
Mapping of Human Polyomavirus in Renal Cell Carcinoma Tissues


 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Human Polyomaviruses—DNA PCR
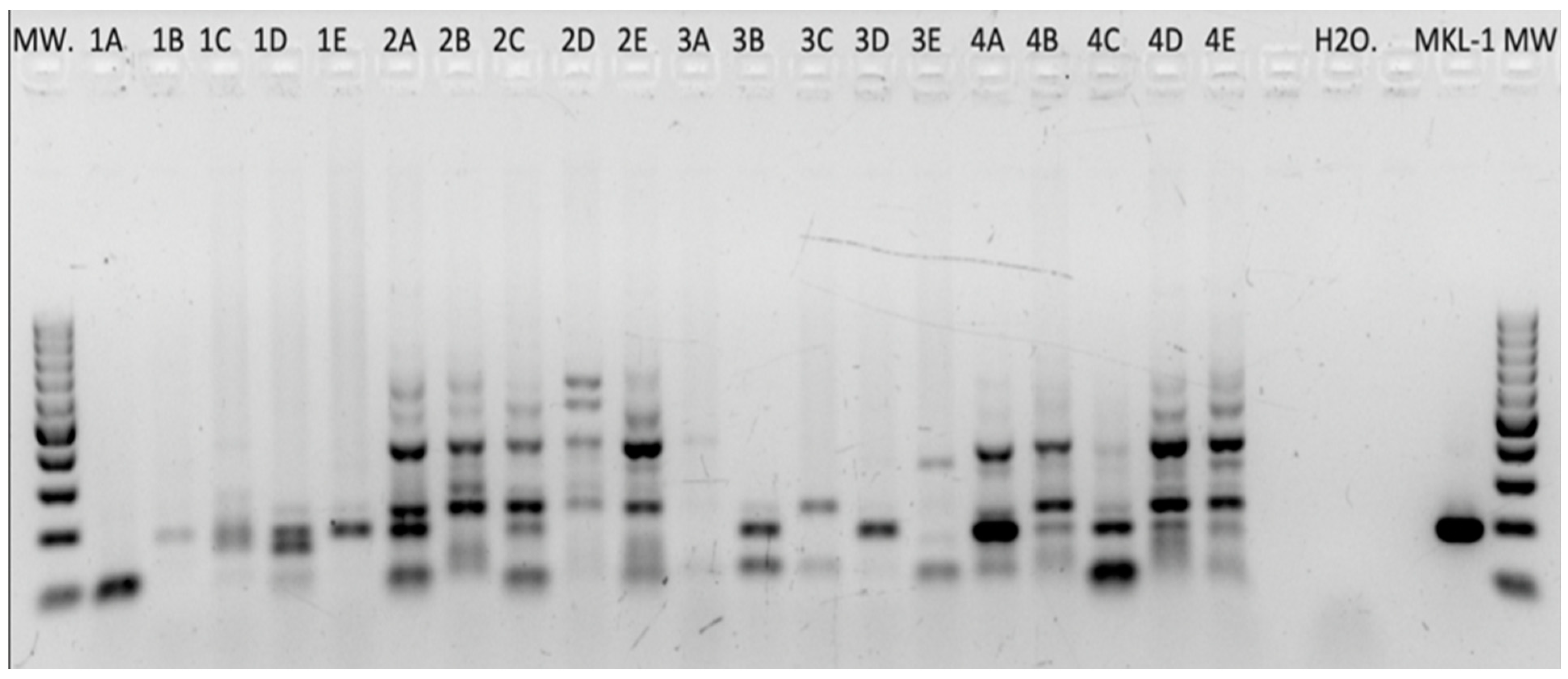
2.1.1. HPyV Consensus PCR
2.1.2. HPyV-Specific PCRs
2.1.3. Combined PCR Results (HPyV Consensus PCR and HPyV-Specific PCRs)
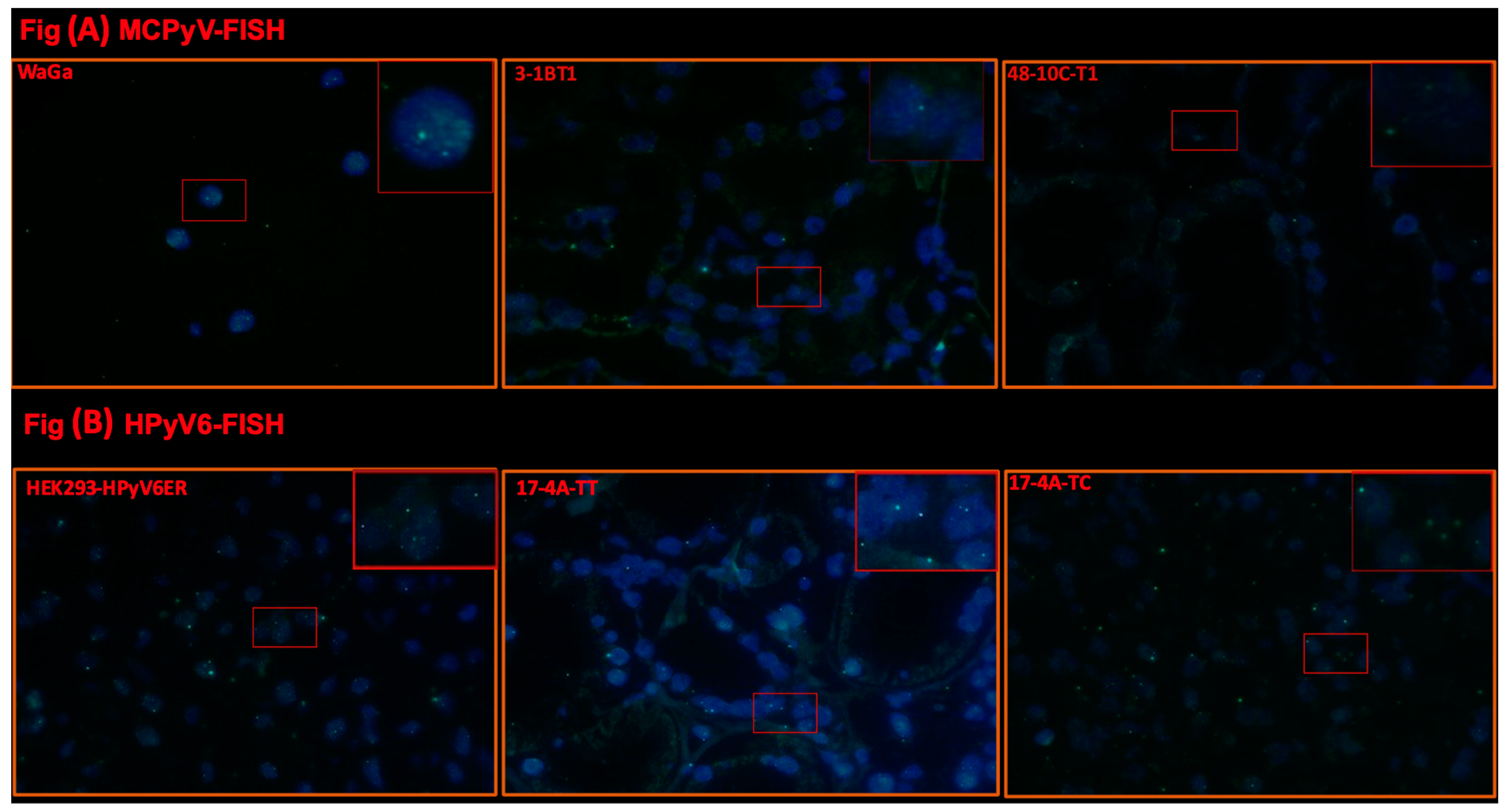
2.2. Fluorescence In Situ Hybridization (FISH)
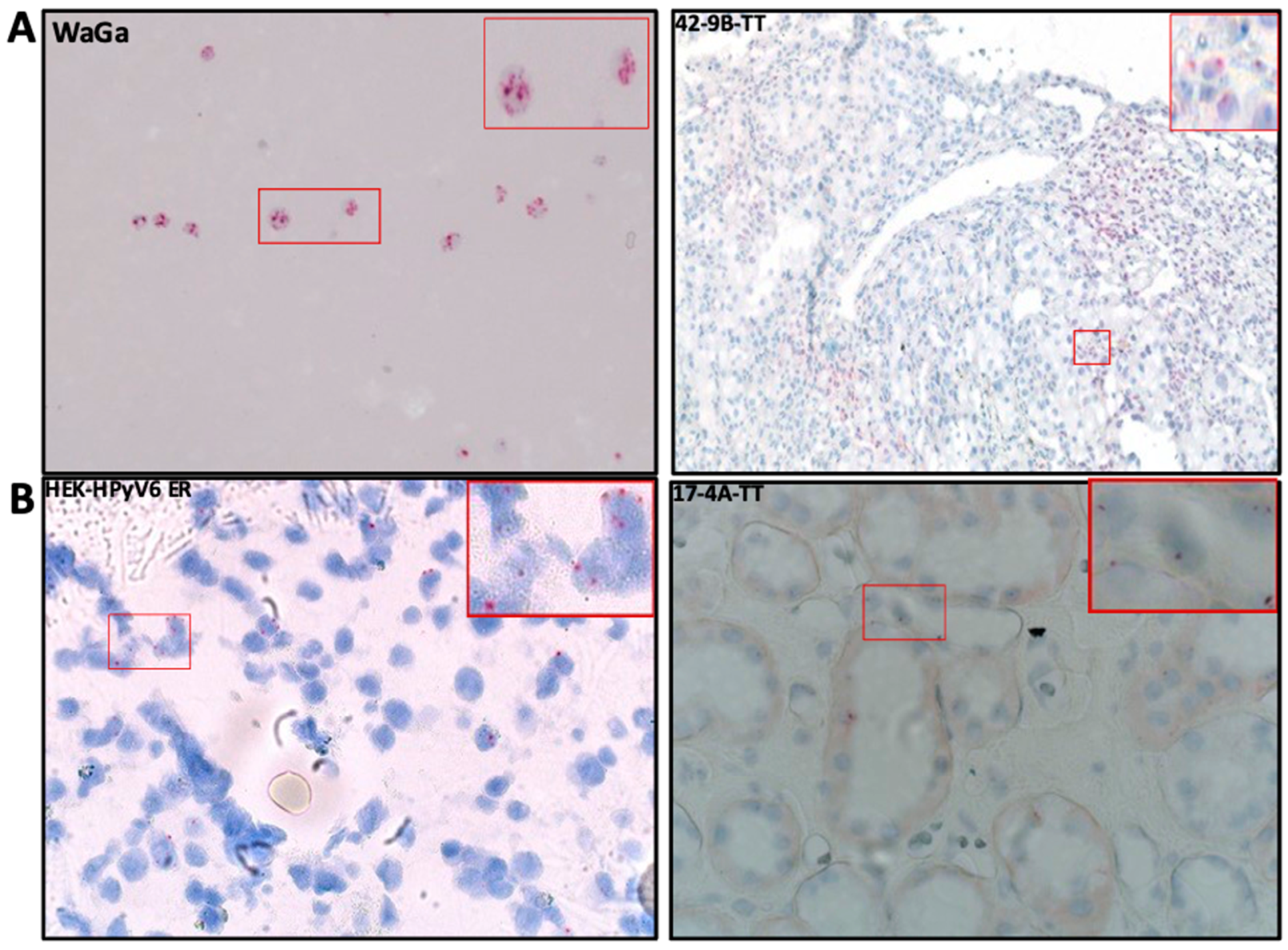
2.3. RNA In Situ Hybridization (RISH)
2.4. HPyV LT-Ag Immunohistochemistry
2.4.1. MCPyV-LTAg
2.4.2. PAb416 LTAg Immunoreactivity
3. Discussion
4. Materials and Methods
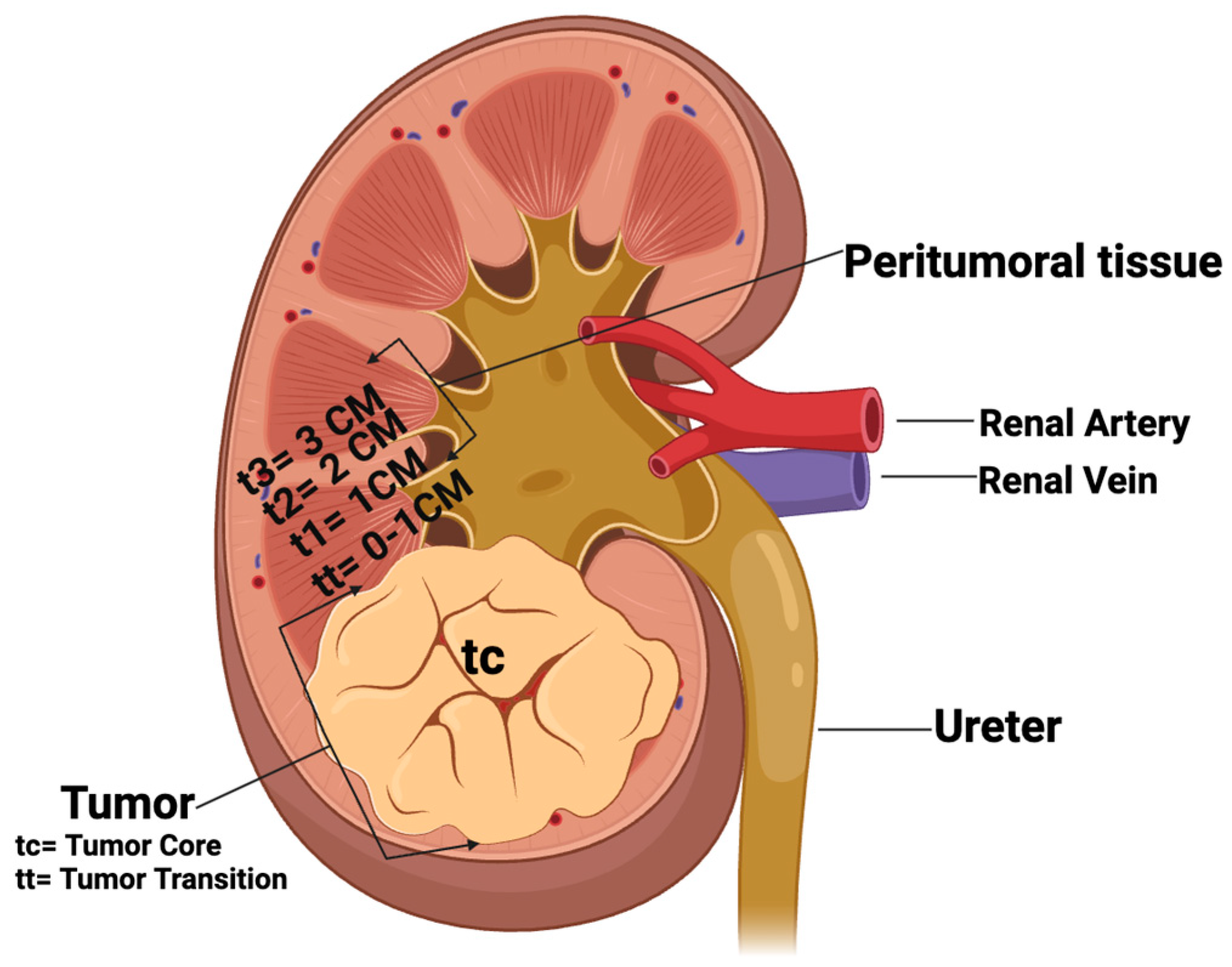
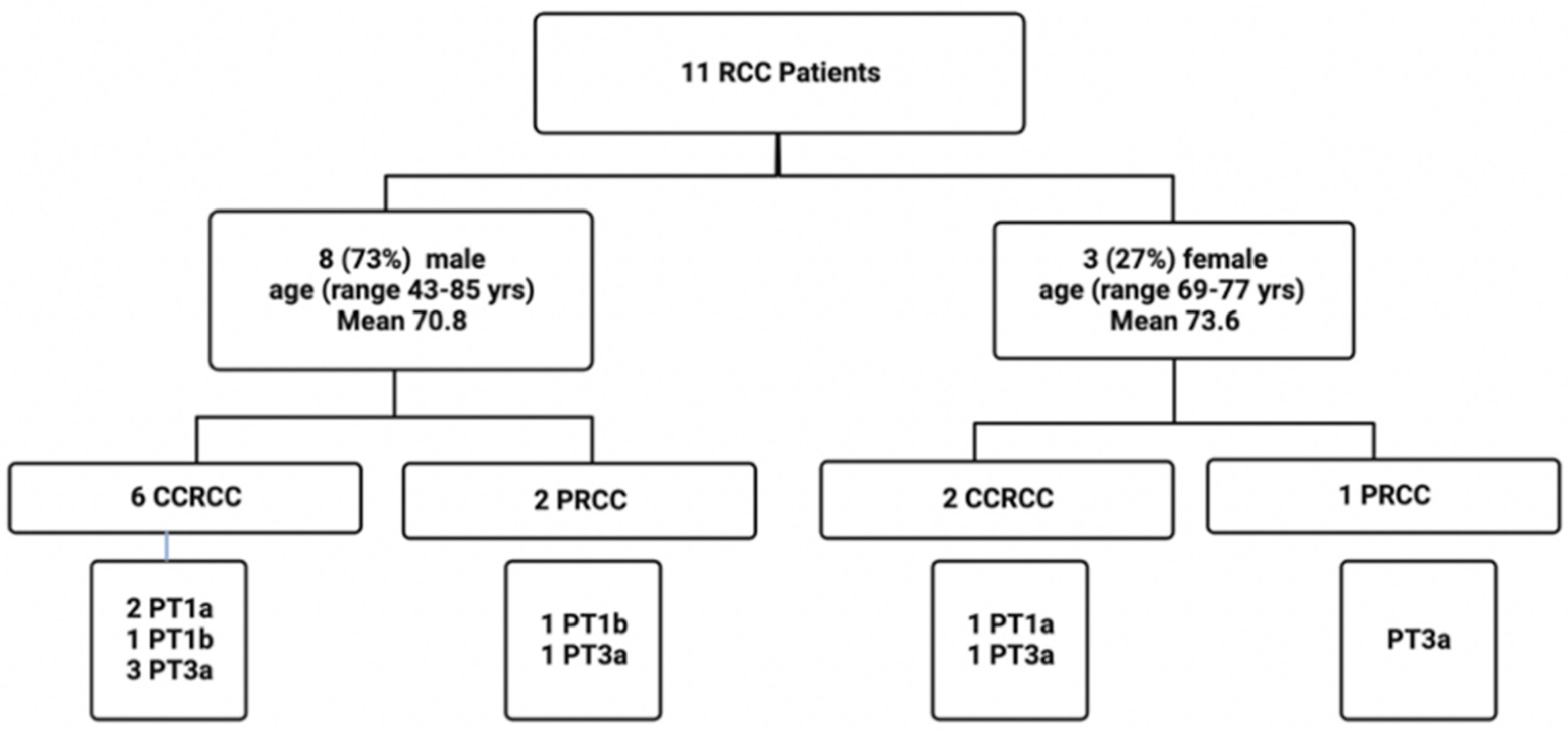
4.1. Patients and Tissues
4.2. DNA Extraction
4.3. HPyV PCR
4.3.1. Consensus PCR
4.3.2. HPyV-Specific PCRs
4.4. Sequence Analyses
4.5. Construction of HPyV6 ER Lentiviral Vector and Generation of HPyV6 ER-Expressing HEK Cell Line
4.6. Fluorescence In Situ Hybridization (FISH)
4.7. RNA In Situ Hybridization (RISH)
4.8. Immunohistochemistry (IHC)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Scelo, G.; Larose, T.L. Epidemiology and Risk Factors for Kidney Cancer. J. Clin. Oncol. 2018, 36, 3574–3581. [Google Scholar] [CrossRef] [PubMed]
- Kathuria-Prakash, N.; Drolen, C.; Hannigan, C.A.; Drakaki, A. Immunotherapy and Metastatic Renal Cell Carcinoma: A Review of New Treatment Approaches. Life 2021, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Trpkov, K.; Hes, O.; Williamson, S.R.; Adeniran, A.J.; Agaimy, A.; Alaghehbandan, R.; Amin, M.B.; Argani, P.; Chen, Y.B.; Cheng, L.; et al. New developments in existing WHO entities and evolving molecular concepts: The Genitourinary Pathology Society (GUPS) update on renal neoplasia. Mod. Pathol. 2021, 34, 1392–1424. [Google Scholar] [CrossRef] [PubMed]
- Engels, E.A.; Pfeiffer, R.M.; Fraumeni, J.F., Jr.; Kasiske, B.L.; Israni, A.K.; Snyder, J.J.; Wolfe, R.A.; Goodrich, N.P.; Bayakly, A.R.; Clarke, C.A.; et al. Spectrum of cancer risk among US solid organ transplant recipients. Jama 2011, 306, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Macleod, L.C.; Hotaling, J.M.; Wright, J.L.; Davenport, M.T.; Gore, J.L.; Harper, J.; White, E. Risk factors for renal cell carcinoma in the VITAL study. J. Urol. 2013, 190, 1657–1661. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.M.; Hofmann, J.N.; Cho, E.; Pollak, M.N.; Chow, W.H.; Purdue, M.P. Circulating levels of obesity-related markers and risk of renal cell carcinoma in the PLCO cancer screening trial. Cancer Causes Control 2017, 28, 801–807. [Google Scholar] [CrossRef]
- Webster, B.R.; Gopal, N.; Ball, M.W. Tumorigenesis Mechanisms Found in Hereditary Renal Cell Carcinoma: A Review. Genes 2022, 13, 2122. [Google Scholar] [CrossRef]
- Capitanio, U.; Bensalah, K.; Bex, A.; Boorjian, S.A.; Bray, F.; Coleman, J.; Gore, J.L.; Sun, M.; Wood, C.; Russo, P. Epidemiology of Renal Cell Carcinoma. Eur. Urol. 2019, 75, 74–84. [Google Scholar] [CrossRef]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global burden of cancers attributable to infections in 2012: A synthetic analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef] [PubMed]
- zur Hausen, H. Infections Causing Human Cancer; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]
- zur Hausen, H. Viruses in human cancers. Science 1991, 254, 1167–1173. [Google Scholar] [CrossRef]
- Kim, K.H.; Han, E.M.; Lee, E.S.; Park, H.S.; Kim, I.; Kim, Y.S. Epstein-Barr virus infection in sarcomatoid renal cell carcinoma tissues. BJU Int. 2005, 96, 547–552. [Google Scholar] [CrossRef]
- Farhadi, A.; Behzad-Behbahani, A.; Geramizadeh, B.; Sekawi, Z.; Rahsaz, M.; Sharifzadeh, S. High-risk human papillomavirus infection in different histological subtypes of renal cell carcinoma. J. Med. Virol. 2014, 86, 1134–1144. [Google Scholar] [CrossRef]
- Ma, Y.; Huang, Z.; Jian, Z.; Wei, X. The association between hepatitis C virus infection and renal cell cancer, prostate cancer, and bladder cancer: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 10833. [Google Scholar] [CrossRef] [PubMed]
- Bulut, Y.; Ozdemir, E.; Ozercan, H.I.; Etem, E.O.; Aker, F.; Toraman, Z.A.; Seyrek, A.; Firdolas, F. Potential relationship between BK virus and renal cell carcinoma. J. Med. Virol. 2013, 85, 1085–1089. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, M.; Szymanski, J.; Slavcheva, E.; Rao, A.; Kelly, A.; Jones, K.; Jaffers, G. BK virus associated renal cell carcinoma: Case presentation with optimized PCR and other diagnostic tests. Am. J. Transplant. 2007, 7, 1666–1671. [Google Scholar] [CrossRef]
- Pyöriä, L.; Pratas, D.; Toppinen, M.; Hedman, K.; Sajantila, A.; Perdomo, M.F. Unmasking the tissue-resident eukaryotic DNA virome in humans. Nucleic Acids Res. 2023, 51, 3223–3239. [Google Scholar] [CrossRef]
- Namdari, S.; Chong, P.P.; Behzad-Behbahani, A.; Geramizadeh, B.; Nazhvani, A.D.; Sekawi, Z.; Farhadi, A. Human herpesvirus 6A and 6B and polyomavirus JC and BK infections in renal cell carcinoma and their relationship with p53, p16INK4a, Ki-67, and nuclear factor-kappa B expression. Microbiol. Immunol. 2022, 66, 510–518. [Google Scholar] [CrossRef]
- Nickeleit, V.; Klimkait, T.; Binet, I.F.; Dalquen, P.; Del Zenero, V.; Thiel, G.; Mihatsch, M.J.; Hirsch, H.H. Testing for polyomavirus type BK DNA in plasma to identify renal-allograft recipients with viral nephropathy. N. Engl. J. Med. 2000, 342, 1309–1315. [Google Scholar] [CrossRef]
- DeCaprio, J.A.; Garcea, R.L. A cornucopia of human polyomaviruses. Nat. Rev. Microbiol. 2013, 11, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.D.; Field, A.M.; Coleman, D.V.; Hulme, B. New human papovavirus (B.K.) isolated from urine after renal transplantation. Lancet 1971, 1, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Baan, R.A.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Straif, K. Carcinogenicity of malaria and of some polyomaviruses. Lancet Oncol. 2012, 13, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Nickeleit, V.; Hirsch, H.H.; Binet, I.F.; Gudat, F.; Prince, O.; Dalquen, P.; Thiel, G.; Mihatsch, M.J. Polyomavirus infection of renal allograft recipients: From latent infection to manifest disease. J. Am. Soc. Nephrol. 1999, 10, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Nickeleit, V.; Singh, H.K. Polyomaviruses and disease: Is there more to know than viremia and viruria? Curr. Opin. Organ Transplant. 2015, 20, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.; Dobson, S. BK and JC virus: A review. J. Infect. 2014, 68 (Suppl. 1), S2–S8. [Google Scholar] [CrossRef] [PubMed]
- Alvarez Orellana, J.; Kwun, H.J.; Artusi, S.; Chang, Y.; Moore, P.S. Sirolimus and Other Mechanistic Target of Rapamycin Inhibitors Directly Activate Latent Pathogenic Human Polyomavirus Replication. J. Infect. Dis. 2021, 224, 1160–1169. [Google Scholar] [CrossRef]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and characterization of a novel putative human polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef]
- Schowalter, R.M.; Pastrana, D.V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel cell polyomavirus and two previously unknown polyomaviruses are chronically shed from human skin. Cell Host Microbe 2010, 7, 509–515. [Google Scholar] [CrossRef]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef]
- Wang, L.; Harms, P.W.; Palanisamy, N.; Carskadon, S.; Cao, X.; Siddiqui, J.; Patel, R.M.; Zelenka-Wang, S.; Durham, A.B.; Fullen, D.R.; et al. Age and Gender Associations of Virus Positivity in Merkel Cell Carcinoma Characterized Using a Novel RNA In Situ Hybridization Assay. Clin. Cancer Res. 2017, 23, 5622–5630. [Google Scholar] [CrossRef]
- Kassem, A.; Schöpflin, A.; Diaz, C.; Weyers, W.; Stickeler, E.; Werner, M.; Zur Hausen, A. Frequent detection of Merkel cell polyomavirus in human Merkel cell carcinomas and identification of a unique deletion in the VP1 gene. Cancer Res. 2008, 68, 5009–5013. [Google Scholar] [CrossRef] [PubMed]
- Rennspiess, D.; Pujari, S.; Keijzers, M.; Abdul-Hamid, M.A.; Hochstenbag, M.; Dingemans, A.M.; Kurz, A.K.; Speel, E.J.; Haugg, A.; Pastrana, D.V.; et al. Detection of human polyomavirus 7 in human thymic epithelial tumors. J. Thorac. Oncol. 2015, 10, 360–366. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.J.; Langerak, A.W.; Brüggemann, M.; Evans, P.A.; Hummel, M.; Lavender, F.L.; Delabesse, E.; Davi, F.; Schuuring, E.; García-Sanz, R.; et al. Design and standardization of PCR primers and protocols for detection of clonal immunoglobulin and T-cell receptor gene recombinations in suspect lymphoproliferations: Report of the BIOMED-2 Concerted Action BMH4-CT98-3936. Leukemia 2003, 17, 2257–2317. [Google Scholar] [CrossRef]
- Klufah, F.; Mobaraki, G.; Chteinberg, E.; Alharbi, R.A.; Winnepenninckx, V.; Speel, E.J.M.; Rennspiess, D.; Olde Damink, S.W.; Neumann, U.P.; Kurz, A.K.; et al. High Prevalence of Human Polyomavirus 7 in Cholangiocarcinomas and Adjacent Peritumoral Hepatocytes: Preliminary Findings. Microorganisms 2020, 8, 1125. [Google Scholar] [CrossRef]
- Haugg, A.M.; Rennspiess, D.; zur Hausen, A.; Speel, E.J.; Cathomas, G.; Becker, J.C.; Schrama, D. Fluorescence in situ hybridization and qPCR to detect Merkel cell polyomavirus physical status and load in Merkel cell carcinomas. Int. J. Cancer 2014, 135, 2804–2815. [Google Scholar] [CrossRef] [PubMed]
- Haugg, A.M.; Speel, E.J.; Pantulu, N.D.; Pallasch, C.; Kurz, A.K.; Kvasnicka, H.M.; Cathomas, G.; Wendtner, C.M.; zur Hausen, A. Fluorescence in situ hybridization confirms the presence of Merkel cell polyomavirus in chronic lymphocytic leukemia cells. Blood 2011, 117, 5776–5777. [Google Scholar] [CrossRef]
- Hopman, A.H.; Kamps, M.A.; Smedts, F.; Speel, E.J.; Herrington, C.S.; Ramaekers, F.C. HPV in situ hybridization: Impact of different protocols on the detection of integrated HPV. Int. J. Cancer 2005, 115, 419–428. [Google Scholar] [CrossRef]
- Hafkamp, H.C.; Manni, J.J.; Haesevoets, A.; Voogd, A.C.; Schepers, M.; Bot, F.J.; Hopman, A.H.; Ramaekers, F.C.; Speel, E.J. Marked differences in survival rate between smokers and nonsmokers with HPV 16-associated tonsillar carcinomas. Int. J. Cancer 2008, 122, 2656–2664. [Google Scholar] [CrossRef]
- Anderson, C.M.; Zhang, B.; Miller, M.; Butko, E.; Wu, X.; Laver, T.; Kernag, C.; Kim, J.; Luo, Y.; Lamparski, H.; et al. Fully Automated RNAscope In Situ Hybridization Assays for Formalin-Fixed Paraffin-Embedded Cells and Tissues. J. Cell Biochem. 2016, 117, 2201–2208. [Google Scholar] [CrossRef]
- Moshiri, A.S.; Doumani, R.; Yelistratova, L.; Blom, A.; Lachance, K.; Shinohara, M.M.; Delaney, M.; Chang, O.; McArdle, S.; Thomas, H.; et al. Polyomavirus-Negative Merkel Cell Carcinoma: A More Aggressive Subtype Based on Analysis of 282 Cases Using Multimodal Tumor Virus Detection. J. Investig. Dermatol. 2017, 137, 819–827. [Google Scholar] [CrossRef]
- Toptan, T.; Yousem, S.A.; Ho, J.; Matsushima, Y.; Stabile, L.P.; Fernández-Figueras, M.T.; Bhargava, R.; Ryo, A.; Moore, P.S.; Chang, Y. Survey for human polyomaviruses in cancer. JCI Insight 2016, 1, e85562. [Google Scholar] [CrossRef]
- Loyo, M.; Guerrero-Preston, R.; Brait, M.; Hoque, M.O.; Chuang, A.; Kim, M.S.; Sharma, R.; Liégeois, N.J.; Koch, W.M.; Califano, J.A.; et al. Quantitative detection of Merkel cell virus in human tissues and possible mode of transmission. Int. J. Cancer 2010, 126, 2991–2996. [Google Scholar] [CrossRef]
- Jiang, M.; Abend, J.R.; Tsai, B.; Imperiale, M.J. Early events during BK virus entry and disassembly. J. Virol. 2009, 83, 1350–1358. [Google Scholar] [CrossRef]
- Bersanelli, M.; Casartelli, C.; Buti, S.; Porta, C. Renal cell carcinoma and viral infections: A dangerous relationship? World J. Nephrol. 2022, 11, 1–12. [Google Scholar] [CrossRef]
- Garayeva, N.; Demir, E.; Dirim, A.B.; Safak, S.; Artan, A.S.; Ozluk, Y.; Kílícaslan, I.; Turkmen, A. Expression of JC virus in a kidney transplant recipient with renal cell carcinoma. Nefrologia 2024, 44, 288–289. [Google Scholar] [CrossRef]
- Knöll, A.; Stoehr, R.; Jilg, W.; Hartmann, A. Low frequency of human polyomavirus BKV and JCV DNA in urothelial carcinomas of the renal pelvis and renal cell carcinomas. Oncol. Rep. 2003, 10, 487–491. [Google Scholar] [CrossRef]
- Kenan, D.J.; Mieczkowski, P.A.; Burger-Calderon, R.; Singh, H.K.; Nickeleit, V. The oncogenic potential of BK-polyomavirus is linked to viral integration into the human genome. J. Pathol. 2015, 237, 379–389. [Google Scholar] [CrossRef]
- Kenan, D.J.; Mieczkowski, P.A.; Latulippe, E.; Côté, I.; Singh, H.K.; Nickeleit, V. BK Polyomavirus Genomic Integration and Large T Antigen Expression: Evolving Paradigms in Human Oncogenesis. Am. J. Transplant. 2017, 17, 1674–1680. [Google Scholar] [CrossRef] [PubMed]
- zur Hausen, H. Oncogenic DNA viruses. Oncogene 2001, 20, 7820–7823. [Google Scholar] [CrossRef] [PubMed]
- Zur Hausen, H. The search for infectious causes of human cancers: Where and why. Virology 2009, 392, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.S.; Chang, Y. Why do viruses cause cancer? Highlights of the first century of human tumour virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Howley, P.M. Gordon Wilson Lecture: Infectious Disease Causes of Cancer: Opportunities for Prevention and Treatment. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 117–132. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Gene | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | Products Size | Reference |
|---|---|---|---|---|---|
| Modified Degenerated | LTAg | GTAAAACGACGGCCAGTCTGYCCYTTYACATCCTCAA | CAGGAAACAGCTATGACGCCCYATHAAYAGTGGVAAAAC | 186 bp | Our group |
| HPyV7 | sTAg | AGATTTAGCTGTCCCCAAAG (3946–3965) | AAGAAGGCCAAAGAGTATGC (4108–4127) | 181 bp | [32] |
| HPyV6 | sTAg | ATCAGCTTCCACAGGTAGGC (3438–3457) | TTGCCTTCTCAAAAAGGAGC (3542–3561) | 123 bp | [32] |
| MCPyV | LT3 | TTGTCTCGCCAGCATTGTAG (571–590) | ATATAGGGGCCTCGTCAACC (860–879) | 308 bp | [33] |
| MCPyV | VP1 | TGGATCTAGGCCCTGATTTTT (3786–3806) | TTTGCCAGCTTACAGTGTGG (4118–4137) | 351 bp | [33] |
| MCPyV | M1/M2 | GGCATGCCTGTGAATTAGGA (1711–1730) | TTGCAGTAATTTGTAAGGGGGCT (1867–1889) | 178 bp | [35] |
| JCPyV | LTAg | AAGTCTTTAGGGTCTTCTAC (4254–4273) | GTGCCAACCTATGGAACAGA (4407–4429) | 176 bp | [21] |
| BKPyV | LTAg | GGTGCCAACCTATGGAACAG (4322–4338) | ACAGCAAAGCAGGCAAG (4548–4567) | 246 bp | [18] |
| Lab# * | Lab ID * | Tissue Resection | SCS (bp) | Gender | Age | Diagnosis | Cons. PCR | MCPyV PCR | HPyV6 PCR | HPyV7 PCR | JCV PCR | BKV PCR | FISH MCPyV | RISH MCPyV | FISH HPyV6 | RISH HPyV6 | CM2B4 IHC | PAb 416 IHC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M1/M2 | LT3 | VP1 | ||||||||||||||||||
| 1 | 1A | TC | 300 | M | 62 | CCRCC | − | − | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | − | − | |
| 2 | 1E | TT | 300 | M | 62 | CCRCC | HPyV6 | −− | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 3 | 1B | T1 | 300 | M | 62 | CCRCC | MCPyV | − | + | − | − | − | − | − | + | + | N.D. | N.D. | + | − |
| 4 | 1C | T2 | 300 | M | 62 | CCRCC | MCPyV | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − | |
| 5 | 1D | T3 | 300 | M | 62 | CCRCC | MCPyV | − | + | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 6 | 2D | TC | 500 | M | 70 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 7 | 2C | TT | 500 | M | 70 | CCRCC | − | − | − | − | − | − | − | + | N.D. | N.D. | N.D. | N.D. | − | + |
| 8 | 2A | T1 | 500 | M | 70 | CCRCC | MCPyV | − | − | − | − | − | + | + | N.D. | N.D. | N.D. | N.D. | − | ++ |
| 9 | 2E | T2 | 500 | M | 70 | CCRCC | − | − | − | − | + | − | − | − | N.D. | N.D. | + | N.D. | − | (+) |
| 10 | 2B | T3 | 500 | M | 70 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | (+) |
| 11 | 3A | TC | 300 | F | 75 | PRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 12 | 3B | TT | 300 | F | 75 | PRCC | HPyV6 | − | − | − | + | + | − | − | N.D. | N.D. | + | + | − | (+) |
| 13 | 3C | T1 | 300 | F | 75 | PRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 14 | 3D | T2 | 300 | F | 75 | PRCC | MCPyV | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 15 | 3E | T3 | 300 | F | 75 | PRCC | − | − | − | − | + | − | − | − | N.D. | N.D. | N.D. | N.D. | − | (+) |
| 16 | 4C | TC | 500 | M | 73 | CCRCC | WUPyV | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | + |
| 17 | 4A | TT | 500 | M | 73 | CCRCC | HPyV6 | − | − | − | + | − | − | − | N.D. | N.D. | + | + | + | ++ |
| 18 | 4B | T1 | 500 | M | 73 | CCRCC | HPyV7 | − | − | − | − | − | − | + | N.D. | N.D. | N.D. | N.D. | − | + |
| 19 | 4D | T2 | 500 | M | 73 | CCRCC | HPyV7 | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | (+) |
| 20 | 4E | T3 | 500 | M | 73 | CCRCC | HPyV7 | − | − | − | + | − | − | − | N.D. | N.D. | N.D. | N.D. | − | ++ |
| 21 | 5A | TC | 400 | F | 69 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 22 | 5B | TT | 400 | F | 69 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 23 | 5C | T1 | 400 | F | 69 | CCRCC | MCPyV | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 24 | 5D | T2 | 400 | F | 69 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 25 | 5E | T3 | 400 | F | 69 | CCRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 26 | 6A | TC | 300 | M | 77 | PRCC | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − | |
| 27 | 6B | TT | 300 | M | 77 | PRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 28 | 6C | T1 | 400 | M | 77 | PRCC | − | − | − | − | − | − | + | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 29 | 6D | T2 | 400 | M | 77 | PRCC | HPyV7 | − | − | − | − | − | + | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 30 | 6E | T3 | 400 | M | 77 | PRCC | MCPyV | + | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 31 | 7E | TC | 500 | M | 73 | PRCC | − | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | + |
| 32 | 7D | TT | 500 | M | 73 | PRCC | − | − | − | − | + | − | − | − | N.D. | N.D. | + | N.D. | + | ++ |
| 33 | 7C | T1 | 500 | M | 73 | PRCC | HPyV7 | − | + | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | ++ |
| 34 | 7B | T2 | 500 | M | 73 | PRCC | HPyV7 | − | − | − | − | − | − | + | N.D. | N.D. | N.D. | N.D. | − | ++ |
| 35 | 7A | T3 | 500 | M | 73 | PRCC | − | − | − | − | + | − | − | + | N.D. | N.D. | N.D. | N.D. | − | ++ |
| 36 | 8E | TC | 400 | M | 43 | CCRCC | MCPyV | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 37 | 8D | TT | 400 | M | 43 | CCRCC | MCPyV | − | −− | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | + | ++ |
| 38 | 8C | T1 | 400 | M | 43 | CCRCC | − | − | − | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | − | (+) |
| 39 | 8B | T2 | 400 | M | 43 | CCRCC | − | − | − | −− | −− | − | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 40 | 8A | T3 | 400 | M | 43 | CCRCC | HPyV7 | - | - | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | − | − |
| 41 | 9A | TC | 500 | F | 77 | CCRCC | HPyV7 | − | − | − | − | − | − | + | N.D. | N.D. | N.D. | N.D. | − | − |
| 42 | 9B | TT | 500 | F | 77 | CCRCC | MCPyV | + | + | − | − | − | − | − | + | + | N.D. | N.D. | + | − |
| 43 | 9C | T1 | 500 | F | 77 | CCRCC | MCPyV | − | − | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 44 | 9D | T2 | 500 | F | 77 | CCRCC | MCPyV | + | + | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − |
| 45 | 9E | T3 | 500 | F | 77 | CCRCC | MCPyV | − | + | − | − | − | − | N.D. | N.D. | N.D. | N.D. | + | − | |
| 46 | 10A | TC | 400 | M | 84 | CCRCC | MCPyV | + | + | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | (+) | − |
| 47 | 10B | TT | 400 | M | 84 | CCRCC | MCPyV | + | + | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | (+) | − |
| 48 | 10C | T1 | 400 | M | 84 | CCRCC | MCPyV | + | + | − | − | + | − | − | + | + | N.D. | N.D. | + | − |
| 49 | 10D | T2 | 400 | M | 84 | CCRCC | MCPyV | + | + | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | (+) | − |
| 50 | 10E | T3 | 400 | M | 84 | CCRCC | MCPyV | + | + | − | − | − | − | − | N.D. | N.D. | N.D. | N.D. | (+) | − |
| 51 | 11A | TC | 500 | M | 85 | CCRCC | MCPyV | + | + | + | − | − | − | + | N.D. | N.D. | N.D. | N.D. | + | − |
| 52 | 11B | TT | 500 | M | 85 | CCRCC | MCPyV | + | + | − | − | + | + | + | N.D. | N.D. | N.D. | N.D. | + | ++ |
| 53 | 11C | T1 | 500 | M | 85 | CCRCC | − | + | + | − | − | + | − | − | N.D. | N.D. | N.D. | N.D. | + | + |
| 54 | 11D | T2 | 500 | M | 85 | CCRCC | HPyV7 | + | + | + | − | − | + | − | N.D. | N.D. | N.D. | N.D. | (+) | + |
| 55 | 11E | T3 | 500 | M | 85 | CCRCC | MCPyV | + | − | − | − | + | + | − | N.D. | N.D. | N.D. | N.D. | + | + |
| Total | 34/55 | 13/55 | 15/55 | 2/55 | 7/55 | 11/55 | 6/55 | 8/55 | 3/3 | 3/3 | 4/4 | 2/2 | 27/55 | 22/55 | ||||||
| 34/55 | 17/55 | 7/55 | 11/55 | 6/55 | 8/55 | |||||||||||||||
| Total number of PCR positive tissue blocks | 34/55 | 35/55 | ||||||||||||||||||
| MCPyV PCR results total (consensus. PCR & MCPyV-specific PCR) | 24/55 (44%) | |||||||||||||||||||
| HPyV6 PCR results total (consensus PCR & HPyV6-specific PCR) | 8/55 (15%) | |||||||||||||||||||
| HPyV7 PCR results total (consensus PCR & HPyV7-specific PCR) | 19/55(35%) | |||||||||||||||||||
| JCPyV PCR results total (consensus PCR & JCPyV-specific PCR) | 6/55 (11%) | |||||||||||||||||||
| BKPyV PCR results total (consensus PCR & BKPyV-specific PCR) | 8/55 (15%) | |||||||||||||||||||
| Resection Sample | RCC | Tumor Transition | Non-Tumoral Tissue | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient# | Diagnosis | Gender | Age | Clinical Stage | TC 0 CM | TT 0–1 CM | T1 1 CM | T2 2 CM | T3 3 CM | |||||
| Consensus | Specific | Consensus | Specific | Consensus | Specific | Consensus | Specific | Consensus | Specific | |||||
| 1 | CCRCC | M | 62 | 1A | – | HPyV7 | HPyV6 | – | MCPyV | MCPyV | MCPyV | – | MCPyV | MCPyV & HPyV7 |
| 2 | CCRCC | M | 70 | 3A | – | – | – | BKV | MCPyV | BKV & JCV | – | HPyV6 | – | – |
| 3 | PRCC | F | 75 | 3A | – | – | HPyV6 | HPyV6 & HPyV7 | – | – | MCPyV | – | – | HPyV6 |
| 4 | CCRCC | M | 73 | 3A | WUPyV | – | HPyV6 | HPyV6 | HPyV7 | BKV | HPyV7 | – | HPyV7 | HPyV6 |
| 5 | CCRCC | F | 69 | 1A | – | – | – | – | MCPyV | – | – | – | – | – |
| 6 | PRCC | M | 77 | 3A | – | – | – | – | – | JCV | HPyV7 | JCV | MCPyV | MCPyV |
| 7 | PRCC | M | 73 | 1B | – | – | – | HPyV6 | – | MCPyV | HPyV7 | BKV | HPyV7 | HPyV6 & BKV |
| 8 | CCRCC | M | 43 | 1B | MCPyV | – | MCPyV | HPyV7 | – | HPyV7 | – | – | HPyV7 | – |
| 9 | CCRCC | F | 77 | 3A | HPyV7 | BKV | MCPyV | MCPyV | MCPyV | – | MCPyV | MCPyV | MCPyV | MCPyV |
| 10 | CCRCC | M | 84 | 1A | MCPyV | MCPyV | MCPyV | MCPyV & HPyV7 | MCPyV | MCPyV & HPyV7 | MCPyV | MCPyV | MCPyV | MCPyV |
| 11 | CCRCC | M | 85 | 3A | MCPyV | MCPyV & BKV | MCPyV | MCPyV, HPyV7, BKV & JCV | – | MCPyV & HPyV7 | HPyV7 | MCPyV & JCV | MCPyV | MCPyV, HPyV7 & JCV |
| Total HPyVs | 5/11 | 5\11 | 7/11 | 13/11 | 6/11 | 10/11 | 8/11 | 7/11 | 8/11 | 12/11 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mobaraki, G.; Shi, S.; Liu, D.; Smits, K.M.; Severens, K.; Lommen, K.; Rennspiess, D.; Speel, E.-J.M.; Winnepenninckx, V.; Klufah, F.; et al. Mapping of Human Polyomavirus in Renal Cell Carcinoma Tissues. Int. J. Mol. Sci. 2024, 25, 8213. https://doi.org/10.3390/ijms25158213
Mobaraki G, Shi S, Liu D, Smits KM, Severens K, Lommen K, Rennspiess D, Speel E-JM, Winnepenninckx V, Klufah F, et al. Mapping of Human Polyomavirus in Renal Cell Carcinoma Tissues. International Journal of Molecular Sciences. 2024; 25(15):8213. https://doi.org/10.3390/ijms25158213
Chicago/Turabian StyleMobaraki, Ghalib, Shuai Shi, Dan Liu, Kim M. Smits, Kim Severens, Kim Lommen, Dorit Rennspiess, Ernst-Jan M. Speel, Véronique Winnepenninckx, Faisal Klufah, and et al. 2024. "Mapping of Human Polyomavirus in Renal Cell Carcinoma Tissues" International Journal of Molecular Sciences 25, no. 15: 8213. https://doi.org/10.3390/ijms25158213
APA StyleMobaraki, G., Shi, S., Liu, D., Smits, K. M., Severens, K., Lommen, K., Rennspiess, D., Speel, E.-J. M., Winnepenninckx, V., Klufah, F., Samarska, I., & Hausen, A. z. (2024). Mapping of Human Polyomavirus in Renal Cell Carcinoma Tissues. International Journal of Molecular Sciences, 25(15), 8213. https://doi.org/10.3390/ijms25158213