
Metabolomic and Proteomic Analyses to Reveal the Role of Plant-Derived Smoke Solution on Wheat under Salt Stress
 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Metabolomic Analysis of PDS Solution
2.2. Morphological Analysis of Wheat Treated with PDS Solution under Salt Stress
2.3. Identification and Functional Investigation of Proteins in Wheat Treated with PDS Solution under Salt Stress
2.4. Immuno-Blot Analysis of Ascorbate Peroxidase and H+-ATPase in Wheat with Application of PDS Solution under Salt Stress
2.5. qRT-PCR Analysis of the Gene Encoding Bet v1 in Wheat with Application of PDS Solution under Salt Stress
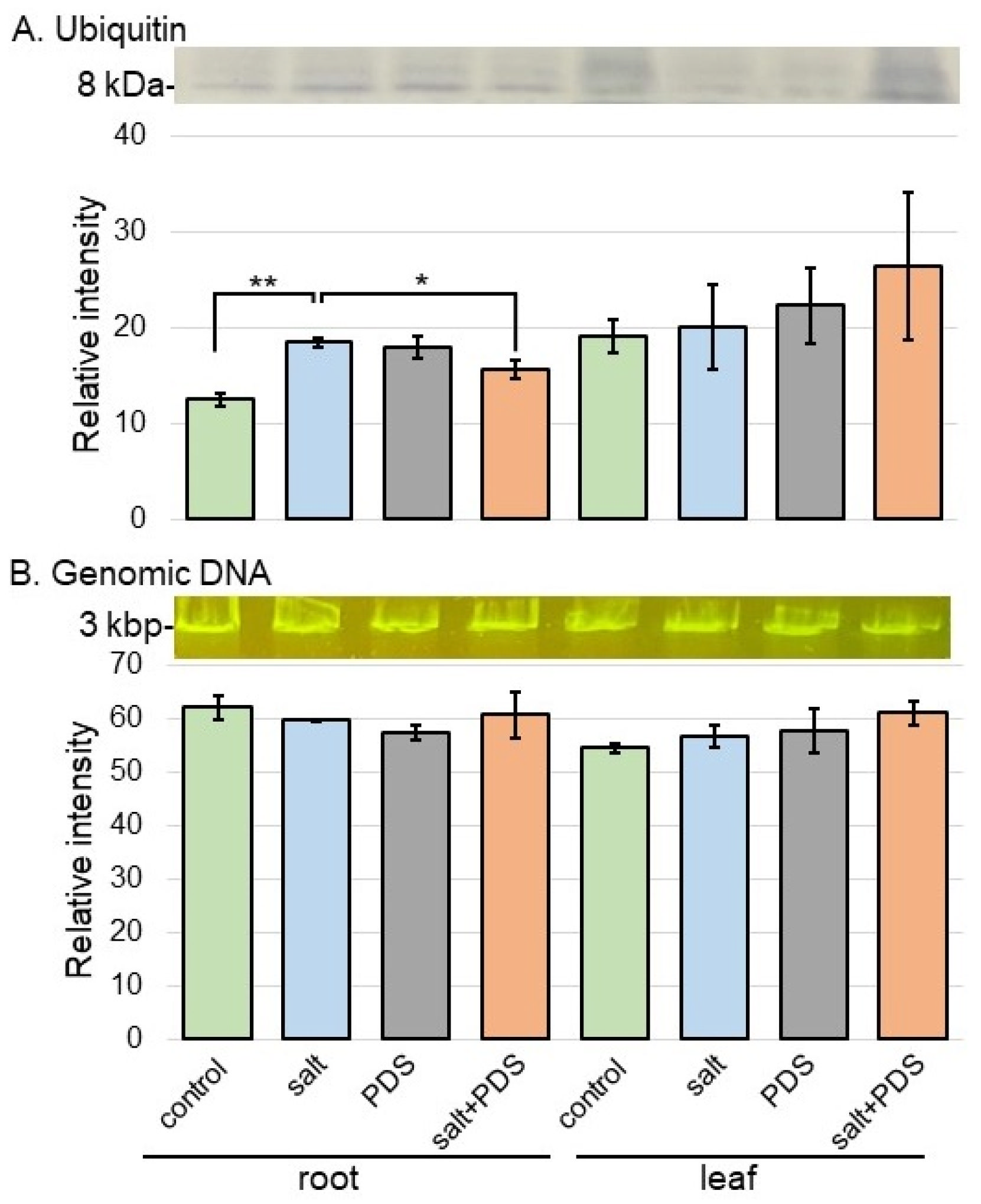
2.6. Analyses of Ubiquitin Accumulation and DNA Degradation in Wheat Treated with PDS Solution under Salt Stress
2.7. ATP Contents in Wheat with Application of PDS Solution under Salt Stress
3. Discussion
3.1. Fraction Containing Metabolites Involved in Flavonoid Biosynthesis Promotes Wheat Growth under Salt Stress
3.2. Reactive Oxygen Species Scavenging System Is Related to Salt-Tolerant Mechanism in Wheat Treated with PDS Solution
3.3. Energy Metabolism Is Related to Salt-Tolerant Mechanism in Wheat Treated with PDS Solution
3.4. PDS Solution Relieves Salt Secretion in Wheat under Salt Stress
3.5. PDS Solution Alleviates Promotion of Cell Death by Salt Stress
4. Materials and Methods
4.1. Preparation of PDS Solution
4.2. Metabolomic Analysis
4.2.1. Metabolites Extraction
4.2.2. UHPLC–MS Analysis
4.2.3. Analysis of Metabolomic Data
4.2.4. Differential Analysis of Metabolites Using Metabolomic Data
4.3. Plant Material and Treatment
4.4. Protein Extraction
4.5. Proteomic Analysis
4.5.1. Protein Enrichment, Reduction, Alkylation, and Digestion
4.5.2. Protein Identification Using Nano LC–MS/MS
4.5.3. MS Data Analysis
4.5.4. Differential Analysis of Proteins Using MS Data
4.6. Immuno-Blot Analysis
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Genomic DNA Extraction and Electrophoresis
4.9. Measurement of ATP Contents
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khatoon, A.; Rehman, S.U.; Aslam, M.M.; Jamil, M.; Komatsu, S. Plant-derived smoke affects biochemical mechanism on plant growth and seed germination. Int. J. Mol. Sci. 2020, 21, 7760. [Google Scholar] [CrossRef] [PubMed]
- Bose, U.; Juhász, A.; Broadbent, J.A.; Komatsu, S.; Colgrave, M.L. Multi-omics strategies for decoding smoke-assisted germination pathways and seed vigour. Int. J. Mol. Sci. 2020, 21, 7512. [Google Scholar] [CrossRef] [PubMed]
- Waters, M.T.; Nelson, D.C. Karrikin perception and signalling. New. Phytol. 2023, 237, 1525–1541. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Tripathi, D.K.; Roychoudhury, A. The karrikin “calisthenics”: Can compounds derived from smoke help in stress tolerance? Physiol. Plant. 2019, 165, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.P.; Baz, L.; Al-Babili, S. From carotenoids to strigolactones. J. Exp. Bot. 2018, 69, 2189–2204. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Lian, Y.; Wang, C. Comparing and contrasting the multiple roles of butenolide plant growth regulators: Strigolactones and karrikins in plant development and adaptation to abiotic stresses. Int. J. Mol. Sci. 2019, 20, 6270. [Google Scholar] [CrossRef] [PubMed]
- Otori, M.; Murashita, Y.; Rehman, S.; Komatsu, S. Proteomic study to understand promotive effects of plant-derived smoke on soybean root growth under flooding stress. Plant Mol. Biol. Rep. 2021, 39, 24–33. [Google Scholar] [CrossRef]
- Murashita, Y.; Nishiuchi, T.; Rehman, S.U.; Komatsu, S. Subcellular proteomics to understand promotive effect of plant-derived smoke solution on soybean root. Proteomes 2021, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Kobayashi, T.; Zhu, W.; Imai, H.; Zhao, R.; Ohno, T.; Rehman, S.U.; Uemura, M.; Tian, J.; Komatsu, S. Plant-derived smoke enhances plant growth through ornithine-synthesis pathway and ubiquitin-proteasome pathway in soybean. J. Proteom. 2020, 221, 103781. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Yamaguchi, T.; Sunohara, Y.; Matsumoto, H.; Komatsu, S. Proteomic analysis of the effect of plant-derived smoke on soybean during recovery from flooding stress. J. Proteom. 2018, 181, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Kimura, T.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K. Proteomic analysis reveals salt-tolerant mechanism in soybean applied with plant-derived smoke solution. Int. J. Mol. Sci. 2023, 24, 13734. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K.; Rehman, S.U.; Ohno, T. Morphological, biochemical, and proteomic analyses to understand the promotive effects of plant-derived smoke solution on wheat growth under flooding stress. Plants 2022, 11, 1508. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A.J. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Wang, D.; Liang, Y.; Liu, L.; Huang, J.; Yin, Z. Crop production on the Chinese Loess Plateau under 1.5 and 2.0 °C global warming scenarios. Sci. Total Environ. 2023, 11, 166158. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.L.; Liu, M.Y.; Ma, D.F.; Wu, J.W.; Li, S.L.; Zhu, Y.X.; Han, B. Identification of circular RNAs and their targets during tomato fruit ripening. Postharvest Biol. Technol. 2018, 136, 90–98. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Oldřich Urban, M.; Kholová, J.; Tom Prášil, I. Breeding for enhanced drought resistance in barley and wheat-drought-associated traits, genetic resources and their potential utilization in breeding programmes. Czech J. Genet. Plant Breed. 2014, 50, 247–261. [Google Scholar] [CrossRef]
- Huang, S.; Gill, S.; Ramzan, M.; Ahmad, M.Z.; Danish, S.; Huang, P.; Al Obaid, S.; Alharbi, S.A. Uncovering the impact of am fungi on wheat nutrient uptake, ion homeostasis, oxidative stress, and antioxidant defense under salinity stress. Sci. Rep. 2023, 13, 8249. [Google Scholar] [CrossRef] [PubMed]
- Sharaya, R.; Gill, R.; Kalwan, G.; Naeem, M.; Tuteja, N.; Gill, S.S. Plant-microbe interaction mediated salinity stress tolerance for sustainable crop production. S. Afr. J. Bot. 2023, 161, 454–471. [Google Scholar] [CrossRef]
- Bazihizina, N.; Colmer, T.D.; Barrett-Lennard, E.G. Response to non-uniform salinity in the root zone of the Halophyte Atriplex Nummularia: Growth, photosynthesis, water relations and tissue ion concentrations. Ann. Bot. 2009, 104, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Khan, A.L.; Ali, L.; Hussain, J.; Waqas, M.; Al-Harrasi, A.; Imran, Q.M.; Kim, Y.H.; Kang, S.M.; Yun, B.W.; et al. Hydroquinone; a novel bioactive compound from plant-derived smoke can cue seed germination of lettuce. Front. Chem. 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Staszak-Kozinski, L.; Davidson, R. Up in smoke: I. Smoke-derived germination cues for postfire annual, Nicotiana attenuata Torr. Ex. Watson. J. Chem. Ecol. 1994, 20, 2345–2371. [Google Scholar] [CrossRef] [PubMed]
- van Staden, J.; Drewes, F.; Jäger, A. The search for germination stimulants in plant-derived smoke extracts. S. Afr. J. Bot. 1995, 61, 260–263. [Google Scholar] [CrossRef]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A compound from smoke that promotes seed germination. Science 2004, 305, 977. [Google Scholar] [CrossRef] [PubMed]
- Sparg, S.G.; Kulkarni, M.G.; Light, M.E.; Van Staden, J. Improving seedling vigour of indigenous medicinal plants with smoke. Bioresour. Technol. 2005, 96, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.G.; Ascough, G.D.; Verschaeve, L.; Baeten, K.; Arruda, M.P.; Van Staden, J. Effect of smoke-water and a smoke-isolated butenolide on the growth and genotoxicity of commercial onion. Sci. Hortic. 2010, 124, 434–439. [Google Scholar] [CrossRef]
- van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post-germination effects of the smoke-derived compound 3-methyl-2H-furo [2,3-c] pyran-2-one, and its potential as a preconditioning agent. Field Crops. Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Ghitti, E.; Rolli, E.; Crotti, E.; Borin, S. Flavonoids are intra- and inter-kingdom modulator signals. Microorganisms 2022, 10, 2479. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Parveen, A.; Hussain, S.; Hussain, I.; Rasheed, R. Investigating the role of different maize (Zea mays L.) cultivars by studying morpho-physiological attributes in chromium-stressed environment. Environ. Sci. Pollut. Res. 2022, 29, 72886–72897. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Badola, P.K.; Gautam, H.; Trivedi, P.K. Heterologous expression of Arabidopsis miR858 modulates biosynthesis of secondary metabolites and affects drought tolerance in tobacco. Plant Cell Tissue Organ Cult. 2023, 152, 287–298. [Google Scholar] [CrossRef]
- Guo, Z.; Yuan, X.; Li, T.; Wang, S.; Yu, Y.; Liu, C.; Duan, C. Integrated transcriptomic and metabolomic analysis reveals the molecular regulatory mechanism of flavonoid biosynthesis in maize roots under lead stress. Int. J. Mol. Sci. 2024, 25, 6050. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ahammed, G.J. Hormonal regulation of anthocyanin biosynthesis for improved stress tolerance in plants. Plant Physiol. Biochem. 2023, 201, 107835. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Zhang, S.; Li, Q.; Xiang, G.; Zhao, Y.; Wei, F.; Zhang, G.; Yang, S.; Hao, B. Genome-wide profiling of WRKY genes involved in flavonoid biosynthesis in Erigeron breviscapus. Front. Plant Sci. 2024, 15, 1412574. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Todorova, D. Antioxidant machinery and glyoxalase system regulation confers salt stress tolerance to wheat (Triticum aestivum L.) plants treated with melatonin and salicylic Acid. J. Soil Sci. Plant Nutr. 2022, 22, 3527–3540. [Google Scholar] [CrossRef]
- Ben Saad, R.; Ben Romdhane, W.; Wiszniewska, A.; Baazaoui, N.; Taieb Bouteraa, M.; Chouaibi, Y.; Alfaifi, M.Y.; Kačániová, M.; Čmiková, N.; Ben Hsouna, A.; et al. Rosmarinus officinalis L. essential oil enhances salt stress tolerance of durum wheat seedlings through ROS detoxification and stimulation of antioxidant defense. Protoplasma 2024. [Google Scholar] [CrossRef]
- Hameed, A.; Maqsood, W.; Hameed, A.; Qayyum, M.A.; Ahmed, T.; Farooq, T. Chitosan nanoparticles encapsulating curcumin counteract salt-mediated ionic toxicity in wheat seedlings: An ecofriendly and sustainable approach. Environ. Sci. Pollut. Res. Int. 2024, 31, 8917–8929. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Hu, Y.; Song, J.; Feng, H.; Wang, J.; Chen, L.; Wang, L.; Diao, X.; Wan, Y.; Liu, S.; et al. Transcriptome sequencing and metabolome analysis reveals the molecular mechanism of drought stress in millet. Int. J. Mol. Sci. 2022, 23, 10792. [Google Scholar] [CrossRef] [PubMed]
- Okay, S.; Derelli, E.; Unver, T. Transcriptome-wide identification of bread wheat WRKY transcription factors in response to drought stress. Mol. Genet. Genom. 2014, 289, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Lee, H.J.; Song, Y.H.; Kim, H.J. Effect of salt stress on the growth, mineral contents, and metabolite profiles of spinach. J. Sci. Food. Agric. 2021, 101, 3787–3794. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Leng, B.Y.; Wang, B.S. Progress in studying salt secretion from the salt glands in recretohalophytes: How do plants secrete salt? Front. Plant Sci. 2016, 7, 977. [Google Scholar] [CrossRef] [PubMed]
- Marriboina, S.; Sekhar, K.M.; Subramanyam, R.; Reddy, A.R. Physiological, biochemical, and root proteome networks revealed new insights into salt tolerance mechanisms in Pongamia pinnata (L.) Pierre. Front. Plant Sci. 2022, 12, 771992. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.S.; Zhu, J.K.; Bressan, R.A.; Hasegawa, P.M.; Shi, H.H. Reactive oxygen species mediate Na+-induced SOS1 mRNA stability in Arabidopsis. Plant J. 2008, 53, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Jiang, L.; Lei, T.; Ouyang, K.; Liu, C.; Zhao, Z.; Li, Y.; Yang, L.; Li, J.; Yi, S.; et al. Increasing Ca2+ accumulation in salt glands under salt stress increases stronger selective secretion of Na+ in Plumbago auriculata tetraploids. Front. Plant Sci. 2024, 15, 1376427. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; El-Sharkawy, I.; Sherif, S. Salt stress signals on demand: Cellular events in the right context. Int. J. Mol. Sci. 2020, 21, 3918. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Takeda, S.; Nick, P. Life and death under salt stress: Same players, different timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.; Lv, Y.; Cao, B.L.; Chen, Z.J.; Xu, K. Physiological and molecular mechanism of ginger Zingiber officinale Roscoe seedling response to salt stress. Front. Plant Sci. 2023, 14, 1073434. [Google Scholar] [CrossRef] [PubMed]
- Tieu, A.; Plummer, J.A.; Dixon, K.A.; Sivasithamparam, K.; Sieler, I.M. Germination of four species of native Western Australian plants using plant-derived smoke. Aust. J. Bot. 1999, 47, 207–219. [Google Scholar] [CrossRef]
- Liu, Y.; Kong, Z.; Liu, J.; Zhang, P.; Wang, Q.; Huan, X.; Li, L.; Qin, P. Non-targeted metabolomics of quinoa seed filling period based on liquid chromatography-mass spectrometry. Food Res. Int. 2020, 137, 109743. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Spigelman, A.F.; MacDonald, P.E.; Wishart, D.S.; Li, S.; et al. MetaboAnalyst 6.0: Towards a unified platform for metabolomics data processing, analysis and interpretation. Nucleic Acids Res. 2024, 52, W398–W406. [Google Scholar] [CrossRef]
- Komatsu, S.; Han, C.; Nanjo, Y.; Altaf-Un-Nahar, M.; Wang, K.; He, D.; Yang, P. Label-free quantitative proteomic analysis of abscisic acid effect in early-stage soybean under flooding. J. Proteome Res. 2013, 12, 4769–4784. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Yamamoto, A.; Nakamura, T.; Nouri, M.Z.; Nanjo, Y.; Nishizawa, K.; Furukawa, K. Comprehensive analysis of mitochondria in roots and hypocotyls of soybean under flooding stress using proteomics and metabolomics techniques. J. Proteome Res. 2011, 10, 3993–4004. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidium thiocyanate–phenol–chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Côté, R.G.; Csordas, A.; Dianes, J.A.; Fabregat, A.; Foster, J.M.; Griss, J.; Alpi, E.; Birim, M.; Contell, J.; et al. The PRoteomics IDEntifications (PRIDE) database and associated tools: Status in 2013. Nucleic Acids Res. 2013, 41, D1063–D1069. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef] [PubMed]
- Ara, T.; Kodama, Y.; Tokimatsu, T.; Fukuda, A.; Kosuge, T.; Mashima, J.; Tanizawa, Y.; Tanjo, T.; Ogasawara, O.; Fujisawa, T.; et al. DDBJ update in 2023: The MetaboBank for metabolomics data and associated metadata. Nucleic Acids Res. 2024, 52, D67–D71. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komatsu, S.; Diniyah, A.; Zhu, W.; Nakano, M.; Rehman, S.U.; Yamaguchi, H.; Hitachi, K.; Tsuchida, K. Metabolomic and Proteomic Analyses to Reveal the Role of Plant-Derived Smoke Solution on Wheat under Salt Stress. Int. J. Mol. Sci. 2024, 25, 8216. https://doi.org/10.3390/ijms25158216
Komatsu S, Diniyah A, Zhu W, Nakano M, Rehman SU, Yamaguchi H, Hitachi K, Tsuchida K. Metabolomic and Proteomic Analyses to Reveal the Role of Plant-Derived Smoke Solution on Wheat under Salt Stress. International Journal of Molecular Sciences. 2024; 25(15):8216. https://doi.org/10.3390/ijms25158216
Chicago/Turabian StyleKomatsu, Setsuko, Azzahrah Diniyah, Wei Zhu, Masataka Nakano, Shafiq Ur Rehman, Hisateru Yamaguchi, Keisuke Hitachi, and Kunihiro Tsuchida. 2024. "Metabolomic and Proteomic Analyses to Reveal the Role of Plant-Derived Smoke Solution on Wheat under Salt Stress" International Journal of Molecular Sciences 25, no. 15: 8216. https://doi.org/10.3390/ijms25158216