
Mouse Model of Parkinson’s Disease with Bilateral Dorsal Striatum Lesion with 6-Hydroxydopamine Exhibits Cognitive Apathy-like Behavior
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
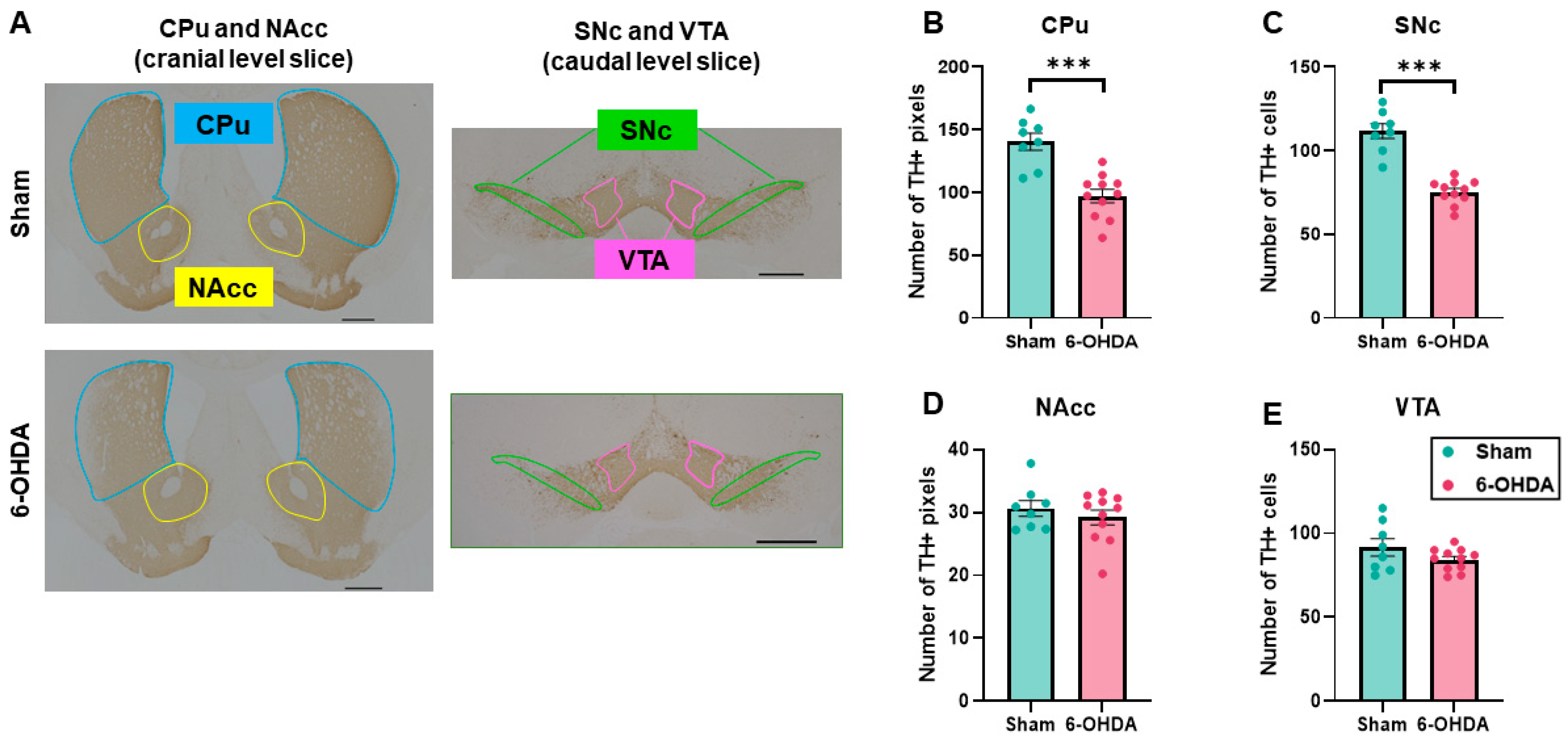
2.1. Decrease in Tyrosine Hydroxylase (TH) Immunostaining in the Caudate Putamen (CPu) and Substantia Nigra Compacta (SNc) by 6-OHDA Injections
2.2. Motor Performance Decline in the 6-OHDA Group
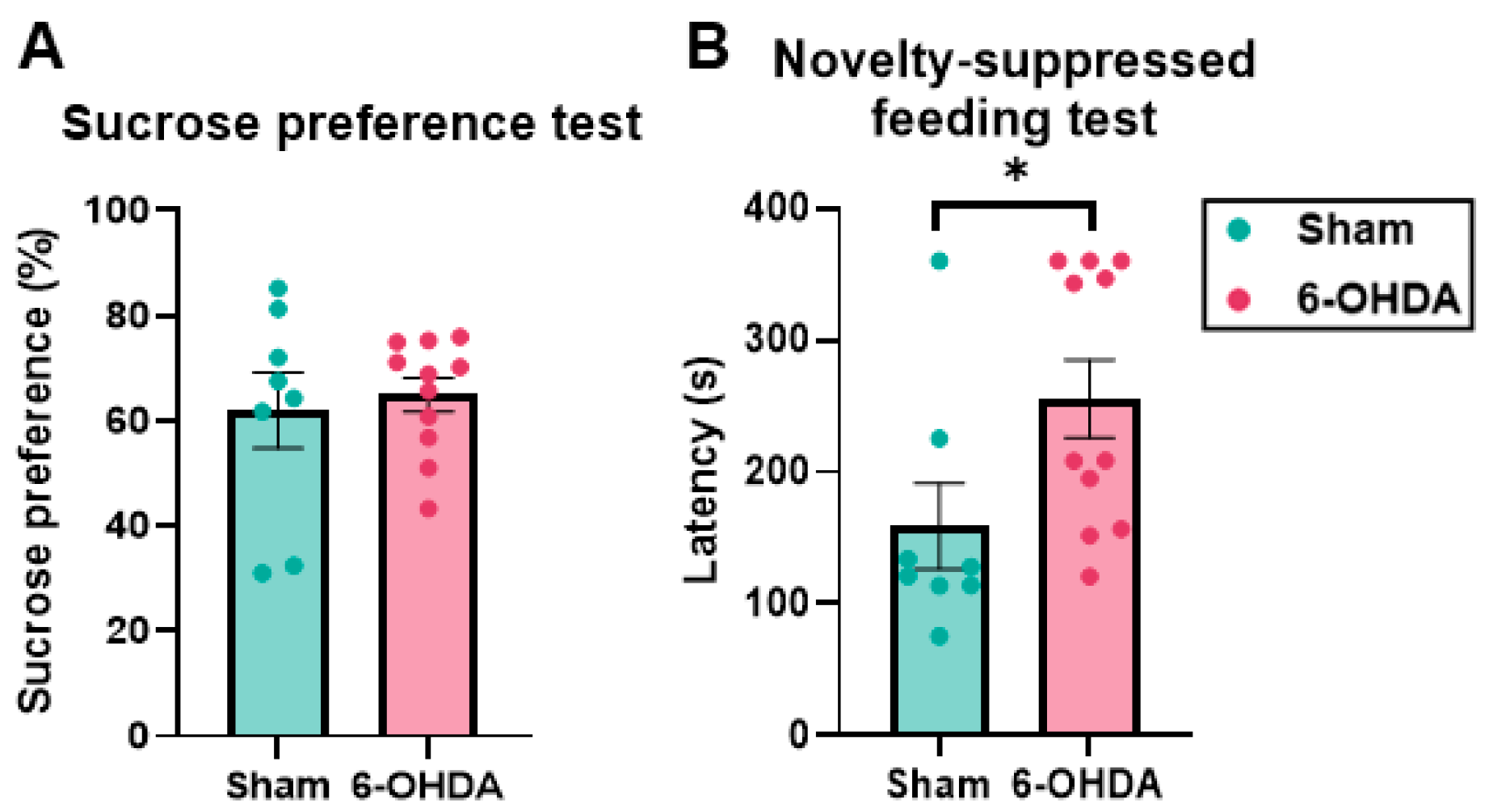
2.3. Anhedonia-like Behavior Suggested by the Novelty-Suppressed Feeding Test (NSFT) but Not by the Sucrose Preference Test (SPT) in the 6-OHDA Group
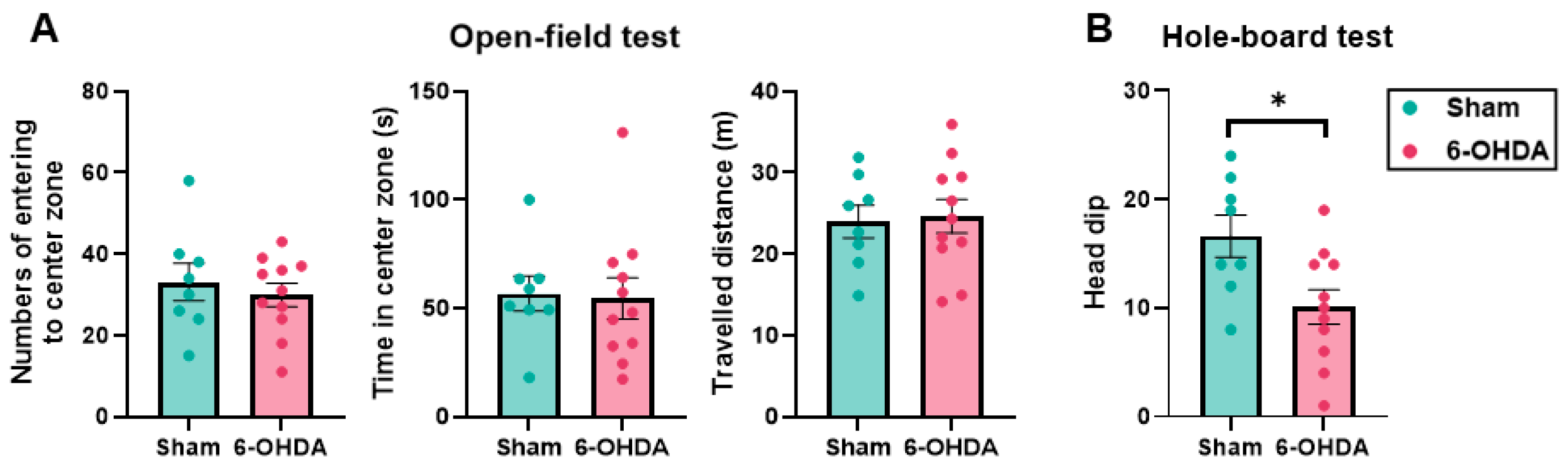
2.4. Anxiety Was Not Observed but Novelty-Seeking Behavior Decreased in the 6-OHDA Group
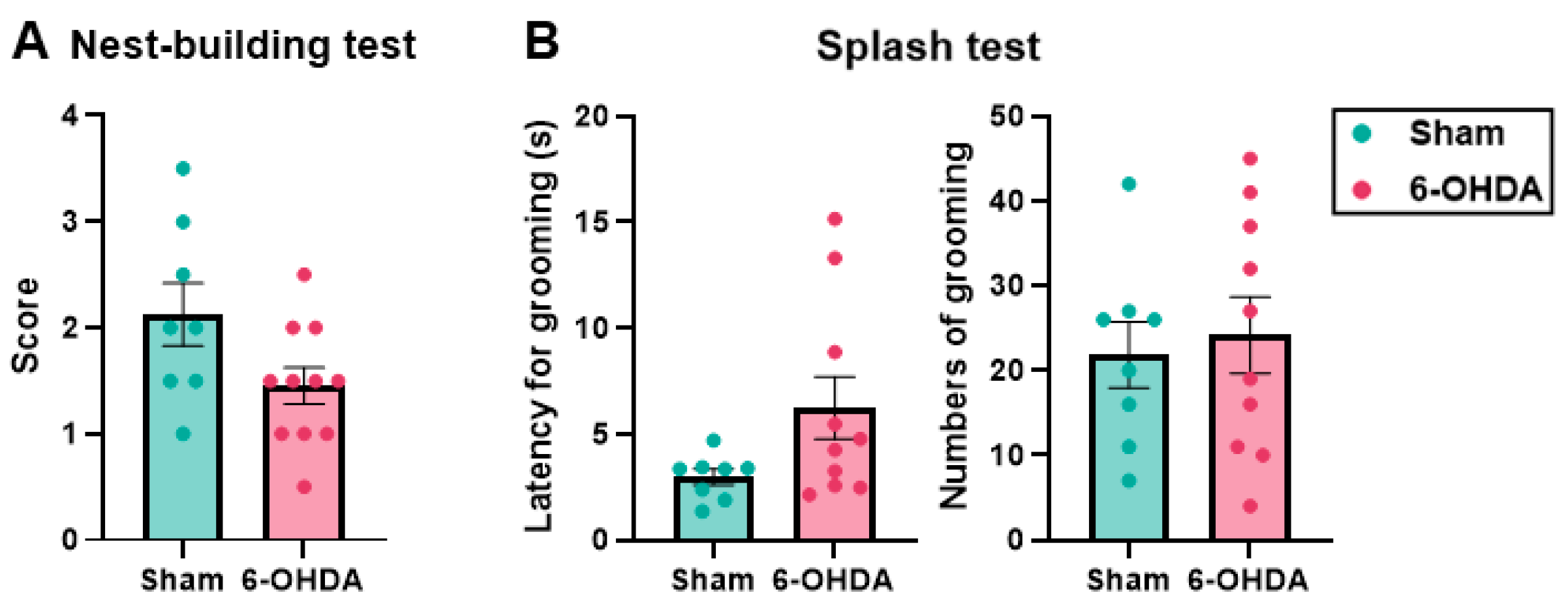
2.5. Self-Care Behavior Was Maintained in the 6-OHDA Group
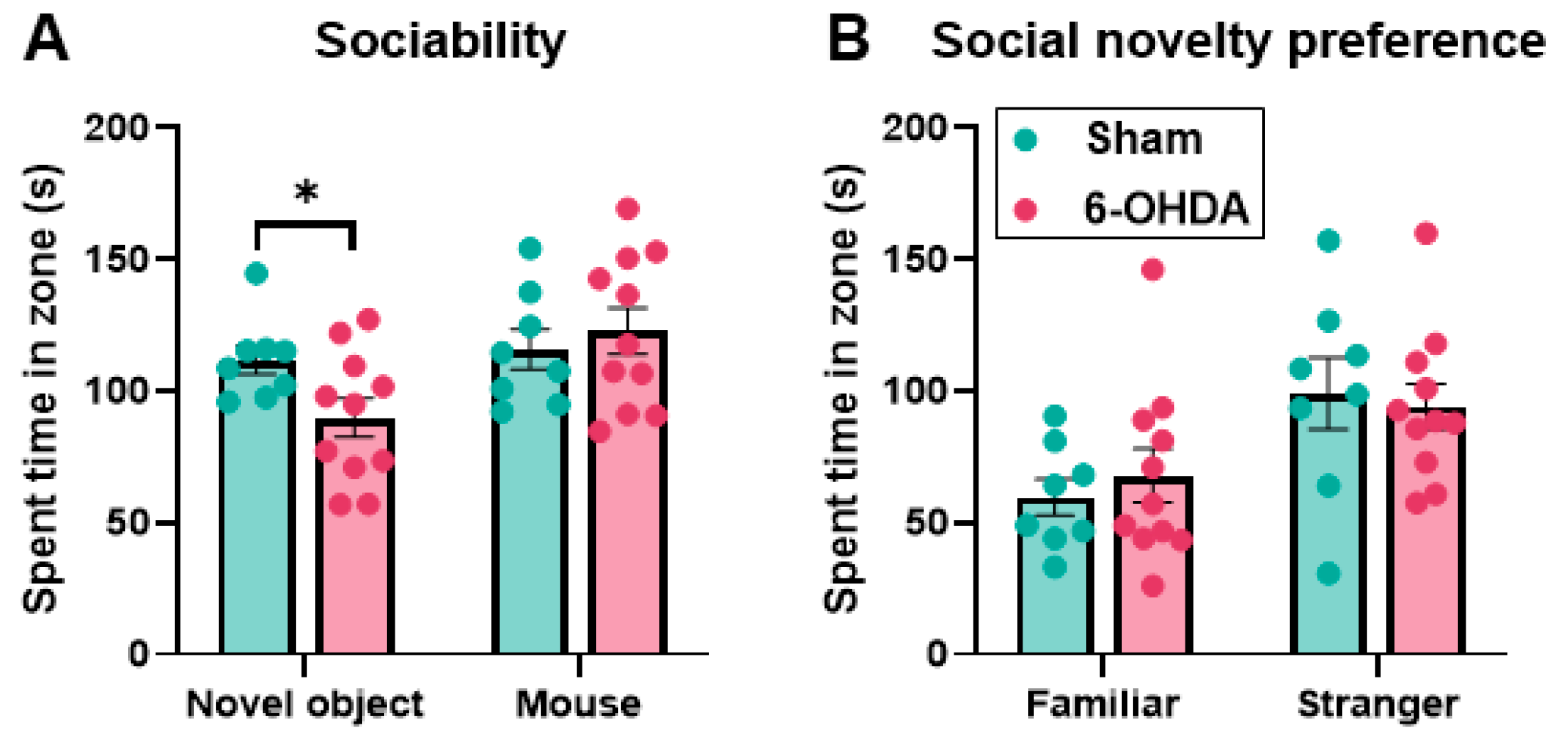
2.6. Object/Inanimate Novelty Was Impaired but Social Novelty Was Maintained in the 6-OHDA Group
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. 6-Hydroxydopamine Lesioning (Day 29)
4.4. Behavioral Assessment (Motor Performance Tests)
4.4.1. Accelerating Rotarod (Days 15 and 16 for the Pretest and Days 50 and 51 for the Posttest)
4.4.2. Pole Test (Days 18 and 19 for the Pretest and Days 53 and 54 for the Posttest)
4.4.3. Balance Beam Test (Days 22–24 for the Pretest and Days 57–59 for the Posttest)
4.5. Behavioral Assessment (Apathy-Related Behavioral Tests)
4.5.1. Sucrose Preference Test (Days 79–81)
4.5.2. Novelty-Suppressed Feeding Test (Days 64 and 65)
4.5.3. Open-Field Test (Day 86)
4.5.4. Hole-Board Test (Day 89)
4.5.5. Nest-Building Test (Day 71)
4.5.6. Splash Test (Day 68)
4.5.7. Three-Chamber TEST (Days 73–75)
4.6. Immunohistochemistry
4.7. Quantitative Analysis of TH-Positive Cells
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef]
- Ray, C.K.; Antonini, A.; Robieson, W.Z.; Sanchez-Soliño, O.; Bergmann, L.; Poewe, W.; GLORIA study co-investigators. Burden of non-motor symptoms in Parkinson’s disease patients predicts improvement in quality of life during treatment with levodopa-carbidopa intestinal gel. Eur. J. Neurol. 2019, 26, 581-e43. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Pagonabarraga, J.; Kulisevsky, J.; Strafella, A.P.; Krack, P. Apathy in Parkinson’s disease: Clinical features, neural substrates, diagnosis, and treatment. Lancet Neurol. 2015, 14, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Ou, R.; Lin, J.; Liu, K.; Jiang, Z.; Wei, Q.; Hou, Y.; Zhang, L.; Cao, B.; Zhao, B.; Song, W.; et al. Evolution of Apathy in Early Parkinson’s Disease: A 4-Years Prospective Cohort Study. Front. Aging Neurosci. 2021, 12, 620762. [Google Scholar] [CrossRef]
- Mele, B.; Van, S.; Holroyd-Leduc, J.; Ismail, Z.; Pringsheim, T.; Goodarzi, Z. Diagnosis, treatment and management of apathy in Parkinson’s disease: A scoping review. BMJ Open 2020, 10, e037632. [Google Scholar] [CrossRef] [PubMed]
- Giannouli, V.; Tsolaki, M. Is Depression or Apathy Playing a Key Role in Predicting Financial Capacity in Parkinson’s Disease with Dementia and Frontotemporal Dementia? Brain Sci. 2021, 11, 785. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Cho, S.H.; Choe, Y.; Kim, B.C. Clinical determinants of apathy and its impact on health-related quality of life in early Parkinson disease. Medicine 2023, 102, e32674. [Google Scholar] [CrossRef] [PubMed]
- LeWitt, P.A.; Chaudhuri, K.R. Unmet needs in Parkinson disease: Motor and non-motor. Park. Relat. Disord. 2020, 80, S7–S12. [Google Scholar] [CrossRef]
- Béreau, M.; Van, W.V.; Servant, M.; Magnin, E.; Tatu, L.; Anheim, M. Apathy in Parkinson’s Disease: Clinical Patterns and Neurobiological Basis. Cells 2023, 12, 1599. [Google Scholar] [CrossRef]
- Levy, R.; Dubois, B. Apathy and the functional anatomy of the prefrontal cortex-basal ganglia circuits. Cereb. Cortex. 2006, 16, 916–928. [Google Scholar] [CrossRef] [PubMed]
- De Waele, S.; Cras, P.; Crosiers, D. Apathy in Parkinson’s Disease: Defining the Park Apathy Subtype. Brain Sci. 2022, 12, 923. [Google Scholar] [CrossRef]
- Del-Monte, J.; Bayard, S.; Graziani, P.; Gély-Nargeot, M.C. Cognitive, Emotional, and Auto-Activation Dimensions of Apathy in Parkinson’s Disease. Front. Behav. Neurosci. 2017, 11, 230. [Google Scholar] [CrossRef] [PubMed]
- Thompson, N.; MacAskill, M.; Pascoe, M.; Anderson, T.; Heron, C.L. Dimensions of apathy in Parkinson’s disease. Brain Behav 2023, 13, e2862. [Google Scholar] [CrossRef] [PubMed]
- Le, H.C.; Apps, M.A.J.; Husain, M. The anatomy of apathy: A neurocognitive framework for amotivated behaviour. Neuropsychologia 2018, 118, 54–67. [Google Scholar] [CrossRef]
- Mah, L.; Arnold, M.C.; Grafman, J. Impairment of social perception associated with lesions of the prefrontal cortex. Am. J. Psychiatry 2004, 161, 1247–1255. [Google Scholar] [CrossRef]
- Dickson, S.S.; Husain, M. Are there distinct dimensions of apathy? The argument for reappraisal. Cortex 2022, 149, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Robert, P.; Onyike, C.U.; Leentjens, A.F.; Dujardin, K.; Aalten, P.; Starkstein, S.; Verhey, F.R.; Yessavage, J.; Clement, J.P.; Drapier, D.; et al. Proposed diagnostic criteria for apathy in Alzheimer’s disease and other neuropsychiatric disorders. Eur. Psychiatry 2009, 24, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Sockeel, P.; Dujardin, K.; Devos, D.; Denève, C.; Destée, A.; Defebvre, L. The Lille apathy rating scale (LARS), a new instrument for detecting and quantifying apathy: Validation in Parkinson’s disease. J. Neurol. Neurosurg Psychiatry 2006, 77, 579–584. [Google Scholar] [CrossRef]
- Ang, Y.S.; Lockwood, P.; Apps, M.A.; Muhammed, K.; Husain, M. Distinct Subtypes of Apathy Revealed by the Apathy Motivation Index. PLoS ONE 2017, 12, e0169938. [Google Scholar] [CrossRef]
- Drui, G.; Carnicella, S.; Carcenac, C.; Favier, M.; Bertrand, A.; Boulet, S.; Savasta, M. Loss of dopaminergic nigrostriatal neurons accounts for the motivational and affective deficits in Parkinson’s disease. Mol. Psychiatry 2014, 19, 358–367. [Google Scholar] [CrossRef]
- Baumann, A.; Moreira, C.G.; Morawska, M.M.; Masneuf, S.; Baumann, C.R.; Noain, D. Preliminary Evidence of Apathetic-Like Behavior in Aged Vesicular Monoamine Transporter 2 Deficient Mice. Front. Hum. Neurosci. 2016, 10, 587. [Google Scholar] [CrossRef]
- Magnard, R.; Vachez, Y.; Carcenac, C.; Krack, P.; David, O.; Savasta, M.; Boulet, S.; Carnicella, S. What can rodent models tell us about apathy and associated neuropsychiatric symptoms in Parkinson’s disease? Transl. Psychiatry 2016, 6, e753. [Google Scholar] [CrossRef]
- Favier, M.; Carcenac, C.; Drui, G.; Vachez, Y.; Boulet, S.; Savasta, M.; Carnicella, S. Implication of dorsostriatal D3 receptors in motivational processes: A potential target for neuropsychiatric symptoms in Parkinson’s disease. Sci. Rep. 2017, 7, 41589. [Google Scholar] [CrossRef]
- Parrella, E.; Del, G.F.; Porrini, V.; Gussago, C.; Benarese, M.; Fabene, P.F.; Pizzi, M. Age-Dependent Neuropsychiatric Symptoms in the NF-kappaB/c-Rel Knockout Mouse Model of Parkinson’s Disease. Front. Behav. Neurosci. 2022, 16, 831664. [Google Scholar] [CrossRef]
- Cathomas, F.; Hartmann, M.N.; Seifritz, E.; Pryce, C.R.; Kaiser, S. The translational study of apathy-an ecological approach. Front. Behav. Neurosci. 2015, 9, 241. [Google Scholar] [CrossRef]
- Stanojlovic, M.; Pallais, Y.J.P.; Vijayakumar, A.; Kotz, C. Early Sociability and Social Memory Impairment in the A53T Mouse Model of Parkinson’s Disease Are Ameliorated by Chemogenetic Modulation of Orexin Neuron Activity. Mol. Neurobiol. 2019, 56, 8435–8450. [Google Scholar] [CrossRef]
- Subramaniam, S.R.; Magen, I.; Bove, N.; Zhu, C.; Lemesre, V.; Dutta, G.; Elias, C.J.; Lester, H.A.; Chesselet, M.F. Chronic nicotine improves cognitive and social impairment in mice overexpressing wild type alpha-synuclein. Neurobiol. Dis. 2018, 117, 170–180. [Google Scholar] [CrossRef]
- Husain, M.; Roiser, J.P. Neuroscience of apathy and anhedonia: A transdiagnostic study. Anhedonia: A transdiagnostic approach. Nat. Rev. Neurosci. 2018, 19, 470–484. [Google Scholar] [CrossRef]
- Yu, Z.; Lin, Y.T.; Chen, J.C. Knockout of NPFFR2 Prevents LPS-Induced Depressive-like Responses in Mice. Int. J. Mol. Sci. 2021, 22, 7611. [Google Scholar] [CrossRef]
- Liu, M.Y.; Yin, C.Y.; Zhu, L.J.; Zhu, X.H.; Xu, C.; Luo, C.X.; Chen, H.; Zhu, D.Y.; Zhou, Q.G. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat. Protoc. 2018, 13, 1686–1698. [Google Scholar] [CrossRef]
- Markov, D.D. Sucrose Preference Test as a Measure of Anhedonic Behavior in a Chronic Unpredictable Mild Stress Model of Depression: Outstanding Issues. Brain Sci. 2022, 12, 1287. [Google Scholar] [CrossRef]
- Scheggi, S.; De Montis, M.G.; Gambarana, C. Making Sense of Rodent Models of Anhedonia. Int. J. Neuropsychopharmacol. 2018, 21, 1049–1065. [Google Scholar] [CrossRef]
- Fogaça, M.V.; Fukumoto, K.; Franklin, T.; Liu, R.J.; Duman, C.H.; Vitolo, O.V.; Duman, R.S. N-Methyl-D-aspartate receptor antagonist d- methadone produces rapid, mTORC1-dependent antidepressant effects. Neuropsychopharmacology 2019, 44, 2230–2238. [Google Scholar] [CrossRef]
- Gilak-Dalasm, M.; Peeri, M.; Azarbayjani, M.A. Swimming exercise decreases depression-like behaviour and inflammatory cytokines in a mouse model of type 2 diabetes. Exp. Physiol. 2021, 106, 1981–1991. [Google Scholar] [CrossRef]
- Darvas, M.; Postupna, N.; Ladiges, W. Mouse modeling for anxiety disorders in older adults. Aging Pathobiol. Ther. 2021, 3, 77–78. [Google Scholar] [CrossRef]
- Nakamoto, C.; Kawamura, M.; Nakatsukasa, E.; Natsume, R.; Takao, K.; Watanabe, M.; Abe, M.; Takeuchi, T.; Sakimura, K. GluD1 knockout mice with a pure C57BL/6N background show impaired fear memory, social interaction, and enhanced depressive-like behavior. PLoS ONE 2020, 15, e0229288. [Google Scholar] [CrossRef]
- Calpe-López, C.; García-Pardo, M.P.; Martínez-Caballero, M.A.; Santos-Ortíz, A.; Aguilar, M.A. Behavioral Traits Associated With Resilience to the Effects of Repeated Social Defeat on Cocaine-Induced Conditioned Place Preference in Mice. Front. Behav. Neurosci. 2020, 13, 278. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, E.; de la Fuente, L.; de Amo, E.; Cubero, I. Environmental Enrichment During Adolescence Acts as a Protective and Therapeutic Tool for Ethanol Binge-Drinking, Anxiety-like, Novelty Seeking and Compulsive-like Behaviors in C57BL/6J Mice during Adulthood. Front. Behav. Neurosci. 2018, 12, 177. [Google Scholar] [CrossRef]
- Yuan, D.; Liu, C.; Wu, J.; Hu, B. Nest-building activity as a reproducible and long-term stroke deficit test in a mouse model of stroke. Brain Behav. 2018, 8, e00993. [Google Scholar] [CrossRef]
- Planchez, B.; Surget, A.; Belzung, C. Animal models of major depression: Drawbacks and challenges. J. Neural Transm. 2019, 126, 1383–1408. [Google Scholar] [CrossRef]
- Bouguiyoud, N.; Roullet, F.; Bronchti, G.; Frasnelli, J.; Al Aïn, S. Anxiety and Depression Assessments in a Mouse Model of Congenital Blindness. Front. Neurosci. 2022, 15, 807434. [Google Scholar] [CrossRef]
- Rein, B.; Ma, K.; Yan, Z. A standardized social preference protocol for measuring social deficits in mouse models of autism. Nat. Protoc. 2020, 15, 3464–3477. [Google Scholar] [CrossRef]
- Slézia, A.; Hegedüs, P.; Rusina, E.; Lengyel, K.; Solari, N.; Kaszas, A.; Balázsfi, D.; Botzanowski, B.; Acerbo, E.; Missey, F.; et al. Behavioral, neural and ultrastructural alterations in a graded-dose 6-OHDA mouse model of early-stage Parkinson’s disease. Sci. Rep. 2023, 13, 19478. [Google Scholar] [CrossRef]
- Mendes-Pinheiro, B.; Soares-Cunha, C.; Marote, A.; Loureiro-Campos, E.; Campos, J.; Barata-Antunes, S.; Monteiro-Fernandes, D.; Santos, D.; Duarte-Silva, S.; Pinto, L.; et al. Unilateral Intrastriatal 6-Hydroxydopamine Lesion in Mice: A Closer Look into Non-Motor Phenotype and Glial Response. Int. J. Mol. Sci. 2021, 22, 11530. [Google Scholar] [CrossRef]
- Tsao, C.H.; Wu, K.Y.; Su, N.C.; Edwards, A.; Huang, G.J. The influence of sex difference on behavior and adult hippocampal neurogenesis in C57BL/6 mice. Sci. Rep. 2023, 13, 17297. [Google Scholar] [CrossRef]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef]
- Bonito-Oliva, A.; Masini, D.; Fisone, G. A mouse model of non-motor symptoms in Parkinson’s disease: Focus on pharmacological interventions targeting affective dysfunctions. Front. Behav. Neurosci. 2014, 8, 290. [Google Scholar] [CrossRef]
- Fearnley, J.M.; Lees, A.J. Ageing and Parkinson’s disease: Substantia nigra regional selectivity. Brain 1991, 114, 2283–2301. [Google Scholar] [CrossRef]
- Menza, M.A.; Golbe, L.I.; Cody, R.A.; Forman, N.E. Dopamine-related personality traits in Parkinson’s disease. Neurology 1993, 43, 50–58. [Google Scholar] [CrossRef]
- Tapper, A.R.; Molas, S. Midbrain circuits of novelty processing. Neurobiol. Learn Mem. 2020, 176, 107323. [Google Scholar] [CrossRef]
- Schiemann, J.; Schlaudraff, F.; Klose, V.; Bingmer, M.; Seino, S.; Magill, P.J.; Zaghloul, K.A.; Schneider, G.; Liss, B.; Roeper, J. K-ATP channels in dopamine substantia nigra neurons control bursting and novelty-induced exploration. Nat. Neurosci. 2012, 15, 1272–1280. [Google Scholar] [CrossRef]
- Shan, Q.; Tian, Y.; Chen, H.; Lin, X.; Tian, Y. Reduction in the activity of VTA/SNc dopaminergic neurons underlies aging-related decline in novelty seeking. Commun. Biol. 2023, 6, 1224. [Google Scholar] [CrossRef]
- Santangelo, G.; Garramone, F.; Baiano, C.; D’Iorio, A.; Piscopo, F.; Raimo, S.; Vitale, C. Personality and Parkinson’s disease: A meta-analysis. Park. Relat. Disord 2018, 49, 67–74. [Google Scholar] [CrossRef]
- Ponsi, G.; Scattolin, M.; Villa, R.; Aglioti, S.M. Human moral decision-making through the lens of Parkinson’s disease. NPJ Park. Dis. 2021, 7, 18. [Google Scholar] [CrossRef]
- Bódi, N.; Kéri, S.; Nagy, H.; Moustafa, A.; Myers, C.E.; Daw, N.; Dibó, G.; Takáts, A.; Bereczki, D.; Gluck, M.A. Reward-learning and the novelty-seeking personality: A between- and within-subjects study of the effects of dopamine agonists on young Parkinson’s patients. Brain 2009, 132, 2385–2395. [Google Scholar] [CrossRef]
- Dulawa, S.C.; Grandy, D.K.; Low, M.J.; Paulus, M.P.; Geyer, M.A. Dopamine D4 receptor-knock-out mice exhibit reduced exploration of novel stimuli. J. Neurosci. 1999, 19, 9550–9556. [Google Scholar] [CrossRef]
- Nuber, S.; Harmuth, F.; Kohl, Z.; Adame, A.; Trejo, M.; Schönig, K.; Zimmermann, F.; Bauer, C.; Casadei, N.; Giel, C.; et al. A progressive dopaminergic phenotype associated with neurotoxic conversion of α-synuclein in BAC-transgenic rats. Brain 2013, 136, 412–432. [Google Scholar] [CrossRef]
- He, L.W.; Zeng, L.; Tian, N.; Li, Y.; He, T.; Tan, D.M.; Zhang, Q.; Tan, Y. Optimization of food deprivation and sucrose preference test in SD rat model undergoing chronic unpredictable mild stress. Animal Model Exp. Med. 2020, 3, 69–78. [Google Scholar] [CrossRef]
- Beck, R.C.; Self, J.L.; Carter, D.J. Sucrose preference thresholds for satiated and water-deprived rats. Psychol. Rep. 1965, 16, 901–905. [Google Scholar] [CrossRef]
- Cohen, J.S.; Hachey, G.J. Development of sucrose preferences under two levels of water deprivation. Psychol. Rep. 1980, 46, 820–822. [Google Scholar] [CrossRef]
- Stenfeld, K.A.; Clinton, S.M.; Kerman, I.A.; Akil, H.; Watson, S.J.; Sved, A.F. Novelty-seeking behavior predicts vulnerability in a rodent model of depression. Physiol. Behav. 2011, 10, 210–216. [Google Scholar] [CrossRef]
- Gunaydin, L.A.; Grosenick, L.; Finkelstein, J.C.; Kauvar, I.V.; Fenno, L.E.; Adhikari, A.; Lammel, S.; Mirzabekov, J.J.; Airan, R.D.; Zalocusky, K.A.; et al. Natural neural projection dynamics underlying social behavior. Cell 2014, 157, 1535–1551. [Google Scholar] [CrossRef]
- Mortby, M.E.; Adler, L.; Agüera-Ortiz, L.; Bateman, D.R.; Brodaty, H.; Cantillon, M.; Geda, Y.E.; Ismail, Z.; Lanctôt, K.L.; Marshall, G.A.; et al. Apathy as a Treatment Target in Alzheimer’s Disease: Implications for Clinical Trials. Am. J. Geriatr. Psychiatry 2022, 30, 119–147. [Google Scholar] [CrossRef]
- Pasquini, J.; Brooks, D.J.; Pavese, N. The Cholinergic Brain in Parkinson’s Disease. Mov. Disord. Clin. Pract 2021, 8, 1012–1026. [Google Scholar] [CrossRef]
- Anglade, P.; Tsuji, S.; Javoy-Agid, F.; Agid, Y.; Hirsch, E.C. Plasticity of nerve afferents to nigrostriatal neurons in Parkinson’s disease. Ann. Neurol. 1995, 37, 265–272. [Google Scholar] [CrossRef]
- Sanchez-Catasus, C.A.; Bohnen, N.I.; D’Cruz, N.; Muller, M.L.T.M. Striatal acetylcholine-dopamine imbalance in Parkinson’s disease: In vivo neuroimaging study with dual-tracer PET and dopaminergic PET-informed correlational tractography. J. Nucl. Med. 2021, 63, 438–445. [Google Scholar] [CrossRef]
- Polito, C.; Berti, V.; Ramat, S.; Vanzi, E.; De Cristofaro, M.T.; Pellicanò, G.; Mungai, F.; Marini, P.; Formiconi, A.R.; Sorbi, S.; et al. Interaction of caudate dopamine depletion and brain metabolic changes with cognitive dysfunction in early Parkinson’s disease. Neurobiol. Aging 2012, 33, 206.e29–206.e39. [Google Scholar] [CrossRef]
- Cools, R. Dopaminergic control of the striatum for high-level cognition. Curr. Opin. Neurobiol. 2011, 21, 402–407. [Google Scholar] [CrossRef]
- Narayanan, N.S.; Rodnitzky, R.L.; Uc, E.Y. Prefrontal dopamine signaling and cognitive symptoms of Parkinson’s disease. Rev. Neurosci. 2013, 24, 267–278. [Google Scholar] [CrossRef]
- Liu, X.; Yu, H.; Chen, B.; Friedman, V.; Mu, L.; Kelly, T.J.; Ruiz-Pérez, G.; Zhao, L.; Bai, X.; Hillard, C.J.; et al. CB2 Agonist GW842166x Protected against 6-OHDA-Induced Anxiogenic- and Depressive-Related Behaviors in Mice. Biomedicines 2022, 10, 1776. [Google Scholar] [CrossRef]
- Delaville, C.; Deurwaerdère, P.D.; Benazzouz, A. Noradrenaline and Parkinson’s disease. Front. Syst. Neurosci. 2011, 5, 31. [Google Scholar] [CrossRef]
- Thiele, S.L.; Warre, R.; Nash, J.E. Development of a unilaterally-lesioned 6-OHDA mouse model of Parkinson’s disease. J. Vis. Exp. 2012, 60, 3234. [Google Scholar] [CrossRef]
- Campos, F.L.; Carvalho, M.M.; Cristovão, A.C.; Je, G.; Baltazar, G.; Salgado, A.J.; Kim, Y.S.; Sousa, N. Rodent models of Parkinson’s disease: Beyond the motor symptomatology. Front. Behav. Neurosci. 2013, 7, 175. [Google Scholar] [CrossRef]
- Jacquez, B.; Choi, H.; Bird, C.W.; Linsenbardt, D.N.; Valenzuela, C.F. Characterization of motor function in mice developmentally exposed to ethanol using the Catwalk system: Comparison with the triple horizontal bar and rotarod tests. Behav. Brain Res. 2021, 396, 112885. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Chen, B.; Vickstrom, C.R.; Friedman, V.; Kelly, T.J.; Bai, X.; Zhao, L.; Hillard, C.J.; Liu, Q.S. The Neuroprotective Effects of the CB2 Agonist GW842166x in the 6-OHDA Mouse Model of Parkinson’s Disease. Cells 2021, 10, 3548. [Google Scholar] [CrossRef]
- Yoshizaki, K.; Asai, M.; Hara, T. High-Fat Diet Enhances Working Memory in the Y-Maze Test in Male C57BL/6J Mice with Less Anxiety in the Elevated Plus Maze Test. Nutrients 2020, 12, 2036. [Google Scholar] [CrossRef]
- Cao, H.; Zuo, C.; Huang, Y.; Zhu, L.; Zhao, J.; Yang, Y.; Jiang, Y.; Wang, F. Hippocampal proteomic analysis reveals activation of necroptosis and ferroptosis in a mouse model of chronic unpredictable mild stress-induced depression. Behav. Brain Res. 2021, 407, 113261. [Google Scholar] [CrossRef]
- Dieterich, A.; Srivastava, P.; Sharif, A.; Stech, K.; Floeder, J.; Yohn, S.E.; Samuels, B.A. Chronic corticosterone administration induces negative valence and impairs positive valence behaviors in mice. Transl. Psychiatry 2019, 9, 337. [Google Scholar] [CrossRef]
- Frausto-González, O.; Bautista, C.J.; Narváez-González, F.; Hernandez-Leon, A.; Estrada-Camarena, E.; Rivero-Cruz, F.; González-Trujano, M.E. Bertholletia excelsa Seeds Reduce Anxiety-Like Behavior, Lipids, and Overweight in Mice. Molecules 2021, 26, 3212. [Google Scholar] [CrossRef]
- Gaskill, B.N.; Karas, A.Z.; Garner, J.P.; Pritchett-Corning, K.R. Nest building as an indicator of health and welfare in laboratory mice. J. Vis. Exp. 2013, 82, 51012. [Google Scholar] [CrossRef]
- Fujita, M.; Hagino, Y.; Takamatsu, Y.; Shimizu, Y.; Takamatsu, Y.; Ikeda, K.; Hashimoto, M. Early manifestation of depressive-like behavior in transgenic mice that express dementia with Lewy body-linked mutant beta-synuclein. Neuropsychopharmacol. Rep. 2018, 38, 95–97. [Google Scholar] [CrossRef]
- Hess, S.E.; Rohr, S.; Dufour, B.D.; Gaskill, B.N.; Pajor, E.A.; Garner, J.P. Home improvement: C57BL/6J mice given more naturalistic nesting materials build better nests. J. Am. Assoc. Lab. Anim. Sci. 2008, 47, 25–31. [Google Scholar]
- Ho, E.V.; Welch, A.; Thompson, S.L.; Knowles, J.A.; Dulawa, S.C. Mice lacking Ptprd exhibit deficits in goal-directed behavior and female-specific impairments in sensorimotor gating. PLoS ONE 2023, 18, e0277446. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Age-related behavioral changes from young to old age in male mice of a C57BL/6J strain maintained under a genetic stability program. Neuropsychopharmacol. Rep. 2019, 39, 100–118. [Google Scholar] [CrossRef]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: New York, NY, USA, 2008; pp. 23–56. ISBN 978–0123742445. [Google Scholar]
- Ochiai, Y.; Fujita, M.; Hagino, Y.; Kobayashi, K.; Okiyama, R.; Takahashi, K.; Ikeda, K. Therapeutic Effects of Quetiapine and 5-HT1A Receptor Agonism on Hyperactivity in Dopamine-Deficient Mice. Int. J. Mol. Sci. 2022, 23, 7436. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okitsu, M.; Fujita, M.; Moriya, Y.; Kotajima-Murakami, H.; Ide, S.; Kojima, R.; Sekiyama, K.; Takahashi, K.; Ikeda, K. Mouse Model of Parkinson’s Disease with Bilateral Dorsal Striatum Lesion with 6-Hydroxydopamine Exhibits Cognitive Apathy-like Behavior. Int. J. Mol. Sci. 2024, 25, 7993. https://doi.org/10.3390/ijms25147993
Okitsu M, Fujita M, Moriya Y, Kotajima-Murakami H, Ide S, Kojima R, Sekiyama K, Takahashi K, Ikeda K. Mouse Model of Parkinson’s Disease with Bilateral Dorsal Striatum Lesion with 6-Hydroxydopamine Exhibits Cognitive Apathy-like Behavior. International Journal of Molecular Sciences. 2024; 25(14):7993. https://doi.org/10.3390/ijms25147993
Chicago/Turabian StyleOkitsu, Masato, Masayo Fujita, Yuki Moriya, Hiroko Kotajima-Murakami, Soichiro Ide, Rika Kojima, Kazunari Sekiyama, Kazushi Takahashi, and Kazutaka Ikeda. 2024. "Mouse Model of Parkinson’s Disease with Bilateral Dorsal Striatum Lesion with 6-Hydroxydopamine Exhibits Cognitive Apathy-like Behavior" International Journal of Molecular Sciences 25, no. 14: 7993. https://doi.org/10.3390/ijms25147993