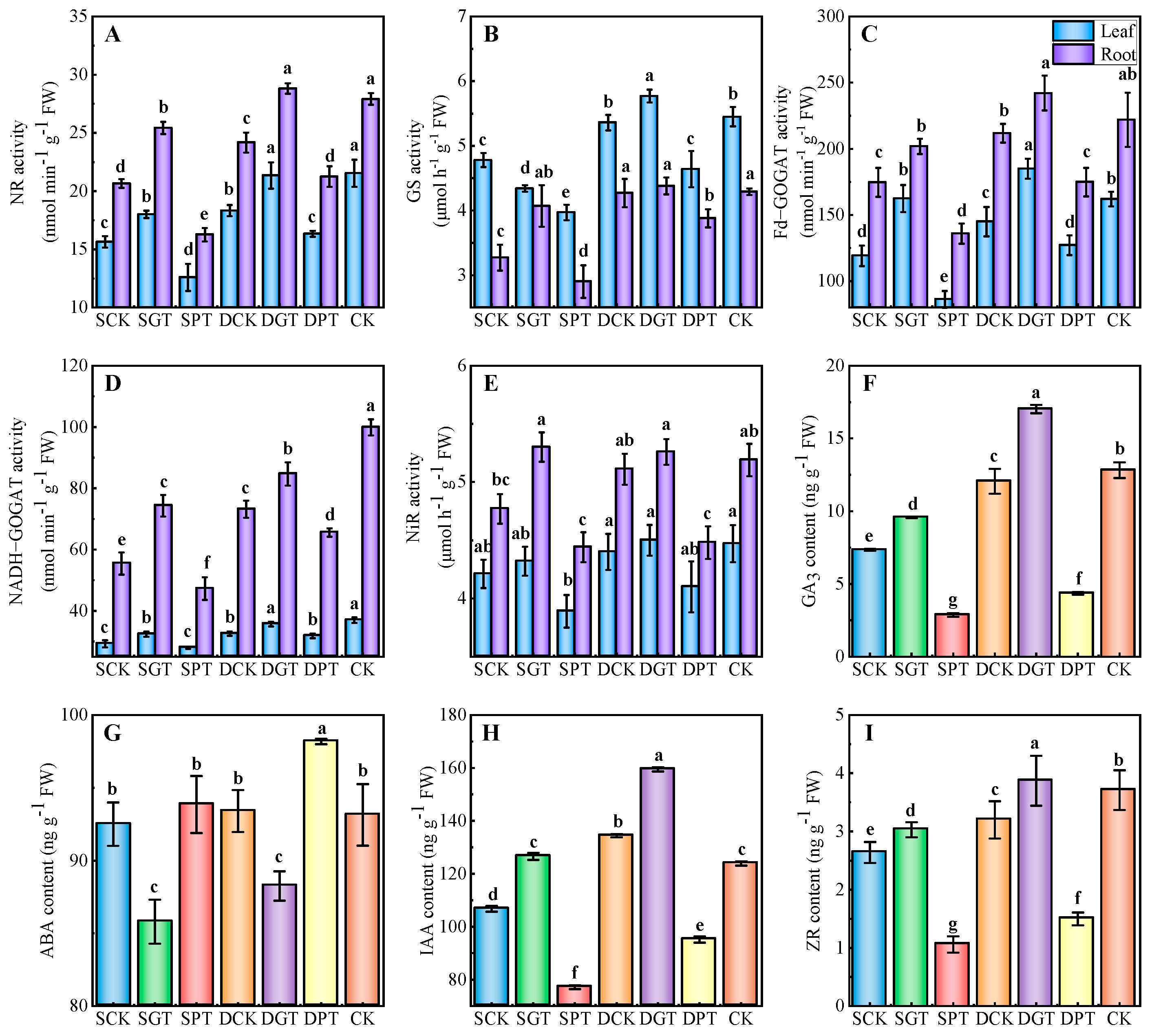
2.5. Effects of Exogenous GA3 on the Activities of N−Metabolizing Enzymes and Content of Endogenous Hormones in ‘Duli’ under NO3− Deficiency
2.6. Effects of Exogenous GA3 Application on the Content of Mineral Elements in ‘Duli’ under NO3− Deficiency
2.7. Effects of Exogenous GA3 Application on the Expression of N Uptake and Metabolism−Related Genes in ‘Duli’ under NO3− Deficiency
2.8. Correlation and Principal Component Analysis (PCA)
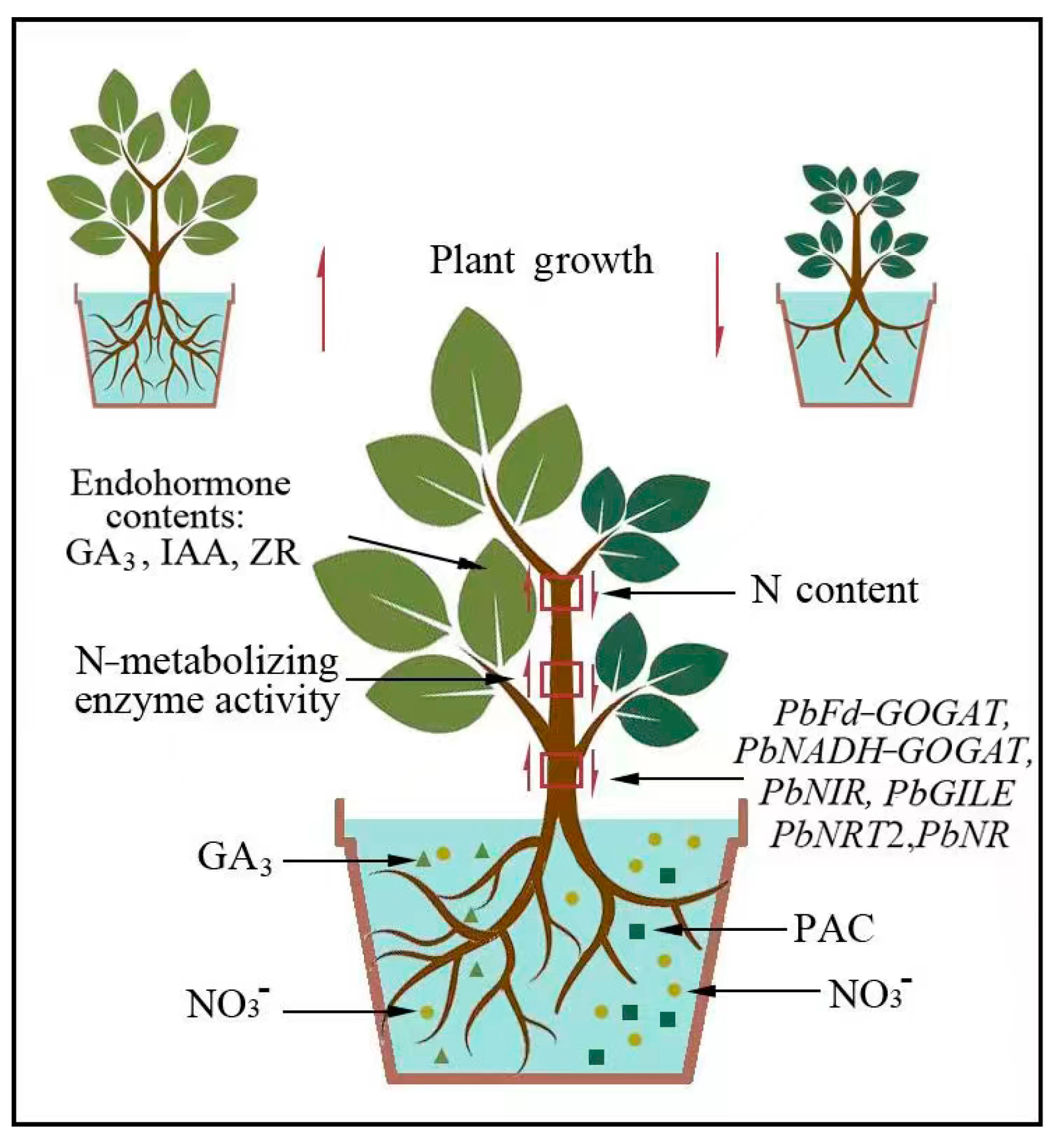
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Analysis of Growth, Chlorophyll Content, and Root System Architecture
4.3. Photosynthetic Parameters and Chlorophyll Fluorescence Determination
4.4. N-Metabolizing Enzyme Activities and Endogenous Hormone Measurements
4.5. Elemental Measurements
4.6. qRT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Tripathi, P.; Chandra, A.; Prakash, J. Physio-biochemical assessment and expression analysis of genes associated with drought tolerance in sugarcane (Saccharum spp. hybrids) exposed to GA3 at grand growth stage. Plant Biol. 2019, 21, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutierrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.L.; Li, M.J.; Zhou, K.; Sun, T.T.; Hu, L.Y.; Li, C.Y.; Ma, F.W. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol. Bioch. 2018, 127, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Du, P.H.; Yin, B.Y.; Zhou, S.S.; Li, Z.Y.; Zhang, X.Y.; Cao, Y.; Han, R.X.; Shi, C.J.; Liang, B.W.; Xu, J.Z. Melatonin and dopamine mediate the regulation of nitrogen uptake and metabolism at low ammonium levels in Malus hupehensis. Plant Physiol. Biochem. 2022, 171, 182–190. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A Review of Methods to Improve Nitrogen Use Efficiency in Agriculture. Sustanainability 2018, 10, 51. [Google Scholar] [CrossRef]
- Zaman, M.; Kurepin, L.V.; Catto, W.; Pharis, R.P. Enhancing crop yield with the use of N-based fertilizers co-applied with plant hormones or growth regulators. J. Sci. Food Agric. 2015, 95, 1777–1785. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.H.; Wu, K.; Ye, Y.F.; Yu, J.P.; Zhang, J.Q.; Liu, Q.; Hu, M.Y.; Li, H.; Tong, Y.P.; et al. Modulating plant growth-metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef]
- Ren, Y.R.; Zhao, Q.; Yang, Y.Y.; Zhang, R.; Wang, X.F.; Zhang, T.E.; You, C.X.; Huo, H.Q.; Hao, Y.J. Interaction of BTB-TAZ protein MdBT2 and DELLA protein MdRGL3a regulates nitrate-mediated plant growth. Plant Physiol. 2021, 186, 750–766. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Li, H.; Lin, J.; Li, X.; Chang, Y. Isolation and characterization of Calcineurin B-like gene (PbCBL1) and its promoter in birch-leaf pear (Pyrus betulifolia Bunge). Genet. Mol. Res. 2015, 14, 16756–16770. [Google Scholar] [CrossRef]
- Zheng, D.C.; Han, X.; An, Y.; Guo, H.W.; Xia, X.L.; Yin, W.L. The nitrate transporter NRT2.1 functions in the ethylene response to nitrate deficiency in Arabidopsis. Plant Cell Environ. 2013, 36, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. The current status of research on gibberellin biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, J.Y.; Cui, L.W.; Fang, J.G. Mining of candidate genes for grape berry cracking using a genome-wide association study. J. Integr. Agric. 2022, 21, 2291–2304. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef]
- Kaviani, B.; Barandan, A.; Tymoszuk, A.; Kulus, D. Optimization of In vitro propagation of pear (Pyrus communis L.) ‘Pyrodwarf® (S)’ rootstock. Agronomy 2023, 13, 268. [Google Scholar] [CrossRef]
- Ye, Y.; Ou, C.Q.; Wang, F.; Zhang, Y.J.; Ma, L.; Li, S.R.; Liu, Y.L.; Jiang, S.L.; Lin, Y.G. Study on green branches cutting propagation of pear dwarfing rootstock ‘Zhong’ai 1′. China Fruits 2023, 12, 24–29. [Google Scholar] [CrossRef]
- Hu, J.; Huang, B.S.; Yin, H.; Qi, K.J.; Jia, Y.Y.; Xie, Z.H.; Gao, Y.; Li, H.X.; Li, Q.H.; Wang, Z.W.; et al. PearMODB: A multiomics database for pear (Pyrus) genomics, genetics and breeding study. Database 2023, 2023, baad050. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; Gruber, B.D.; von Wiren, N. Its time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, J.F.; Liu, R.; Liu, X.; Jiang, J. Gibberellins regulate root growth by antagonizing the jasmonate pathway in tomato plants in response to potassium deficiency. Sci. Hortic. 2023, 309, 111693. [Google Scholar] [CrossRef]
- Khalil, I.A.; Rahman, H.U. Effect of paclobutrazol on growth, chloroplast pigments and sterol biosynthesis of maize (Zea-mays L). Plant Sci. 1995, 105, 15–21. [Google Scholar] [CrossRef]
- Saito, S.; Okamoto, M.; Shinoda, S.; Kushiro, T.; Koshiba, T.; Kamiya, Y.; Hirai, N.; Todoroki, Y.; Sakata, K.; Nambara, E.; et al. A plant growth retardant, uniconazole, is a potent inhibitor of ABA catabolism in Arabidopsis. Biosci. Biotechnol. Biochem. 2006, 70, 1731–1739. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Ahmad, S.; Ahmad, I.; Hussain, I.; Meng, X.P.; Zhang, X.D.; Javed, T.; Ullah, M.; Ding, R.X.; Xu, P.Z.; et al. Paclobutrazol application favors yield improvement of maize under semiarid regions by delaying leaf senescence and regulating photosynthetic capacity and antioxidant system during grain-filling stage. Agronomy 2020, 10, 187. [Google Scholar] [CrossRef]
- Gopi, R.; Jaleel, C.A.; Sairam, R.; Lakshmanan, G.M.A.; Gomathinayagam, M.; Panneerselvam, R. Differential effects of hexaconazole and paclobutrazol on biomass, electrolyte leakage, lipid peroxidation and antioxidant potential of Daucus carota L. Colloids Surf. B Biointerfaces 2007, 60, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Song, X.D.; Ibrahim, M.E.H.; Jamal, Y.; Younas, M.U.; Zhu, G.L.; Zhou, G.S.; Ali, A.Y.A. The role of melatonin in plant growth and metabolism, and its interplay with nitric oxide and auxin in plants under different types of abiotic stress. Front. Plant Sci. 2023, 14, 1108507. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.Q.; Deng, H.H.; Zhang, X.C.; Yu, X.C.; Li, Y.S. Gibberellin is involved in inhibition of cucumber growth and nitrogen uptake at suboptimal root-zone temperatures. PLoS ONE 2016, 11, e0156188. [Google Scholar] [CrossRef] [PubMed]
- Ockerse, R.; Galston, A.W. Gibberellin-auxin interaction in pea stem elongation. Plant Physiol. 1967, 42, 47–54. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; George, G.; Ongaro, V.; Williamson, L.; Willetts, B.; Ljung, K.; McCulloch, H.; Leyser, O. Auxin and strigolactone signaling are required for modulation of Arabidopsis shoot branching by nitrogen supply. Plant Physiol. 2014, 166, 384–395. [Google Scholar] [CrossRef]
- Gho, Y.S.; Song, M.Y.; Bae, D.Y.; Choi, H.; Jung, K.H. Rice PIN auxin efflux carriers modulate the nitrogen response in a changing nitrogen growth environment. Int. J. Mol. Sci. 2021, 22, 3243. [Google Scholar] [CrossRef] [PubMed]
- Otvos, K.; Marconi, M.; Vega, A.; O’Brien, J.; Johnson, A.; Abualia, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.Z.; Tan, S.T.; et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Davenport, S.; Le Lay, P.; Sanchez-Tamburrrino, J.P. Nitrate metabolism in tobacco leaves overexpressing Arabidopsis nitrite reductase. Plant Physiol. Biochem. 2015, 97, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Chanda, S.V.; Sood, C.R.; Reddy, V.S.; Singh, Y.D. Influence of plant growth regulators on some enzymes of nitrogen assimilation in mustard seedlings. J. Plant Nutr. 1998, 21, 1765–1777. [Google Scholar] [CrossRef]
- Qin, C.; Li, B.S.; Wu, W.; Su, Y.Y.; Niu, G.L.; Chen, F.C.; Zhang, C.X.; Wei, C.C.; Yuan, S.; Zhang, L.X. Exogenous application of indole acetic acid (IAA) and giberrelic acid (GA3) induces changes in carbon and nitrogen metabolisms that affect tobacco (Nicotiana tabacum L.) production. Pak. J. Bot. 2019, 51, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Dumlupinar, R.; Genisel, M.; Erdal, S.; Korkut, T.; Taspinar, M.S.; Taskin, M. Effects of progesterone, beta-estradiol, and androsterone on the changes of inorganic element content in barley leaves. Biol. Trace Elem. Res. 2011, 143, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Lin, L.J.; Ma, Q.Q.; Liao, M.A.; Wang, X.; Lai, Y.S.; Liang, D.; Xia, H.; Tang, Y.; Wang, J.; et al. Uniconazole (S-3307) strengthens the growth and cadmium accumulation of accumulator plant Malachium aquaticum. Int. J. Phytoremediat. 2016, 19, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Bai, L.Q.; Sun, M.T.; Wang, J.; Li, S.Z.; Miao, L.; Yan, Y.; He, C.X.; Yu, X.C.; Li, Y.S. Adaptation of cucumber seedlings to low temperature stress by reducing nitrate to ammonium during it’s transportation. BMC Plant Biol. 2021, 21, 189. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Zho, Y.W.; Chen, S.Y.; Liu, J.L.; Fan, K.; Li, Z.W.; Liu, Z.J.; Lin, W.X. Gibberellins play dual roles in response to phosphate starvation of tomato seedlings, negatively in shoots but positively in roots. J. Plant Physiol. 2019, 234, 145–153. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Vieira Pires, A.J.; de Matos Neto, U.; Pinto de Carvalho, G.G.; Veloso, C.M.; da Silva, F.F. Morphogenesis of tanzaniagrass submitted to fertilization and cut intensities. Rev. Bras. Zootec. 2007, 36, 1006–1013. [Google Scholar] [CrossRef]
- Wild, M.; Davière, J.M.; Regnault, T.; Sakvarelidze-Achard, L.; Carrera, E.; Lopez Diaz, I.; Cayrel, A.; Dubeaux, G.; Vert, G.; Achard, P. Tissue-specific regulation of gibberellin signaling fine-tunes Arabidopsis iron-deficiency responses. Dev. Cell 2016, 37, 190–200. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrate transporters in plants: Structure, function and regulation. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Sun, Y.J.; Sun, Y.Y.; Yan, F.J.; Li, Y.X.; Wu, Y.; Guo, C.C.; Ma, P.; Yang, G.T.; Yang, Z.Y.; Ma, J. Coordinating postanthesis carbon and nitrogen metabolism of hybrid rice through different irrigation and nitrogen regimes. Agronomy 2020, 10, 1187. [Google Scholar] [CrossRef]
- Wang, J.; Qin, H.; Zhou, S.R.; Wei, P.C.; Zhang, H.W.; Zhou, Y.; Miao, Y.C.; Huang, R.F. The ubiquitin-binding protein OsDSK2a mediates seedling growth and salt responses by regulating gibberellin metabolism in rice OPEN. Plant Cell 2020, 32, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, M.; Simonovic, A.; Bogdanovic, M.; Subotic, A.; Ghalawenji, N.; Dragicevic, I.; Todorovic, S. Differential regulation of GS-GOGAT gene expression by plant growth regulators in Arabidopsis seedlings. Arch. Biol. Sci. 2016, 68, 399–404. [Google Scholar] [CrossRef]
- Song, P.L.; Li, G.; Xu, J.F.; Ma, Q.C.; Qi, B.X.; Zhang, Y.X. Genome-wide analysis of genes involved in the GA signal transduction pathway in ‘duli’ pear (Pyrus betulifolia Bunge). Int. J. Mol. Sci. 2022, 23, 6570. [Google Scholar] [CrossRef]
- Li, X.G.; Gao, Y.; Li, Y.L.; Yan, S.; Li, W.Z.; Zhang, J.G. Isolation and characterization of PbCS2 gene regulated by iron deficiency and auxin-based systemic signals in Pyrus betulifolia. Sci. Hortic. 2016, 205, 25–31. [Google Scholar] [CrossRef]
- Chen, G.D.; Wang, L.; Fabrice, M.R.; Tian, Y.N.; Qi, K.J.; Chen, Q.; Cao, P.; Wang, P.; Zhang, S.L.; Wu, J.; et al. Physiological and nutritional responses of pear seedlings to nitrate concentrations. Front. Plant Sci. 2018, 9, 1679. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Liang, B.W.; Ma, C.Q.; Zhang, Z.J.; Wei, Z.W.; Gao, T.T.; Zhao, Q.; Ma, F.W.; Li, C. Long-term exogenous application of melatonin improves nutrient uptake fluxes in apple plants under moderate drought stress. Environ. Exp. Bot. 2018, 155, 650–661. [Google Scholar] [CrossRef]
- Li, X.J.; Lu, X.H.; Liu, M.S.; Xiang, C.G.; Liu, W.Q.; Wang, C.C.; Zhang, X.J.; Wang, T.; Liu, Z.X.; Gao, L.H.; et al. Genome-wide characterization of glutamine synthetase family genes in cucurbitaceae and their potential roles in cold response and rootstock-scion signaling communication. Agriculture 2021, 11, 1156. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.J.; Shi, X.X.; Du, G.Q.; Zhang, Y.X. Sufficient assimilate provision to fruits in different positions within an inflorescence: A critical factor determining the morphogenesis and development of Yatu in ‘Yali’ pear (Pyrus bretschneideri Rehd). J. Hortic. Sci. Biotechnol. 2021, 96, 356–363. [Google Scholar] [CrossRef]
- Amir, G.; Hamid, D.; Omidali, A.; Ashkboos, A.; Kaveh, S.; Mehrdad, H.; Aram, S.Z. Assessment of Iranian wheat germplasm for salinity tolerance using analysis of the membership function value of salinity tolerance (MFVS). J. Crop Sci. Biotechnol. 2022, 25, 611–619. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pn (µM CO2 m−2 s−1) | Gs (mol H2O m−2 s−1) | Ci (µM CO2 m−2 s−1) | Tr (mM H2O m−2 s−1) | Fo | Fv/Fm | qP | Rfd |
|---|---|---|---|---|---|---|---|---|
| SCK | 12.02 ± 0.30 e | 0.30 ± 0.02 e | 371.24 ± 13.86 a | 11.61 ± 0.67 e | 120.41 ± 4.83 d | 0.77 ± 0.05 c | 0.25 ± 0.04 b | 1.44 ± 0.03 e |
| SGT | 12.97 ± 0.55 e | 0.28 ± 0.01 e | 341.08 ± 19.95 b | 11.89 ± 0.40 e | 118.05 ± 7.32 d | 0.77 ± 0.04 c | 0.27 ± 0.03 ab | 1.55 ± 0.03 c |
| SPT | 14.80 ± 0.07 d | 0.50 ± 0.02 d | 284.46 ± 9.42 c | 16.48 ± 0.42 d | 159.06 ± 5.30 ab | 0.83 ± 0.02 b | 0.27 ± 0.01 ab | 1.43 ± 0.02 e |
| DCK | 15.78 ± 0.23 c | 0.53 ± 0.04 c | 281.65 ± 9.68 c | 17.17 ± 0.48 bc | 136.29 ± 6.38 c | 0.83 ± 0.01 b | 0.26 ± 0.01 ab | 1.49 ± 0.02 d |
| DGT | 16.91 ± 0.36 a | 0.68 ± 0.01 a | 246.11 ± 21.78 d | 19.21 ± 0.12 a | 139.61 ± 4.69 c | 0.82 ± 0.01 b | 0.29 ± 0.01 a | 1.81 ± 0.01 a |
| DPT | 16.25 ± 0.39 b | 0.62 ± 0.01 b | 258.16 ± 19.51 cd | 16.76 ± 1.32 cd | 166.76 ± 3.26 a | 0.89 ± 0.02 a | 0.27 ± 0.02 ab | 1.50 ± 0.03 d |
| CK | 16.54 ± 0.05 ab | 0.60 ± 0.01 b | 201.99 ± 10.54 e | 17.48 ± 1.77 b | 151.44 ± 5.20 b | 0.86 ± 0.01 a | 0.27 ± 0.01 ab | 1.69 ± 0.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Cheng, X.; Jing, Z.; Cao, Y.; Yuan, S.; Zhang, H.; Zhang, Y. Exogenous GA3 Enhances Nitrogen Uptake and Metabolism under Low Nitrate Conditions in ‘Duli’ (Pyrus betulifolia Bunge) Seedlings. Int. J. Mol. Sci. 2024, 25, 7967. https://doi.org/10.3390/ijms25147967
Zhang W, Cheng X, Jing Z, Cao Y, Yuan S, Zhang H, Zhang Y. Exogenous GA3 Enhances Nitrogen Uptake and Metabolism under Low Nitrate Conditions in ‘Duli’ (Pyrus betulifolia Bunge) Seedlings. International Journal of Molecular Sciences. 2024; 25(14):7967. https://doi.org/10.3390/ijms25147967
Chicago/Turabian StyleZhang, Weilong, Xiaohua Cheng, Zhaotian Jing, Ying Cao, Shuai Yuan, Haixia Zhang, and Yuxing Zhang. 2024. "Exogenous GA3 Enhances Nitrogen Uptake and Metabolism under Low Nitrate Conditions in ‘Duli’ (Pyrus betulifolia Bunge) Seedlings" International Journal of Molecular Sciences 25, no. 14: 7967. https://doi.org/10.3390/ijms25147967