
Elevated LAMTOR4 Expression Is Associated with Lethal Prostate Cancer and Its Knockdown Decreases Cell Proliferation, Invasion, and Migration In Vitro

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
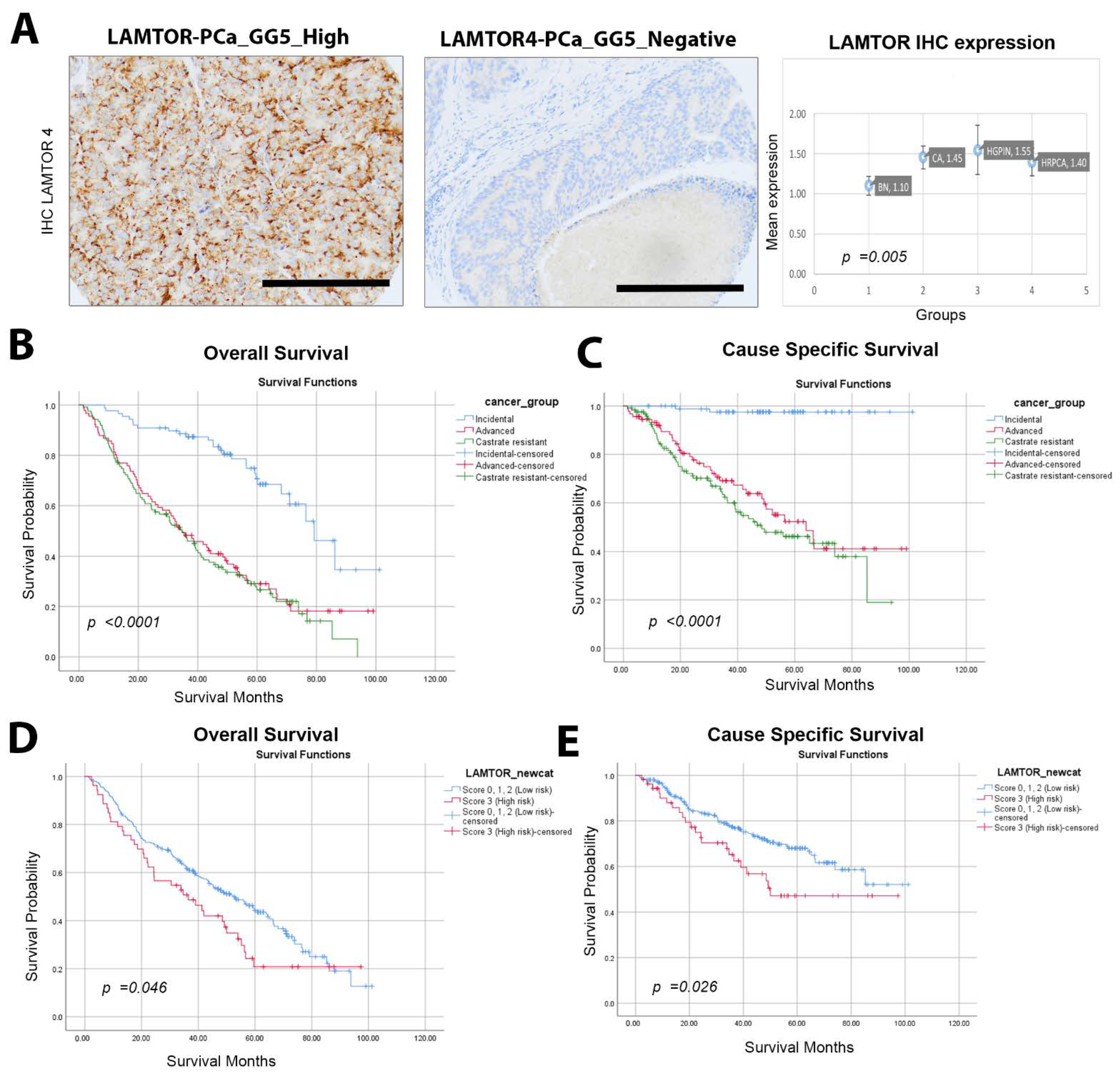
2.1. High LAMTOR4 Expression Correlates with Poor Overall Survival (OS) and Cancer Specific Survival (CSS)
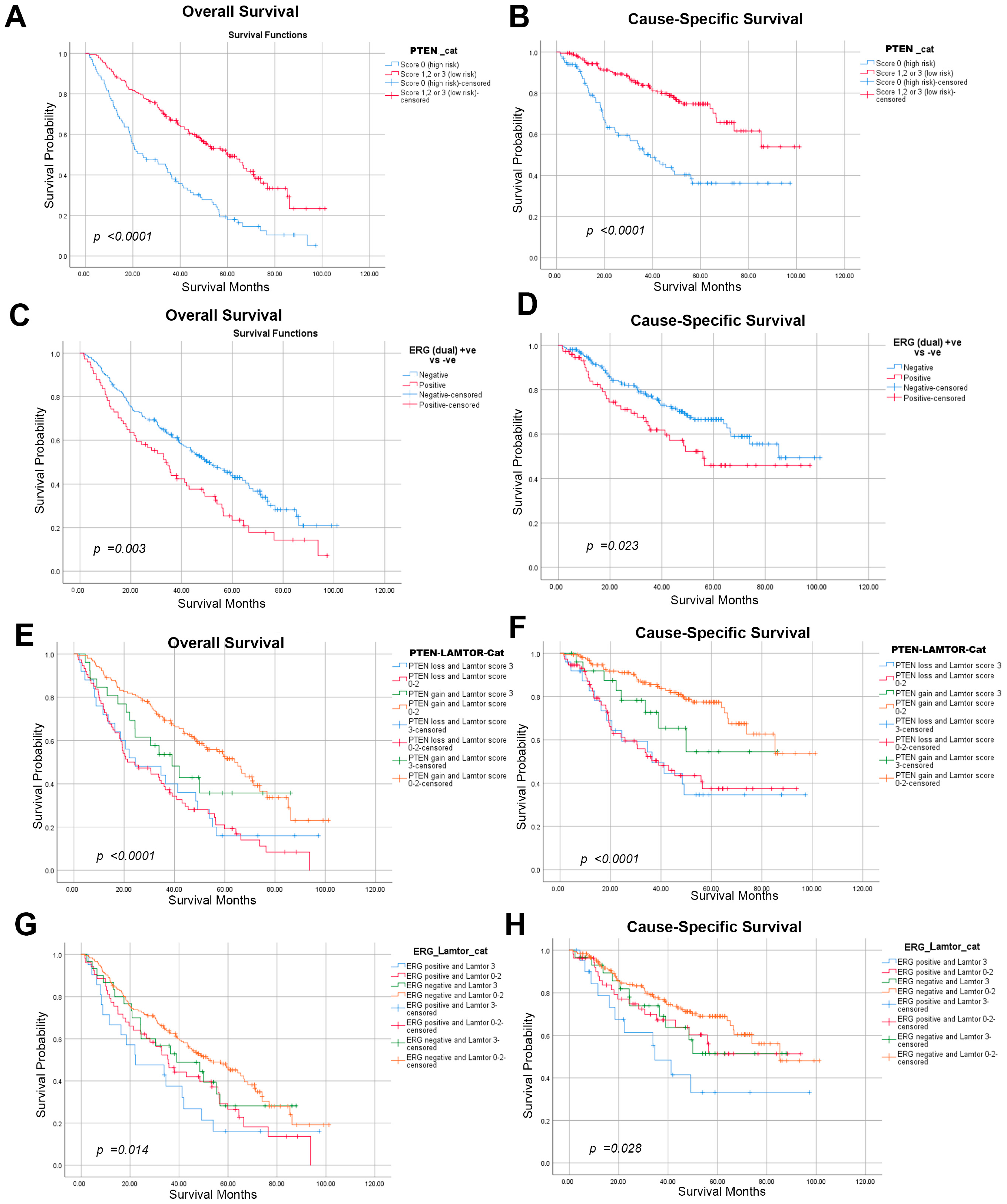
2.2. The Association between High LAMTOR4 Expression and Prognostic Markers PTEN or ERG
2.3. PanCancer Analysis Revealed High Expression of LAMTOR4 mRNA in Most Types of Cancer
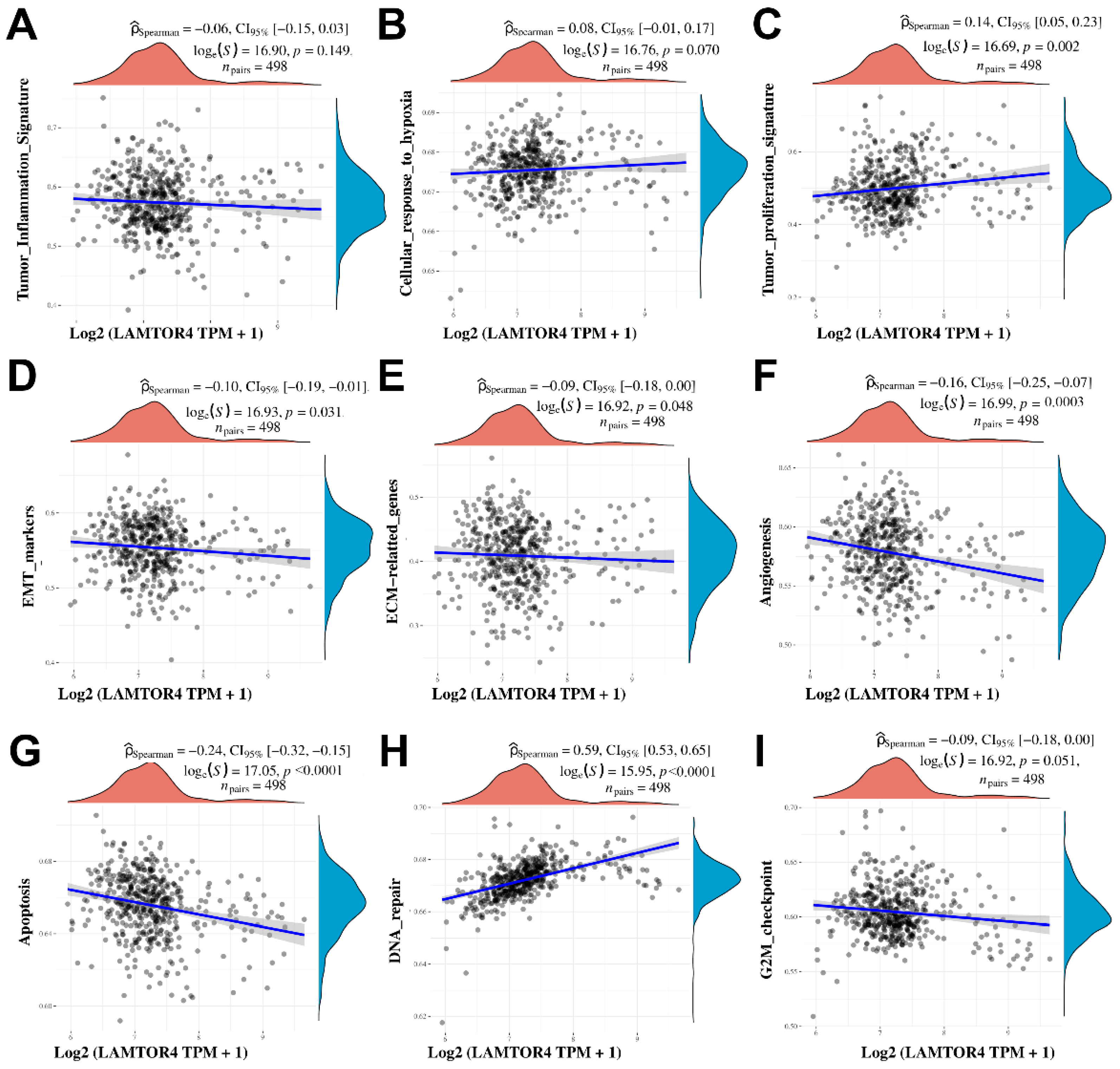
2.4. Differential Gene Expression and Gene Signatures Associated with LAMTOR4 Overexpression
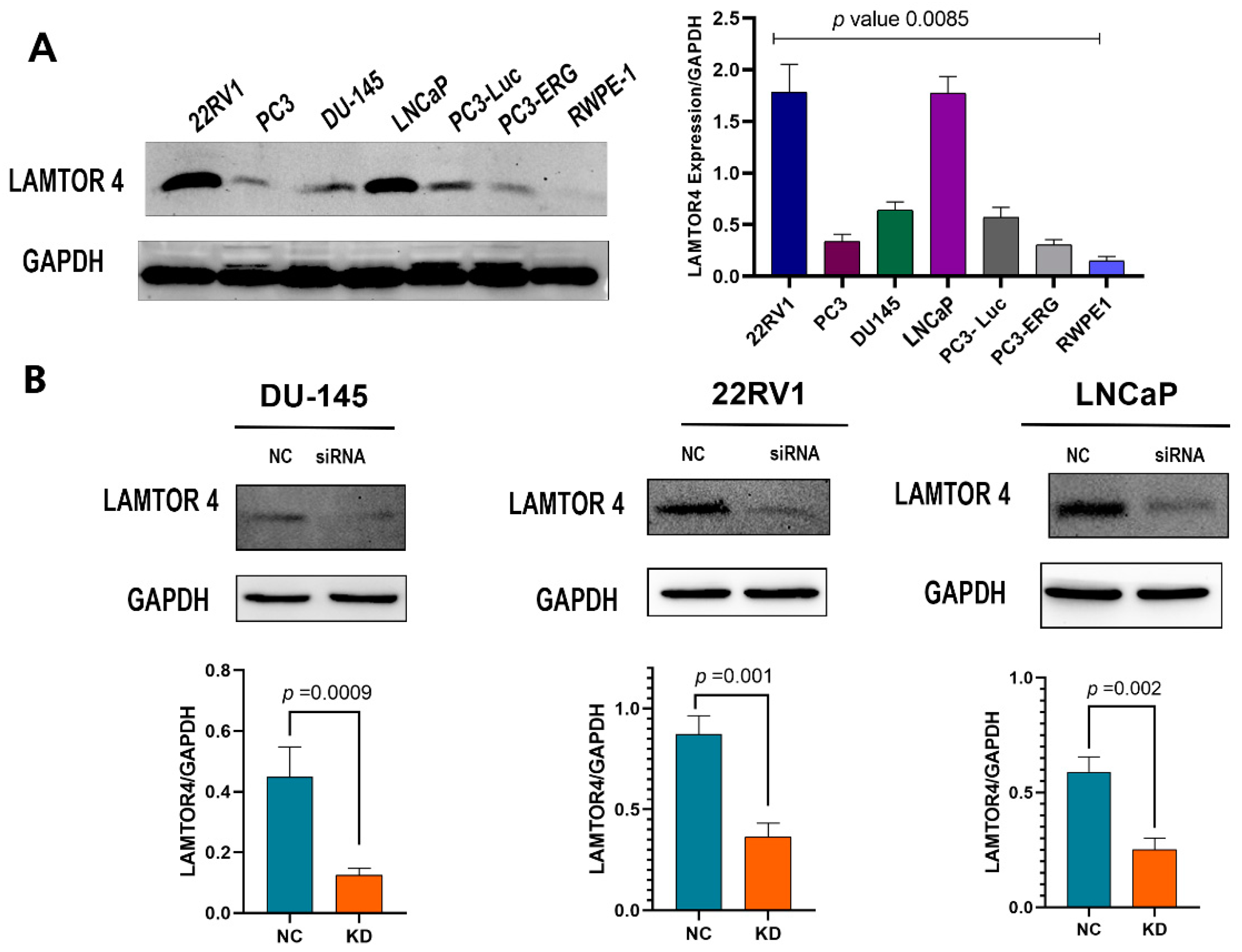
2.5. LAMTOR4 Expression in Human PCa Cell Lines
2.6. LAMTOR4 Knockdown Decreases Proliferation of PCa Cells
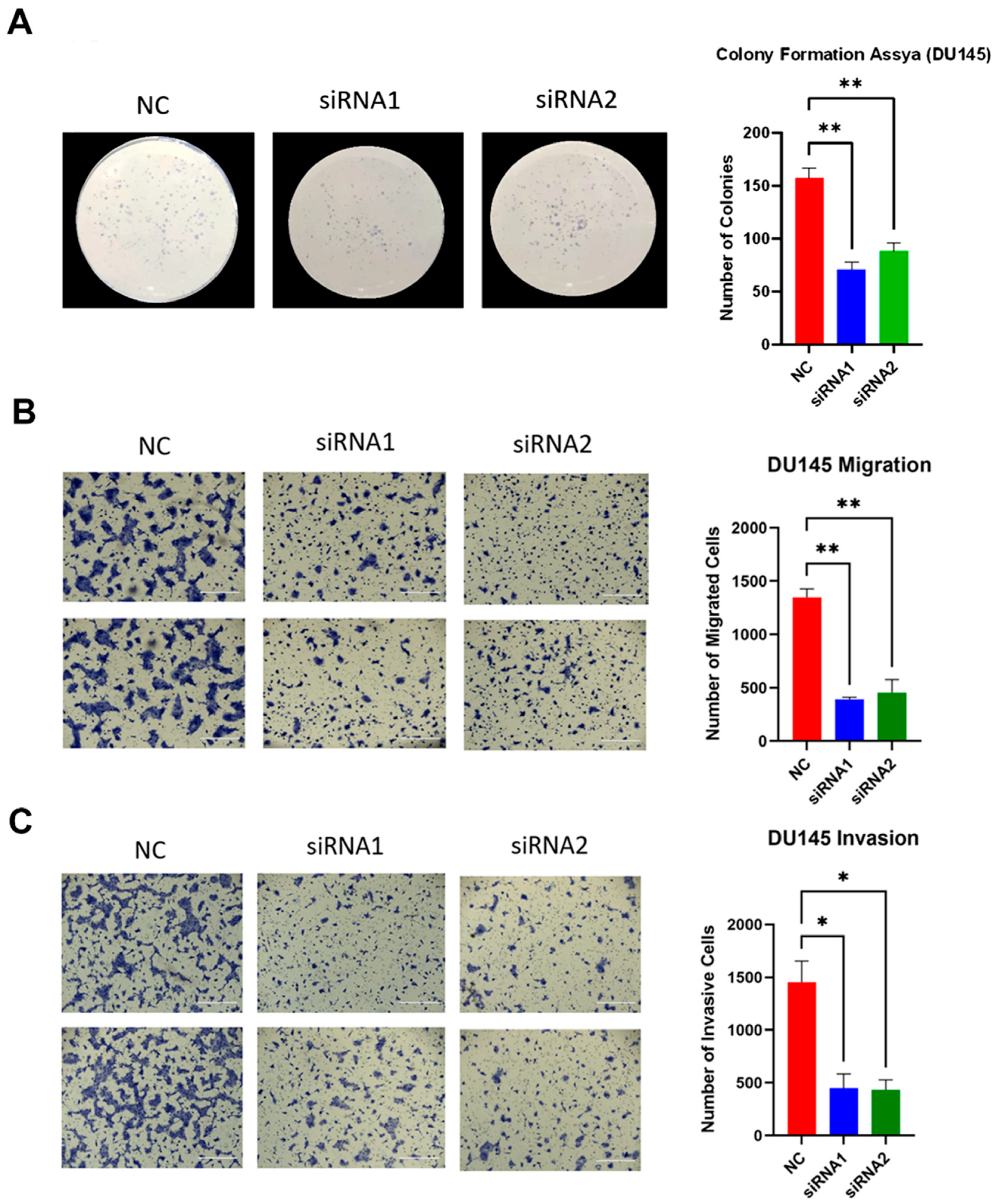
2.7. LAMTOR4 Knockdown Reduces PCa Cell Migration and Invasion of DU-145 Cells
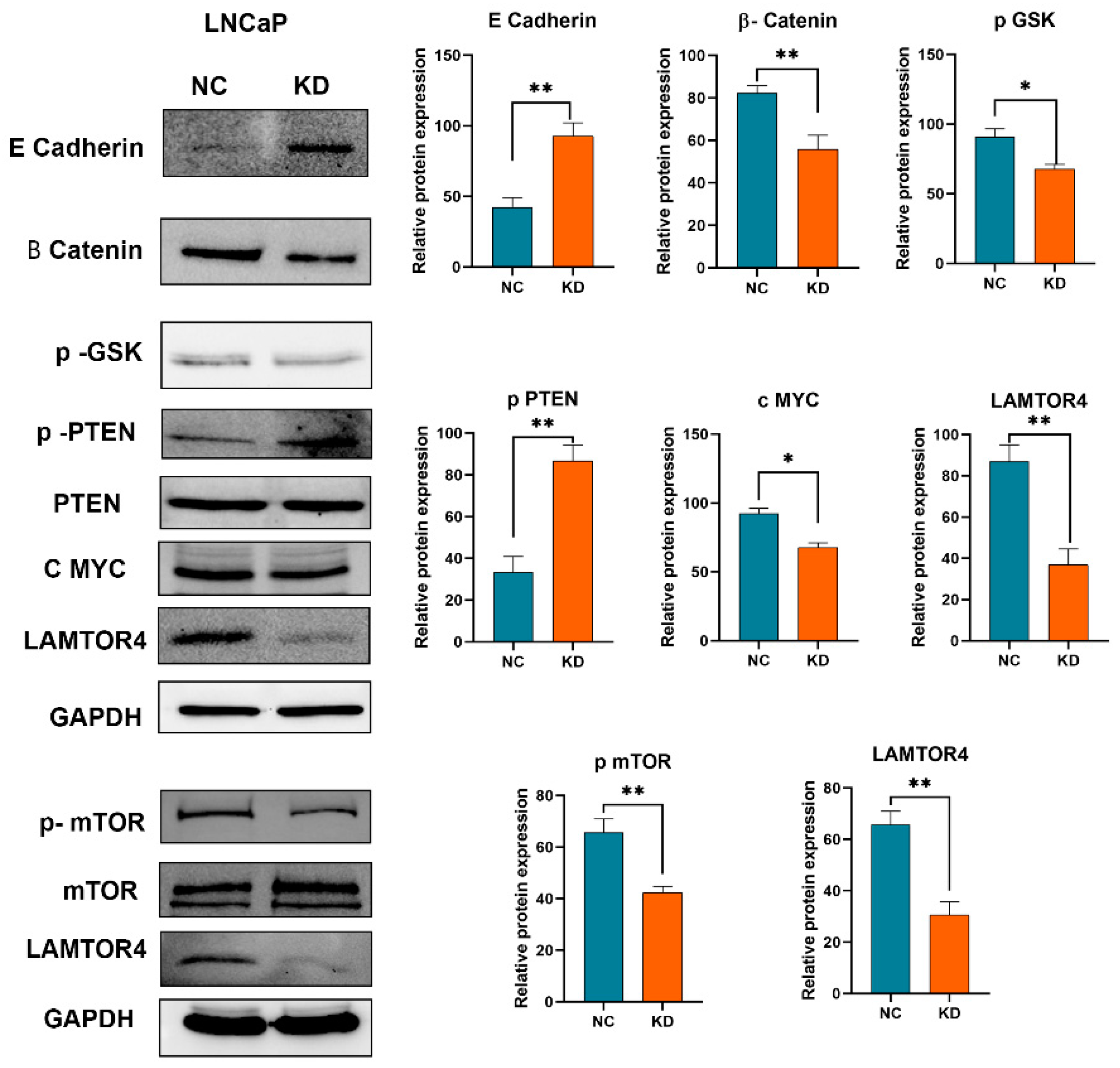
2.8. LAMTOR4 Knockdown Attenuates Potential Pathways in Tumorigenesis
3. Discussion
4. Materials and Methods
4.1. Study Population and Tissue Microarray Construction
4.2. Immunohistochemistry
4.3. Pathological Analysis
4.4. Cell Lines and Cell Culture
4.5. Cell Line Transfection and RNA Silencing
4.6. Western Blot
4.7. Colony Formation Assay
4.8. Migration and Invasion Assay
4.9. LAMTOR4 Expression in the Cancer Genome Atlas TCGA-PRAD
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| LAMTOR4 | Late endosomal/lysosomal adaptor, MAPK, and mTOR activator 4 |
| mTOR | Mammalian target of rapamycin |
| IHC | Immunohistochemistry |
| WB | Western blotting |
| GSEA | Gene Set Enrichment Analysis |
| TCGA | The Cancer Genome Atlas |
| PRAD | Prostate Adenocarcinoma |
| PTEN | Phosphatase and tensin homolog |
| ERG | ETS Transcription Factor ERG |
| CSS | Cause-specific survival |
| OS | Overall survival |
| CRPC | Castration-resistant prostate cancer |
References
- Berenguer, C.V.; Pereira, F.; Camara, J.S.; Pereira, J.A.M. Underlying Features of Prostate Cancer-Statistics, Risk Factors, and Emerging Methods for Its Diagnosis. Curr. Oncol. 2023, 30, 2300–2321. [Google Scholar] [CrossRef] [PubMed]
- Grubb, R.L., 3rd. Prostate Cancer: Update on Early Detection and New Biomarkers. Mo. Med. 2018, 115, 132–134. [Google Scholar] [PubMed]
- Unterberger, C.J.; McIlwain, S.J.; Tsourkas, P.K.; Maklakova, V.I.; Prince, J.L.; Onesti, A.; Hu, R.; Kopchick, J.J.; Swanson, S.M.; Marker, P.C. Conditional gene regulation models demonstrate a pro-proliferative role for growth hormone receptor in prostate cancer. Prostate 2023, 83, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Nonomura, N. Role of Androgen Receptor in Prostate Cancer: A Review. World J. Mens. Health 2019, 37, 288–295. [Google Scholar] [CrossRef]
- Audet-Walsh, E.; Dufour, C.R.; Yee, T.; Zouanat, F.Z.; Yan, M.; Kalloghlian, G.; Vernier, M.; Caron, M.; Bourque, G.; Scarlata, E.; et al. Nuclear mTOR acts as a transcriptional integrator of the androgen signaling pathway in prostate cancer. Genes Dev. 2017, 31, 1228–1242. [Google Scholar] [CrossRef]
- Jacob, A.; Raj, R.; Allison, D.B.; Myint, Z.W. Androgen Receptor Signaling in Prostate Cancer and Therapeutic Strategies. Cancers 2021, 13, 5417. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Li, D.; Yin, J.; Pan, H.; Ye, H.; Bowman, J.; Capaldo, B.; Kelly, K. TMPRSS2-ERG promotes the initiation of prostate cancer by suppressing oncogene-induced senescence. Cancer Gene Ther. 2022, 29, 1463–1476. [Google Scholar] [CrossRef] [PubMed]
- Khosh Kish, E.; Choudhry, M.; Gamallat, Y.; Buharideen, S.M.; Bismar, T.A. The expression of proto-oncogene ETS-related gene (ERG) plays a central role in the oncogenic mechanism involved in the development and progression of prostate cancer. Int. J. Mol. Sci. 2022, 23, 4772. [Google Scholar] [CrossRef] [PubMed]
- Lorenzin, F.; Demichelis, F. Past, current, and future strategies to target ERG fusion-positive prostate cancer. Cancers 2022, 14, 1118. [Google Scholar] [CrossRef]
- Colaço, A.; Jäättelä, M. Ragulator—A multifaceted regulator of lysosomal signaling and trafficking. J. Cell Biol. 2017, 216, 3895. [Google Scholar] [CrossRef]
- Seibert, M.; Kurrle, N.; Schnütgen, F.; Serve, H. Amino acid sensory complex proteins in mTORC1 and macroautophagy regulation. Matrix Biol. 2021, 100, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.J.; Abraham, R.T.; Yu, K. Mammalian target of rapamycin: Discovery of rapamycin reveals a signaling pathway important for normal and cancer cell growth. Semin. Oncol. 2009, 36 (Suppl. S3), S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Roudsari, N.M.; Lashgari, N.-A.; Momtaz, S.; Abaft, S.; Jamali, F.; Safaiepour, P.; Narimisa, K.; Jackson, G.; Bishayee, A.; Rezaei, N. Inhibitors of the PI3K/Akt/mTOR pathway in prostate cancer chemoprevention and intervention. Pharmaceutics 2021, 13, 1195. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, S.I.; Pe Benito, R.; Henshall, S.M.; Horvath, L.G.; Kench, J.G. Expression of phosphorylated-mTOR during the development of prostate cancer. Prostate 2014, 74, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Etemad, S.; Rezaei, S.; Ziaolhagh, S.; Rajabi, R.; Rahmanian, P.; Abdi, S.; Koohpar, Z.K.; Rafiei, R.; Raei, B. Progress in targeting PTEN/PI3K/Akt axis in glioblastoma therapy: Revisiting molecular interactions. Biomed. Pharmacother. 2023, 158, 114204. [Google Scholar] [CrossRef] [PubMed]
- Ramezanidoraki, N.; Ouardi, D.E.; Le, M.; Moriceau, S.; Ahmadi, M.; Dossi, E.; Rolland, D.; Bun, P.; Le Pen, G.; Canaud, G. Activation of the PI3K/AKT/mTOR Pathway in Cajal–Retzius Cells Leads to Their Survival and Increases Susceptibility to Kainate-Induced Seizures. Int. J. Mol. Sci. 2023, 24, 5376. [Google Scholar] [CrossRef]
- Vicentini, C.; Cantù, C.; Antonello, D.; Simbolo, M.; Mafficini, A.; Luchini, C.; Rusev, B.; Porcaro, A.B.; Iacovelli, R.; Fassan, M. ERG alterations and mTOR pathway activation in primary prostate carcinomas developing castration-resistance. Pathol.-Res. Pract. 2018, 214, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, N.; Lima, T.B.; Mercaldi, G.F.; Nascimento, A.F.Z.; Silva, A.L.S.; Righetto, G.L.; Bar-Peled, L.; Shen, K.; Sabatini, D.M.; Gozzo, F.C.; et al. C7orf59/LAMTOR4 phosphorylation and structural flexibility modulate Ragulator assembly. FEBS Open Bio 2019, 9, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, R.; Wang, Z.; Wang, X.; Wang, F.; Ding, J. Structural basis for Ragulator functioning as a scaffold in membrane-anchoring of Rag GTPases and mTORC1. Nat. Commun. 2017, 8, 1394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lee, S.H.; Nie, L.; Huang, Y.; Zou, G.; Jung, Y.S.; Jun, S.; Zhang, J.; Lien, E.M.; Chen, J.; et al. Lysosomal TMEM9-LAMTOR4-controlled mTOR signaling integrity is required for mammary tumorigenesis. Cancer Commun. 2023, 43, 159–163. [Google Scholar] [CrossRef]
- Ding, D.; Blee, A.M.; Zhang, J.; Pan, Y.; Becker, N.A.; Maher, L.J., 3rd; Jimenez, R.; Wang, L.; Huang, H. Gain-of-function mutant p53 together with ERG proto-oncogene drive prostate cancer by beta-catenin activation and pyrimidine synthesis. Nat. Commun. 2023, 14, 4671. [Google Scholar] [CrossRef] [PubMed]
- Goschzik, T.; Gessi, M.; Denkhaus, D.; Pietsch, T. PTEN mutations and activation of the PI3K/Akt/mTOR signaling pathway in papillary tumors of the pineal region. J. Neuropathol. Exp. Neurol. 2014, 73, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, J.; Zhang, M.; Chen, N.; Sun, G.; Yang, Y.; Zhang, X.; Chen, J.; Shen, P.; Shi, M. The validation of the 2014 International Society of Urological Pathology (ISUP) grading system for patients with high-risk prostate cancer: A single-center retrospective study. Cancer Manag. Res. 2019, 11, 6521–6529. [Google Scholar] [CrossRef] [PubMed]
- Bartha, Á.; Győrffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | LAMTOR4 Low Risk (Score 0, 1, 2) | LAMTOR4 High Risk (Score 3) | p-Value |
|---|---|---|---|
| Gleason Score | |||
| ≤6 | 94 (37.6) | 11 (22.0) | 0.265 |
| 3 + 4 | 15 (6.0) | 5 (10.0) | |
| 4 + 3 | 24 (9.6) | 6 (12.0) | |
| 8 | 16 (6.4) | 5 (10.0) | |
| 9–10 | 101 (40.4) | 23 (46.0) | |
| PTEN Intensity | |||
| Score 0 (negative) | 78 (32.8) | 25 (48.1) | 0.037 |
| Score (1, 2, 3) (positive) | 160 (67.2) | 27 (51.9) | |
| ERG Expression | |||
| Negative | 183 (76.9) | 31 (59.6) | 0.01 |
| Positive | 55 (23.1) | 21 (40.4) |
| Variables | Overall Survival HR (95% CI) | p-Value | Cause-Specific Survival HR (95% CI) | p-Value |
|---|---|---|---|---|
| PTEN (Positive- score 1, 2 or 3) | ||||
| Negative (score 0) | 2.29 (1.70–3.08) | <0.0001 | 3.10 (2.05–4.68) | <0.0001 |
| ERG (Negative) | ||||
| Positive | 1.61 (1.18–2.22) | 0.003 | 1.65 (1.07–2.55) | 0.025 |
| GG 1 | ||||
| GG2 | 2.73 (1.145–5.13) | 0.002 | 12.18 (3.05–48.73) | <0.0001 |
| GG3 | 2.21 (1.24–3.95) | 0.008 | 3.91 (0.77–18.88) | 0.102 |
| GG4 | 3.59 (1.87–6.90) | <0.0001 | 18.22 (4.70–70.68) | <0.0001 |
| GG5 | 4.63 (3.08 -6.95) | <0.0001 | 30.95 (9.71–98.68) | <0.0001 |
| LAMTOR (Low risk Score 0, 1, 2) | ||||
| LAMTOR- high risk score 3 | 1.44 (1.01–2.05) | 0.047 | 1.71 (1.06–2.77) | 0.028 |
| Combination PTEN and LAMTOR (PTEN-score 1, 2, 3 and LAMTOR-score 0, 1, 2) | ||||
| PTEN-score 0 and LAMTOR-score 3 | 2.22 (1.37–3.59) | 0.001 | 3.46 (1.86–6.46) | <0.0001 |
| PTEN-score 0 and LAMTOR-score 0–2 | 2.50 (1.79–3.50) | <0.0001 | 3.36 (2.10–5.38) | <0.0001 |
| PTEN-score 1, 2, 3 and LAMTOR-score 3 | 1.52 (0.87–2.64) | 0.14 | 1.80 (0.83–3.89) | 0.137 |
| Combination PTEN and LAMTOR (PTEN-score 1, 2, 3 and LAMTOR-score 0, 1, 2) * | ||||
| PTEN-score 0 and LAMTOR-score 3 | 1.15 (0.67–1.99) | 0.61 | 1.23 (0.59–2.52) | 0.582 |
| PTEN-score 0 and LAMTOR-score 0–2 | 1.75 (1.23–2.49) | 0.002 | 1.92 (1.17 -3.17) | 0.01 |
| PTEN-score 1, 2, 3 and LAMTOR-score 3 | 1.55 (0.88–2.72) | 0.126 | 2.07 (0.94–4.55) | 0.071 |
| Combination ERG and LAMTOR (ERG- negative and LAMTOR- score 0–2) | ||||
| ERG positive and LAMTOR score 3 | 1.98 (1.18–3.31) | 0.01 | 2.54 (1.32–4.87) | 0.005 |
| ERG positive and LAMTOR score 0–2 | 1.57 (1.09–2.26) | 0.017 | 1.47 (0.87- 2.49) | 0.152 |
| ERG negative and LAMTOR score 3 | 1.26 (0.77–2.06) | 0.357 | 1.40 (0.73–2.69) | 0.313 |
| Combination ERG and LAMTOR (ERG- negative and LAMTOR- score 0–2) * | ||||
| ERG positive and LAMTOR score 3 | 1.13 (0.64–2.01) | 0.67 | 1.08 (0.51–2.31) | 0.834 |
| ERG positive and LAMTOR score 0–2 | 1.10 (0.75–1.62) | 0.625 | 0.87 (0.50–1.51) | 0.615 |
| ERG negative and LAMTOR score 3 | 1.05 (0.63–1.74) | 0.859 | 1.07 (0.54–2.13) | 0.84 |
| Variables | LAMTOR4 (Score 0, 1, 2) | LAMTOR4 (Score 3) | p-Value | Total |
|---|---|---|---|---|
| Age at diagnosis (years) | 75.99 (9.28) | 78.39 (8.67) | 0.081 | 76.40 (9.21) |
| Gleason Score | 0.084 | |||
| GS 6 or 7 | 82 (41.2) | 10 (26.3) | 92 (38.8) | |
| GS 8, 9 or 10 | 117 (58.8) | 28 (73.7) | 145 (61.2) | |
| Cancer subgroup | 0.015 | |||
| Incidental | 82 (31.5) | 10 (18.5) | 92 (29.3) | |
| Advanced | 83 (31.9) | 13 (24.1) | 96 (30.6) | |
| Castration-resistant | 95 (36.5) | 31 (57.4) | 126 (40.1) | |
| Metastasis | 0.112 | |||
| Yes | 28 (10.8) | 10 (18.5) | 38 (12.1) | |
| No | 232 (89.2) | 44 (81.5) | 276 (87.9) | |
| Hormone therapy | 0.016 | |||
| Prior to surgery | 42 (40.4) | 19 (65.5) | 72 (54.1) | |
| After surgery | 62 (59.6) | 10 (34.5) | 72 (54.1) | |
| Radiation Therapy | 0.743 | |||
| Prior to surgery | 18 (29.5) | 7 (33.3) | 25 (30.5) | |
| After surgery | 43 (70.5) | 14 (66.7) | 57 (69.5) | |
| Chemotherapy | 0.999 | |||
| Prior to surgery | 5 (12.9) | 1 (9.1) | 5 (11.9) | |
| After surgery | 27 (87.1) | 10 (90.9) | 37 (88.1) | |
| Deceased | 0.068 | |||
| Yes | 147 (57.0) | 38 (70.4) | 185 (59.3) | |
| No | 111 (43.0) | 16 (29.6) | 127 (40.7) | |
| Specific cause of death | 0.565 | |||
| Prostate cancer-specific death | 70 (27.1) | 22 (40.7) | 92 (29.5) | |
| Other causes or alive | 188 (72.9) | 32 (59.3) | 220 (70.5) | |
| Median follow-up (months) | 43 (1.5–101) | 34 (2.3–97.3) | 0.091 | 39.0 (1.5–101.2) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamallat, Y.; Alwazan, H.; Turko, R.; Dang, V.; Seyedi, S.; Ghosh, S.; Bismar, T.A. Elevated LAMTOR4 Expression Is Associated with Lethal Prostate Cancer and Its Knockdown Decreases Cell Proliferation, Invasion, and Migration In Vitro. Int. J. Mol. Sci. 2024, 25, 8100. https://doi.org/10.3390/ijms25158100
Gamallat Y, Alwazan H, Turko R, Dang V, Seyedi S, Ghosh S, Bismar TA. Elevated LAMTOR4 Expression Is Associated with Lethal Prostate Cancer and Its Knockdown Decreases Cell Proliferation, Invasion, and Migration In Vitro. International Journal of Molecular Sciences. 2024; 25(15):8100. https://doi.org/10.3390/ijms25158100
Chicago/Turabian StyleGamallat, Yaser, Huseen Alwazan, Rasoul Turko, Vincent Dang, Sima Seyedi, Sunita Ghosh, and Tarek A. Bismar. 2024. "Elevated LAMTOR4 Expression Is Associated with Lethal Prostate Cancer and Its Knockdown Decreases Cell Proliferation, Invasion, and Migration In Vitro" International Journal of Molecular Sciences 25, no. 15: 8100. https://doi.org/10.3390/ijms25158100