
Kuwanon C Inhibits Tumor Cell Proliferation and Induces Apoptosis by Targeting Mitochondria and Endoplasmic Reticulum


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Inhibition of HeLa Cell Viability by Kuwanon C
2.2. Suppression of HeLa Cell Migration and Proliferation by Kuwanon C
2.3. Induction of Reactive Oxygen Species Generation in HeLa Cells by Kuwanon C
2.4. Induction of Apoptosis in HeLa Cells by Kuwanon C
2.5. Effect of Kuwanon C on Mitochondrial Membrane Potential
2.6. Modulation of the Cell Cycle by Kuwanon C in HeLa Cells
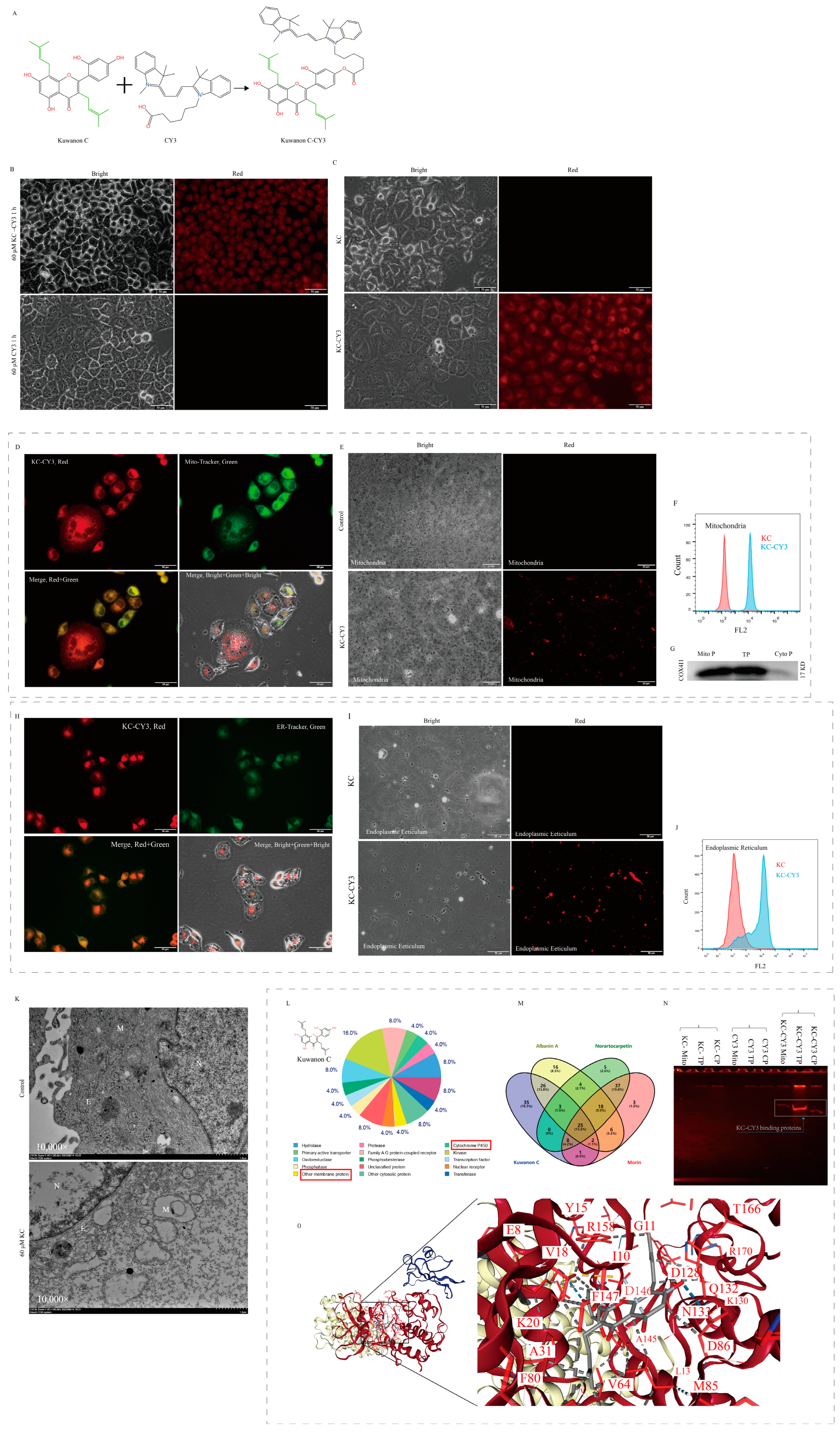
2.7. Molecular Targets of Kuwanon C
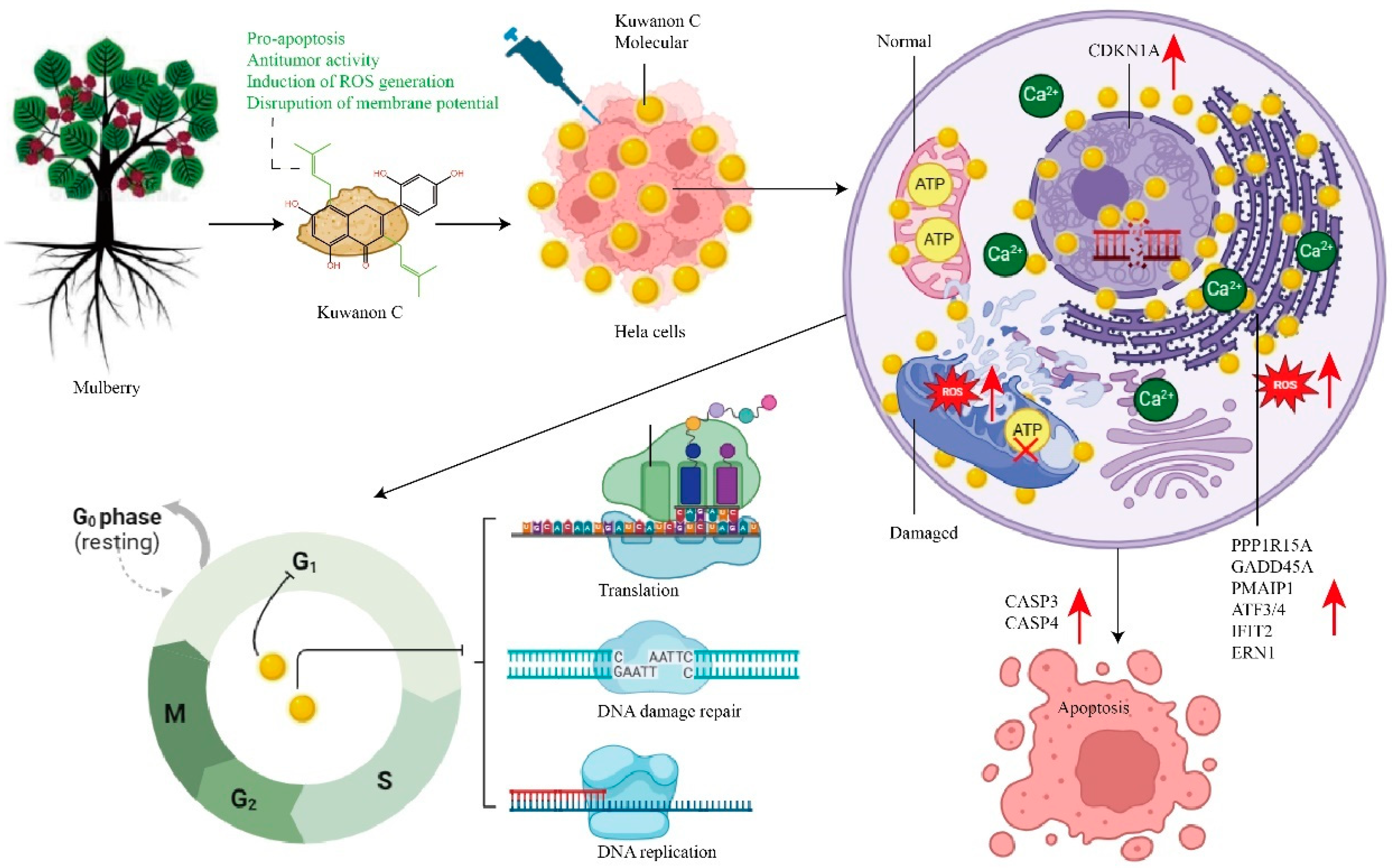
3. Discussion
3.1. Disruption of Tumor Cell Energy Metabolism and Protein Synthesis by Kuwanon C
3.2. Impact of Isopentenyl Groups on the Antitumor Efficacy of Kuwanon C
3.3. Cellular Level Comparison of Kuwanon C with Established Clinical Anticancer Agents
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Cell Culture
4.3. Cell Viability Assay
4.4. Wound Healing Assays
4.5. Colony Formation Assay
4.6. Transmission Electron Microscopy
4.7. 5-Ethynyl-2′-Deoxyuridine (EdU) Staining
4.8. Kuwanon C and Morin Red Fluorescent Group CY3 Labeling
4.9. Bioinformatic Analysis and Mass Spectrometric Characterization of Target Molecules for Kuwanon C Action
4.10. Molecular Docking
4.11. Mitochondrial Membrane Potential Measurement
4.12. Mitochondrial Staining
4.13. Endoplasmic Reticulum Staining
4.14. HeLa Cell Mitochondria Extraction and Analysis
4.15. ROS Assays
4.16. Transcriptome Analysis
4.17. Cellular Calcium Content Analysis
4.18. Gene Expression Analyses
4.19. Western Blotting Analyses
4.20. Cell Cycle Assay
4.21. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer. J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fontham, E.T.H.; Wolf, A.M.D.; Church, T.R.; Etzioni, R.; Flowers, C.R.; Herzig, A.; Guerra, C.E.; Oeffinger, K.C.; Shih, Y.T.; Walter, L.C.; et al. Cervical cancer screening for individuals at average risk: 2020 guideline update from the American Cancer Society. CA Cancer. J. Clin. 2020, 70, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [PubMed]
- Crunkhorn, S. Combination therapy combats aggressive breast cancer. Nat. Rev. Drug. Discov. 2020, 19, 310. [Google Scholar] [CrossRef]
- Melero, I.; Berman, D.M.; Aznar, M.A.; Korman, A.J.; Pérez Gracia, J.L.; Haanen, J. Evolving synergistic combinations of targeted immunotherapies to combat cancer. Nat. Rev. Cancer. 2015, 15, 457–472. [Google Scholar] [CrossRef]
- Furue, H. Chemotherapy cancer treatment during the past sixty years. Gan Kagaku Ryoho Cancer Chemother. 2003, 30, 1404–1411. [Google Scholar]
- Pérez-Herrero, E.; Fernández-Medarde, A. Advanced targeted therapies in cancer: Drug nanocarriers, the future of chemotherapy. Eur. J. Pharm. Biopharm. 2015, 93, 52–79. [Google Scholar] [CrossRef] [PubMed]
- Carcache de Blanco, E.J.; Addo, E.M.; Rakotondraibe, H.L.; Soejarto, D.D.; Kinghorn, A.D. Strategies for the discovery of potential anticancer agents from plants collected from Southeast Asian tropical rainforests as a case study. Nat. Prod. Rep. 2023, 40, 1181–1197. [Google Scholar] [CrossRef]
- Kuo, R.Y.; Qian, K.; Morris-Natschke, S.L.; Lee, K.H. Plant-derived triterpenoids and analogues as antitumor and anti-HIV agents. Nat. Prod. Rep. 2009, 26, 1321–1344. [Google Scholar] [CrossRef]
- Noble, R.L.; Beer, C.T.; Cutts, J.H. Role of chance observations in chemotherapy: Vinca rosea. Ann. N. Y. Acad. Sci. 1958, 76, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hansen, L.G.; Gudich, O.; Viehrig, K.; Lassen, L.M.M.; Schrübbers, L.; Adhikari, K.B.; Rubaszka, P.; Carrasquer-Alvarez, E.; Chen, L.; et al. A microbial supply chain for production of the anti-cancer drug vinblastine. Nature 2022, 609, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Gao, X.; Duan, L.; Zhang, W.; Sun, Y.F. Mulberrin (Mul) reduces spinal cord injury (SCI)-induced apoptosis, inflammation and oxidative stress in rats via miroRNA-337 by targeting Nrf-2. Biomed. Pharmacother. 2018, 107, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Chen, Q.; Chen, X.; Qi, H.; Yang, Y.; Li, W.; Yang, X.; Gunawan, A.; Chen, S.; Zhang, H.; et al. Morusin and mulberrin extend the lifespans of yeast and C. elegans via suppressing nutrient-sensing pathways. Geroscience 2023, 45, 949–964. [Google Scholar] [CrossRef]
- Lv, Q.; Lin, J.; Wu, X.; Pu, H.; Guan, Y.; Xiao, P.; He, C.; Jiang, B. Novel active compounds and the anti-diabetic mechanism of mulberry leaves. Front. Pharmacol. 2022, 13, 986931. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.H.; Yuan, H.H.; Xu, M.T.; Hong, Y.T.; Gao, C.C.; Wu, Z.P.; Han, H.T.; Sun, X.; Gao, R.L.; Yang, S.F.; et al. A novel Diels-Alder adduct of mulberry leaves exerts anticancer effect through autophagy-mediated cell death. Acta. Pharmacol. Sin. 2021, 42, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Gor, R.; Saha, L.; Agarwal, S.; Karri, U.; Sohani, A.; Madhavan, T.; Pachaiappan, R.; Ramalingam, S. Morin inhibits colon cancer stem cells by inhibiting PUM1 expression in vitro. Med. Oncol. 2022, 39, 251. [Google Scholar] [CrossRef]
- Chang, H.; Hou, J.; Shao, Y.; Xu, M.; Weng, X.; Du, Y.; Shi, J.; Zhang, L.; Cui, H. Sanggenon C inhibits cell proliferation and induces apoptosis by regulating the MIB1/DAPK1 axis in glioblastoma. MedComm 2023, 4, e281. [Google Scholar] [CrossRef]
- Chen, L.D.; Liu, Z.H.; Zhang, L.F.; Yao, J.N.; Wang, C.F. Sanggenon C induces apoptosis of colon cancer cells via inhibition of NO production, iNOS expression and ROS activation of the mitochondrial pathway. Oncol. Rep. 2017, 38, 2123–2131. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Luo, J.; Luo, X.; Jia, W.; Fang, Z.; Yi, S.; Li, L. Morusin exerts anti-cancer activity in renal cell carcinoma by disturbing MAPK signaling pathways. Ann. Transl. Med. 2020, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.L.; Lv, Y.; Wang, S.; Zhang, Y.Q. Morusin suppresses A549 cell migration and induces cell apoptosis by downregulating the expression of COX-2 and VEGF genes. Oncol. Rep. 2018, 40, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, X.; Ye, M. Morusin inhibits the growth of human colorectal cancer HCT116-derived sphere-forming cells via the inactivation of Akt pathway. Int. J. Mol. Med. 2021, 47, 1. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.H.; Tsai, Y.T.; Yen, M.H.; Lin, C.C.; Liang, C.J.; Yang, T.H.; Lee, C.W.; Yen, F.L. Norartocarpetin from a folk medicine Artocarpus communis plays a melanogenesis inhibitor without cytotoxicity in B16F10 cell and skin irritation in mice. BMC Complement. Altern. Med. 2013, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Xiao, Y.; Chen, D.; Wang, J. Antiproliferative effects of Norartocarpetin isoflavone in human lung carcinoma cells are mediated via targeting Ras/Raf/MAPK signalling pathway, mitochondrial mediated apoptosis, S-phase cell cycle arrest and suppression of cell migration and invasion. JBUON 2020, 25, 855–861. [Google Scholar] [PubMed]
- Maharjan, S.; Lee, M.G.; Kim, S.Y.; Lee, K.S.; Nam, K.S. Morin Sensitizes MDA-MB-231 Triple-Negative Breast Cancer Cells to Doxorubicin Cytotoxicity by Suppressing FOXM1 and Attenuating EGFR/STAT3 Signaling Pathways. Pharmaceuticals-Base 2023, 16, 672. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kwon, E.B.; Kim, B.; Chung, H.S.; Choi, G.; Kim, Y.H.; Choi, J.G. Mulberry Component Kuwanon C Exerts Potent Therapeutic Efficacy In Vitro against COVID-19 by Blocking the SARS-CoV-2 Spike S1 RBD:ACE2 Receptor Interaction. Int. J. Mol. Sci. 2022, 23, 12156. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, H.; Lin, L.; Gao, Y.; Niu, X. Mulberrin inhibits Botrytis cinerea for strawberry storage by interfering with the bioactivity of 14α-demethylase (CYP51). Food. Funct. 2022, 13, 4032–4046. [Google Scholar] [CrossRef]
- Hu, Y.C.; Ji, D.W.; Zhao, C.Y.; Zheng, H.; Chen, Q.A. Catalytic Prenylation and Reverse Prenylation of Indoles with Isoprene: Regioselectivity Manipulation through Choice of Metal Hydride. Angew. Chem. Int. Ed. Engl. 2019, 58, 5438–5442. [Google Scholar] [CrossRef]
- Hu, Y.C.; Min, X.T.; Ji, D.W.; Chen, Q.A. Catalytic prenylation and reverse prenylation of aromatics. Trends Chem. 2022, 4, 658–675. [Google Scholar] [CrossRef]
- Wu, S.C.; Han, F.; Song, M.R.; Chen, S.; Li, Q.; Zhang, Q.; Zhu, K.; Shen, J.Z. Natural Flavones from Morus alba against Methicillin-Resistant Staphylococcus aureus via Targeting the Proton Motive Force and Membrane Permeability. J. Agric. Food. Chem. 2019, 67, 10222–10234. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; So, I.; Chun, J.N.; Jeon, J.H. The antitumor effects of geraniol: Modulation of cancer hallmark pathways (Review). Int. J. Oncol. 2016, 48, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Botta, B.; Menendez, P.; Zappia, G.; de Lima, R.A.; Torge, R.; Monachea, G.D. Prenylated isoflavonoids: Botanical distribution, structures, biological activities and biotechnological studies. An update (1995–2006). Curr. Med. Chem. 2009, 16, 3414–3468. [Google Scholar] [CrossRef]
- Gao, Y.Q.; Yang, W.; Karplus, M. A structure-based model for the synthesis and hydrolysis of ATP by F1-ATPase. Cell 2005, 123, 195–205. [Google Scholar] [CrossRef]
- Guo, H.; Courbon, G.M.; Bueler, S.A.; Mai, J.; Liu, J.; Rubinstein, J.L. Structure of mycobacterial ATP synthase bound to the tuberculosis drug bedaquiline. Nature 2021, 589, 143–147. [Google Scholar] [CrossRef]
- Guo, X.; Yang, N.; Ji, W.; Zhang, H.; Dong, X.; Zhou, Z.; Li, L.; Shen, H.M.; Yao, S.Q.; Huang, W. Mito-Bomb: Targeting Mitochondria for Cancer Therapy. Adv. Mater. 2021, 33, e2007778. [Google Scholar] [CrossRef]
- Yang, Y.; Karakhanova, S.; Hartwig, W.; D’Haese, J.G.; Philippov, P.P.; Werner, J.; Bazhin, A.V. Mitochondria and Mitochondrial ROS in Cancer: Novel Targets for Anticancer Therapy. J. Cell. Physiol. 2016, 231, 2570–2581. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gao, Z.; Liu, X.; Agarwal, P.; Zhao, S.; Conroy, D.W.; Ji, G.; Yu, J.; Jaroniec, C.P.; Liu, Z.; et al. Targeted production of reactive oxygen species in mitochondria to overcome cancer drug resistance. Nat. Commun. 2018, 9, 562. [Google Scholar] [CrossRef]
- Zou, Z.; Chang, H.; Li, H.; Wang, S. Induction of reactive oxygen species: An emerging approach for cancer therapy. Apoptosis 2017, 22, 1321–1335. [Google Scholar] [CrossRef]
- Giorgio, V.; Burchell, V.; Schiavone, M.; Bassot, C.; Minervini, G.; Petronilli, V.; Argenton, F.; Forte, M.; Tosatto, S.; Lippe, G.; et al. Ca2+ binding to F-ATP synthase β subunit triggers the mitochondrial permeability transition. EMBO Rep. 2017, 18, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Takada, K. Reactive oxygen species in cancer: Current findings and future directions. Cancer Sci. 2021, 112, 3945–3952. [Google Scholar] [CrossRef] [PubMed]
- Juarez, D.; Fruman, D.A. Targeting the Mevalonate Pathway in Cancer. Trends Cancer 2021, 7, 525–540. [Google Scholar] [CrossRef] [PubMed]
- Bloor, S.; Michurin, I.; Titchiner, G.R.; Leys, D. Prenylated flavins: Structures and mechanisms. FEBS J. 2023, 290, 2232–2245. [Google Scholar] [CrossRef] [PubMed]
- Panek-Krzyśko, A.; Stompor-Gorący, M. The Pro-Health Benefits of Morusin Administration—An Update Review. Nutrients 2021, 13, 3043. [Google Scholar] [CrossRef] [PubMed]
- Škovranová, G.; Čulenová, M.; Treml, J.; Dzurická, L.; Marova, I.; Sychrová, A. Prenylated phenolics from Morus alba against MRSA infections as a strategy for wound healing. Front. Pharmacol. 2022, 13, 1068371. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, G.; Ma, B.; Zhong, C.; He, N. Metabolic Profiling and Transcriptome Analysis of Mulberry Leaves Provide Insights into Flavonoid Biosynthesis. J. Agric. Food Chem. 2020, 68, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.-L.; Zhao, M.; Xu, J.-Y.; Lv, T.-M.; Liu, X.-C.; Sun, S.; Guan, Q.; Zhou, Z.-C.; Wu, J.; Zhao, M.-Y.; et al. Inhibitory Mechanism of Prenylated Flavonoids Isolated from Mulberry Leaves on α-Glucosidase by Multi-Spectroscopy and Molecular Dynamics Simulation. J. Agric. Food Chem. 2023, 71, 9135–9147. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Liao, L.; Yu, P.; Long, B.; Che, Y.; Lu, L.; Xu, B. Total syntheses and antibacterial evaluations of cudraflavones A-C and related Flavones. Bioorg. Chem. 2023, 140, 106764. [Google Scholar] [CrossRef]
- Choumessi, A.T.; Johanns, M.; Beaufay, C.; Herent, M.F.; Stroobant, V.; Vertommen, D.; Corbet, C.; Jacobs, R.; Herinckx, G.; Steinberg, G.R.; et al. Two isoprenylated flavonoids from Dorstenia psilurus activate AMPK, stimulate glucose uptake, inhibit glucose production and lower glycemia. Biochem. J. 2019, 476, 3687–3704. [Google Scholar] [CrossRef]
- Chang, Y.; Hung, C.F.; Ko, H.H.; Wang, S.J. Albanin A, Derived from the Root Bark of Morus alba L., Depresses Glutamate Release in the Rat Cerebrocortical Nerve Terminals via Ca2+/Calmodulin/Adenylate Cyclase 1 Suppression. J. Med. Food. 2021, 24, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Yen, F.L.; Ko, H.H.; Li, S.Y.; Chiang, Y.C.; Lee, M.H.; Tsai, M.H.; Hsu, L.F. Cudraflavone C Induces Apoptosis of A375.S2 Melanoma Cells through Mitochondrial ROS Production and MAPK Activation. Int. J. Mol. Sci. 2017, 18, 1508. [Google Scholar] [CrossRef]
- Tang, S.; Huang, W.; Ji, S.; Wang, Y.; Pei, D.; Ye, M.; Yu, S. Prenylated flavonoids from Glycyrrhiza uralensis as promising anti-cancer agents: A preliminary structure-activity study. J. Chin. Pharm. Sci. 2016, 25, 23–29. [Google Scholar]
- Aelenei, P.; Rimbu, C.M.; Horhogea, C.E.; Lobiuc, A.; Neagu, A.N.; Dunca, S.I.; Motrescu, I.; Dimitriu, G.; Aprotosoaie, A.C.; Miron, A. Prenylated phenolics as promising candidates for combination antibacterial therapy: Morusin and kuwanon G. Saudi. Pharm. J. 2020, 28, 1172–1181. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wu, Z.; He, Y.; Xiao, Y.; Xia, C. Single and dual target inhibitors based on Bcl-2: Promising anti-tumor agents for cancer therapy. Eur. J. Med. Chem. 2020, 201, 112446. [Google Scholar] [CrossRef]
- Roy, R.; Ria, T.; RoyMahaPatra, D.; Sk, U.H. Single Inhibitors versus Dual Inhibitors: Role of HDAC in Cancer. ACS Omega 2023, 8, 16532–16544. [Google Scholar] [CrossRef] [PubMed]
- Petrelli, A.; Giordano, S. From single- to multi-target drugs in cancer therapy: When aspecificity becomes an advantage. Curr. Med. Chem. 2008, 15, 422–432. [Google Scholar]
- Jaaks, P.; Coker, E.A.; Vis, D.J.; Edwards, O.; Carpenter, E.F.; Leto, S.M.; Dwane, L.; Sassi, F.; Lightfoot, H.; Barthorpe, S.; et al. Effective drug combinations in breast, colon and pancreatic cancer cells. Nature 2022, 603, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Holton, R.A.; Kim, H.B.; Somoza, C.; Liang, F.; Biediger, R.J.; Boatman, P.D.; Shindo, M.; Smith, C.C.; Kim, S.C.; Nadizadeh, H.; et al. First total synthesis of taxol. 2. Completion of the C-ring and D-ring. J. Am. Chem. Soc. 1994, 116, 1599–1600. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Yang, Z.; Liu, J.J.; Ueno, H.; Nantermet, P.G.; Guy, R.K.; Claiborne, C.F.; Renaud, J.; Couladouros, E.A.; Paulvannan, K.; et al. Total synthesis of taxol. Nature 1994, 367, 630–634. [Google Scholar] [CrossRef]
- Paterson, I.; Naylor, G.J.; Fujita, T.; Guzmán, E.; Wright, A.E. Total synthesis of a library of designed hybrids of the microtubule-stabilising anticancer agents taxol, discodermolide and dictyostatin. Chem. Commun. 2010, 46, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, S.; Croteau, R. Taxol: Biosynthesis, molecular genetics, and biotechnological applications. Appl. Microbiol. Biotechnol. 2001, 57, 13–19. [Google Scholar] [PubMed]
- He, N.; Zhang, C.; Qi, X.; Zhao, S.; Tao, Y.; Yang, G.; Lee, T.H.; Wang, X.; Cai, Q.; Li, D.; et al. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 2445. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.X.; Cao, Y. CB-Dock2: Improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic. Acids. Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, G.; Qian, P.; Chen, L.; He, N. Kuwanon C Inhibits Tumor Cell Proliferation and Induces Apoptosis by Targeting Mitochondria and Endoplasmic Reticulum. Int. J. Mol. Sci. 2024, 25, 8293. https://doi.org/10.3390/ijms25158293
Yuan G, Qian P, Chen L, He N. Kuwanon C Inhibits Tumor Cell Proliferation and Induces Apoptosis by Targeting Mitochondria and Endoplasmic Reticulum. International Journal of Molecular Sciences. 2024; 25(15):8293. https://doi.org/10.3390/ijms25158293
Chicago/Turabian StyleYuan, Gangxiang, Peng Qian, Lin Chen, and Ningjia He. 2024. "Kuwanon C Inhibits Tumor Cell Proliferation and Induces Apoptosis by Targeting Mitochondria and Endoplasmic Reticulum" International Journal of Molecular Sciences 25, no. 15: 8293. https://doi.org/10.3390/ijms25158293