Abstract
Blackleg disease, caused by Leptosphaeria spp. fungi, is one of the most important diseases of Brassica napus, responsible for severe yield losses worldwide. Blackleg resistance is controlled by major R genes and minor quantitative trait loci (QTL). Due to the high adaptation ability of the pathogen, R-mediated resistance can be easily broken, while the resistance mediated via QTL is believed to be more durable. Thus, the identification of novel molecular markers linked to blackleg resistance for B. napus breeding programs is essential. In this study, 183 doubled haploid (DH) rapeseed lines were assessed in field conditions for resistance to Leptosphaeria spp. Subsequently, DArTseq-based Genome-Wide Association Study (GWAS) was performed to identify molecular markers linked to blackleg resistance. A total of 133,764 markers (96,121 SilicoDArT and 37,643 SNP) were obtained. Finally, nine SilicoDArT and six SNP molecular markers were associated with plant resistance to Leptosphaeria spp. at the highest significance level, p < 0.001. Importantly, eleven of these fifteen markers were found within ten genes located on chromosomes A06, A07, A08, C02, C03, C06 and C08. Given the immune-related functions of the orthologues of these genes in Arabidopsis thaliana, the identified markers hold great promise for application in rapeseed breeding programs.
1. Introduction
Brassica napus (genome AACC, 2n = 38) is one of the most widespread oilseed crops in the world [1], known as rapeseed or canola. Rapeseed is an amphidiploid species that originated from two closely related species: Brassica rapa (AA, 2n = 20) and Brassica oleracea (CC, 2n = 18) as a result of interspecific hybridization and chromosome doubling. Its formation is dated to about 7500 years ago [2]. Until the 1970s, rapeseed oil was not considered suitable for consumption, due to its less than favorable taste and high erucic acid (EA) and glucosinolates (GSLs) content [3]. EA is considered to be toxic to humans and animals. Several studies performed on rats stated the presence of cardiotoxic properties due to the incomplete metabolism of EA in the body, resulting in the accumulation of triacylglycerols [4]. Furthermore, GSLs are known for their antimicrobial and anticarcinogenic properties; however, they can turn into anti-nutritious agents. Moreover, the bitter taste of some of the GSLs affect the palatability of the seed oil [5]. Rapeseed oil became suitable for consumption with the deployment of “double-zero” or “00” cultivars with the content of EA <2% and the meal having less than <25 mmol/kg at a moisture content of 9% of glucosinolates [6,7]. Currently, the term “canola” refers to edible cultivars of rapeseed with a lowered content of erucic acid and glucosinolates (“00”) and is a trademark for the Canadian Canola Association [8]. Rapeseed is one of the major players in the global oilseed market, ranking third in terms of oil production, right behind soybean and palm. In Poland, its production area reached 1.1 Mha in 2023 with a yield of 3.32 t/ha (stat.gov.pl). Rapeseed is a relatively demanding crop, as its yield depends on both biotic and abiotic factors such as soil quality, environmental conditions and the occurrence of diseases [8,9]. Regrettably, the reliance on monoculture cultivation for rapeseed creates favorable conditions for the uncontrolled spread of pathogens.
Blackleg is one of the most economically important diseases affecting Brassica crops. Its casual agents are Leptosphaeria maculans and Leptosphaeria biglobosa (new classification: Plenodomus lingam and Plenodomus biglobosus, respectively). This disease causes significant losses worldwide, exceeding 10% annually. In some cases, reports indicate losses as high as 50% [10,11]. Yearly loss in Poland reaches 10–40%, depending on the region [12]. Thus, L. maculans is a major threat for global rapeseed production. It is a hemibiotrophic fungi that can infect the host in every stage of plant development. However, the most severe plant damage is observed between the germination of seeds and the sixth leaf growth stage [13]. The initial symptom of blackleg is the appearance of gray lesions scattered on cotyledons and lower leaves with black pycnidia. As the infection progresses, basal stem cankers emerge and the lesions spread to the upper leaves, stems and siliques [7]. The high adaptability of the pathogen poses another challenge to rapeseed production. Its ability to reproduce both sexually (ascospores) and asexually (pycnidiospores) is the primary driving force of the pathogen evolution. Furthermore, L. maculans can survive in dry and cold environments as a saprophyte in crop residues. The propagation of the fungi occurs over both short and long distances through wind dispersal [14,15].
Several approaches to blackleg disease control have been utilized. Soil tillage, stubble management, crop rotation, the adjustment of sowing date, the use of microbiological control agents and fungicide application are only a few examples of methods used to limit L. maculans infections [16,17,18]. However, one of the most sustainable strategies is the use of resistant cultivars of canola. Several rapeseed varieties with medium to high blackleg tolerance have been released and commercialized. They utilize both race-specific, major gene (R gene)-mediated resistance and non-race-specific, polygenic resistance (quantitative trait loci, QTL). To date, at least 20 blackleg R genes have been identified, but only five of them have been cloned: Rlm2, Rlm4, Rlm7, Rlm9 and LepR3 [19,20,21,22]. Single or pyramided R genes are commonly used in commercial cultivars of rapeseed, such as Aganos (Rlm7, Syngenta) and LG Scorpion (Rlm7, Limagrain), as well as Surpass 400 (Rlm1, RlmS), Jet Neuf (Rlm4) and Skipton (Rlm4, Rlm9), which are used as differential lines in host–pathogen interaction research [23,24]. However, their effectiveness against pathogens can diminish over time due to the evolution of new pathogen strains. Polygenic resistance is considered to be more durable and, when combined with race-specific resistance, may provide long-term protection [25,26,27].
The development of new resistant canola cultivars through traditional methods that rely mainly on the selection of genotypes with a favorable phenotype is time- and work-consuming. Currently, Marker Assisted Selection (MAS) is becoming one of the main methods of selection for resistant germplasm. Thus, its practical usefulness in resistance breeding is undeniable. However, due to the large complexity of the B. napus genome, identifying the exact location of resistance genes is challenging. This hinders the establishment of linked molecular markers [28,29]. Fortunately, with the increasing availability of many publicly accessible rapeseed reference genomes and gene assemblies, MAS becomes easier and is now one of the main research trends in B. napus breeding. Additionally, most studies focusing on the identification of novel molecular markers employ the use of doubled haploid (DH) lines–fully homozygous breeding lines. Their greatest advantage is the reduced breeding time in comparison to conventional methods of backcrossing and selfing. Rapeseed DH production is an extensively investigated topic [30]. Closely linked molecular markers can support and accelerate the development of new rapeseed varieties with high resistance levels. Combined with DH lines, the breeding time can be considerably shortened in both the marker development phase as well as in the selection of homozygous genotypes.
Diversity Arrays Technology sequencing (DArTseqTM) is a method that provides high-throughput data in a relatively short timeframe. It utilizes a reduction in genome complexity and Next-Generation Sequencing (NGS) to generate large numbers of two types of molecular markers: SNP (Single Nucleotide Polymorphism) markers and SilicoDArT markers. It has been successfully applied in genetic studies of numerous crops and genotypes [31,32,33,34].
This study aims to identify new molecular markers linked to blackleg resistance in rapeseed DH lines using the means of NGS and physical mapping.
2. Results
2.1. Phenotyping
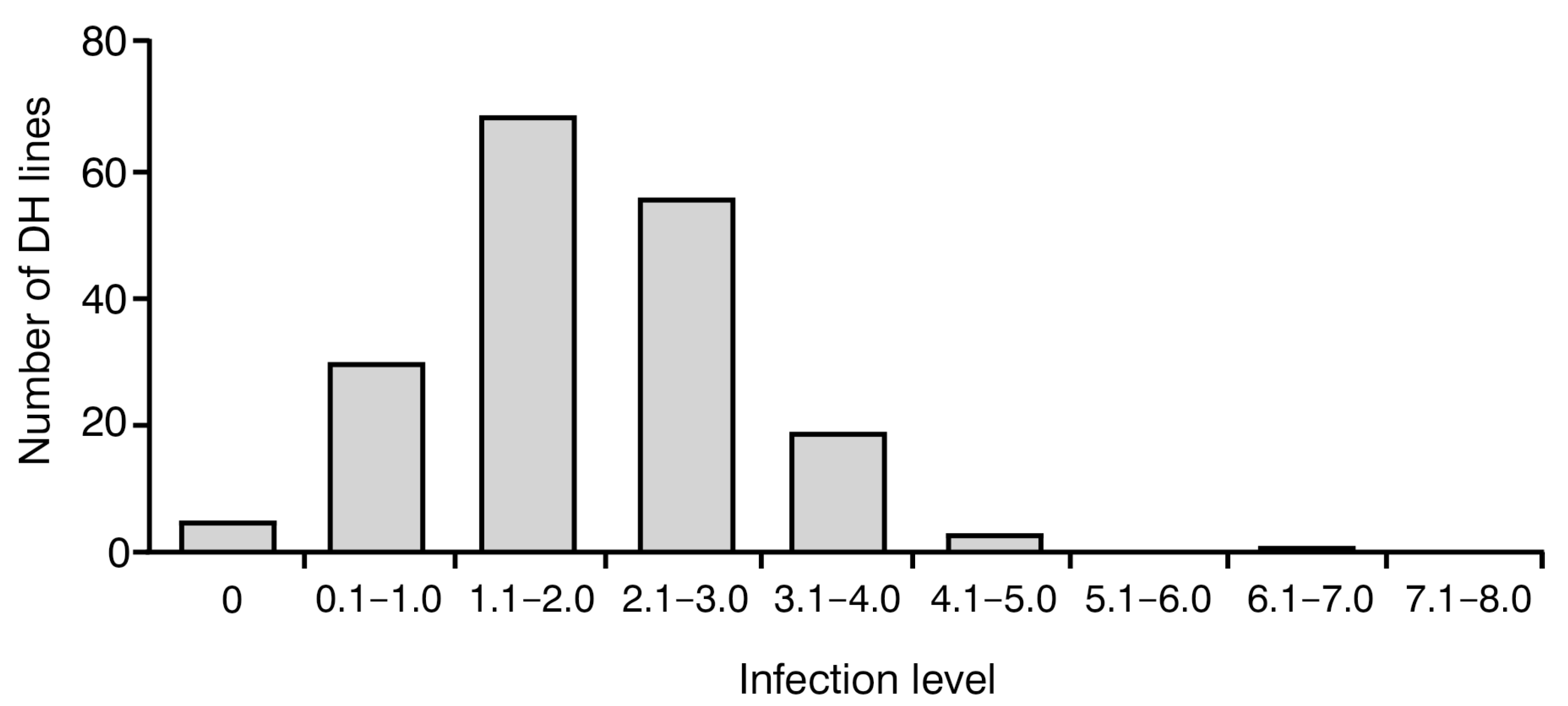
To investigate the level of resistance to blackleg in analyzed 183 DH lines, the frequency of disease symptoms was assessed using a 0–9 severity scale. On this scale, 0 indicates no visible disease damage and 9 indicates a completely damaged plant. The analysis of variance showed a significant effect (p < 0.001) in their genotype on the observed degree of resistance to blackleg. Notably, all plants were infected with Leptosphaeria spp. in varying degrees, ranging between 0 and 7, with an average value of 2.05 ± 1.06. Most of them (179) displayed satisfactory blackleg disease resistance with a score of 0–4. No plants with infection levels of 8–9 were observed (Figure 1).
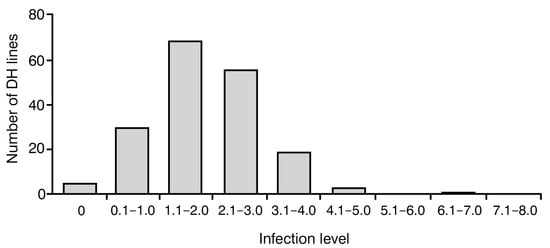
Figure 1.
Frequency distribution of blackleg infection levels in 183 analyzed Brassica napus doubled haploid (DH) lines. Score 0 indicates no visible damage; score 9 indicates completely damaged plant.
2.2. Genotyping
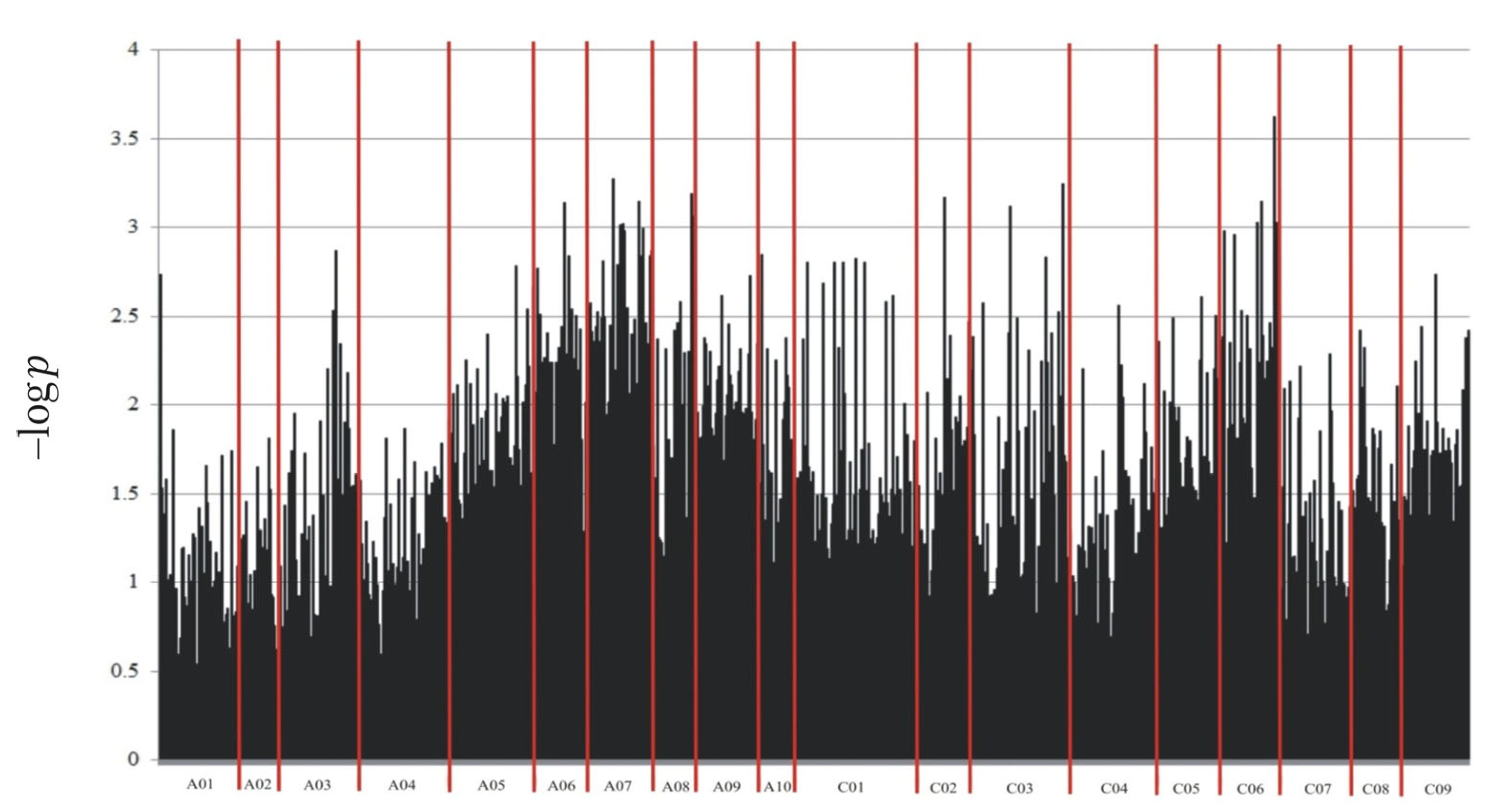
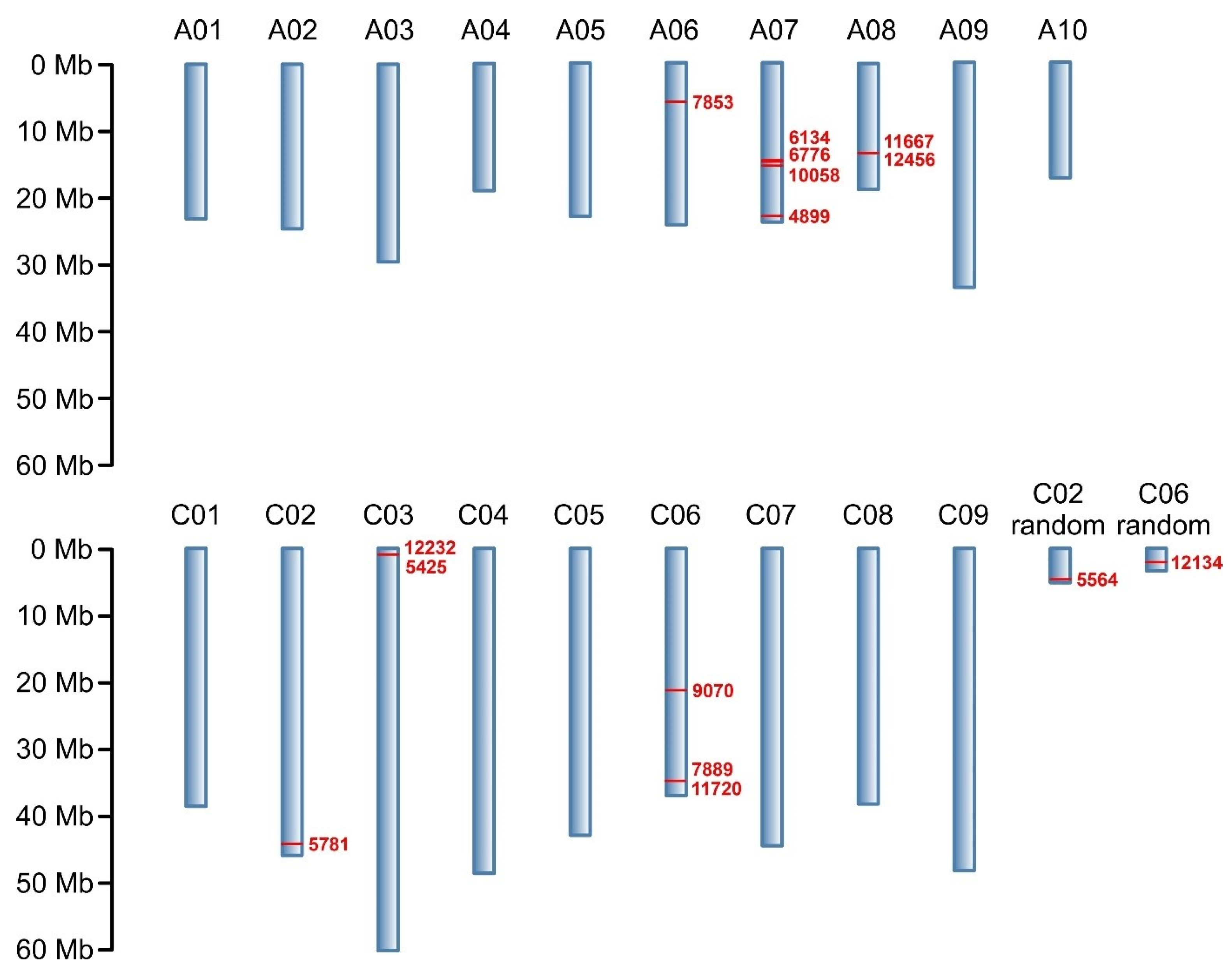
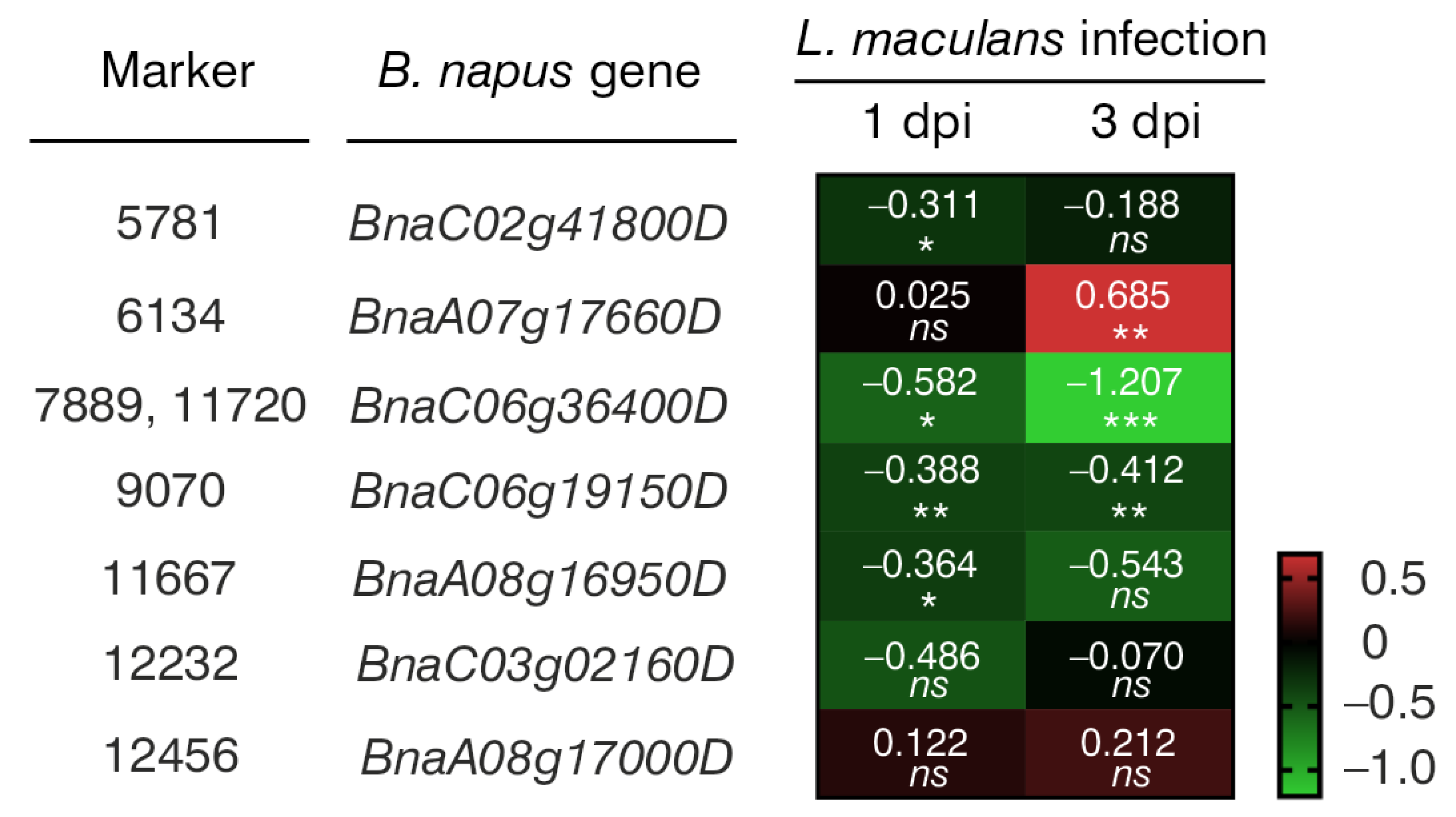
Through NGS, a total of 133,764 molecular markers (96,121 SilicoDArT and 37,643 SNP) were obtained. The significance of the identified markers was determined using MAF >0.25 and the number of missing observations <10%. Following this criteria, 12,948 (10,008 SilicoDArTs and 2940 SNPs) markers were selected for association mapping using GWAS (Figure 2). Importantly, fifteen (nine SilicoDArT and six SNP) markers from the latter group were associated with plant resistance to Leptosphaeria spp. at the highest significance level, p < 0.001 (Table 1). The highest number of them (4 and 3, respectively) was present on A07 and C06 chromosomes (Figure 3). Further analyses also revealed that eleven of these fifteen markers were located within only ten genes, due to the presence of the 7889SilicoDArT and 11720SNP marker in the same gene, BnaC06g36400D (Table 2). The potential role of these genes as candidate genes for rapeseed defense against L. maculans was assessed with a use of transcriptomic data from Becker et al. (2019) [34]. Out of seven genes found in the latter work, five exhibited significant changes in expression within three days after the pathogen inoculation, namely BnaC02g41800D, BnaA07g17660D, BnaC06g36400D, BnaC06g19150D and BnaA08g16950D (Figure 4).
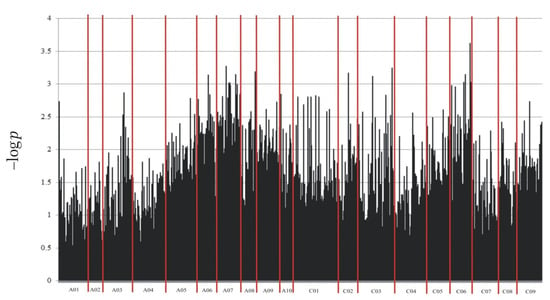
Figure 2.
Manhattan plot for polymorphic markers associated with resistance of B. napus to blackleg.

Table 1.
SilicoDArT and SNP molecular markers significantly associated with resistance of B. napus to blackleg at p < 0.001.
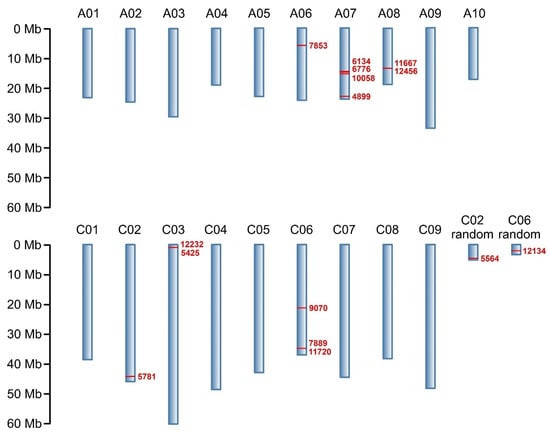
Figure 3.
Physical map of markers associated with resistance of B. napus to blackleg at p < 0.001. Chromosomes are shown in blue; chromosome numbers are annotated above. Reported markers and their positions on chromosomes are shown in red. Marker locations are expressed in megabases (Mb) as indicated by the scale.
Table 2.
Candidate genes linked to the markers identified at p < 0.001.
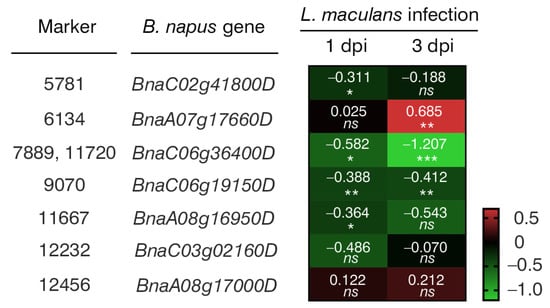
Figure 4.
A heatmap showing changes in the expression levels of seven B. napus genes that harbor selected markers caused by Leptosphaeria maculans 1 and 3 days post-inoculation (dpi). The heatmap depicts log2-fold changes in transcriptomic data acquired from Becker et al. (2019). Intensity of red indicates upregulated gene expression, while the intensity of green indicates downregulated gene expression. Significant differences from the mock-treated plants were determined using two-tailed Student’s t-test. * p < 0.05; ** p < 0.01; *** p < 0.001; ns, not significant.
3. Materials and Methods
3.1. Plant Material
The plant material consisted of 183 DH lines of rapeseed (B. napus L.) derived from Strzelce Plant Breeding Ltd. (Strzelce, Poland) IHAR Group in Borowo. The DH lines were obtained by performing intraspecific crosses of selected registered rapeseed varieties with known blackleg resistance in order to represent a diverse disease response structure in the mapping population.
3.2. Field Assessment
All DH lines were assessed in terms of the resistance to blackleg in field conditions. The trials were conducted on Strzelce Plant Breeding Ltd. IHAR Group Borowo test fields. The experiment was set up in a completely randomized block design and included three replications. The observations were completed on adult plants in the BBCH 70–89 phase. The level of blackleg resistance was determined using disease symptoms evaluation according to the 0–9 scale elaborated by Jedryczka (2006) [35], in which score 0 means no visible disease symptoms and score 9 means a completely damaged plant with numerous pycnidia on the stem and leaf surfaces. The details of the evaluation scale were as follows:
- 0
- healthy plant with no visible disease symptoms;
- 1
- slight surface discoloration of the main stem;
- 2
- discoloration covering up to 10% of the main stem;
- 3
- discoloration covering from 11% to 20% of the main stem;
- 4
- discoloration covering from 21% to 30% of the main stem, a few pycnidia;
- 5
- discoloration covering from 31% to 50% of the main stem, several pycnidia;
- 6
- discoloration covering from 51% to 75% of the main stem, numerous pycnidia;
- 7
- discoloration covering over 76% of the main stem and parts of side branches, numerous pycnidia;
- 8
- discoloration covering the whole main stem and extending to side branches, numerous pycnidia, smaller number of siliques as compared to the healthy plants;
- 9
- dead plant with thin stem, very small number or no siliques, numerous pycnidia on the main stem and side branches.
Like in all visual assessments, this scale was an indication for the evaluator, but variations were possible.
3.3. DNA Extraction
Whole genomic DNA was extracted from young seedlings using a Genomic Mini AX Plant kit (A&A Biotechnology, Gdańsk, Poland) according to supplied protocol. The concentration and purity of the isolated DNA was determined using the spectrophotometer DeNovix DS-11, DeNovix, Wilmington, DE, USA. Samples with a DNA concentration higher than 100 µL and 260/280, 260/230 ratios of ~1.8 and ~2.0, respectively, were diluted with Tris-HCl buffer to 100 ng/µL in 25 µL volume.
3.4. Genotyping
The genomic DNA isolated from 183 DH lines, previously assessed in field conditions, was sent to Diversity Arrays Technology (University of Canberra, Australia) for DArTseq analysis. In the latter procedure, whole genomic material was digested by selected restriction enzymes to reduce the complexity of the genome [36]. This approach allowed the fragmentation of DNA samples in a highly reproducible manner and the consequent selection of the predominantly active, low-copy sequence areas, which are the most informative for genetic analysis. Next, the digested fragments underwent ligation with adaptors and amplification using Polymerase Chain Reaction (PCR). Equimolar quantities of amplification products from each genotype were combined to create gene pool representation, followed by sequencing with Illumina Hiseq2000. Generated reads were processed with DArT analytical pipelines (Diversity Arrays Technology, Australia) to filter poor quality sequences and to identify sequences for marker calling. Only markers meeting the following criteria were selected for the association analysis: one marker within a given sequence (69 nt), an allele frequency (MAF) >0.25 and a missing observations fraction <10%.
The physical map positions of identified markers were determined against the Darmor-bzh v4.1 reference genome. Obtained metadata (i.e., presence/absence and localization of identified SilicoDArTs and SNP markers) were further subjected to statistical analysis.
The distances between the markers and the nearest genes were measured using the JBrowse tool against the Darmor-bzh 4.1 assembly, available through the B. napus multi-omics information resource (BnIR, https://yanglab.hzau.edu.cn/BnIR (accessed on 22 March 2024)) platform [37].
3.5. Statistical Analysis
Data were further subjected to analysis of variance in a model with random effects of genotypes. Association mapping with Genome-Wide Association Studies (GWAS), based on the SilicoDArT and SNP data and the average resistance to blackleg values, was conducted using the method based on the mixed linear model with a population structure estimated via eigenanalysis and modeled using random effects [38,39]. All analyses and visualizations of the results were performed in GenStat 23.1 [40], using the QSASSOCIATION procedure. QSASSOCIATION performs a mixed model marker—trait association analysis (also known as linkage disequilibrium mapping) with data from a single-environment trial. To avoid false positives in association mapping studies, a control is necessary for genetic relatedness. The model used was specified using the RELATIONSHIPMODEL = eigenanalysis option, which infers the underlying genetic substructure in the population by retaining the most significant principal components from the molecular marker matrix [41]; the scores of the significant axes are used as covariables in the mixed model, which effectively is an approximation to the structuring of the genetic variance covariance matrix by a coefficient of the coancestry matrix (kinship matrix) [31]. The significance of association between the resistance to blackleg and SilicoDArT and SNP markers was assessed using p values corrected for multiple testing using the Benjamini–Hochberg method.
3.6. Functional Annotation of Genes
The functional annotation of the potential involvement of selected genes in B. napus defense mechanisms against L. maculans was determined based on transcriptomic data from Becker et al. (2019) [42], available on the BrassicaEDB platform (https://brassica.biodb.org/ (accessed on 25 March 2024)).
4. Discussion
The Leptosphaeria spp. fungal complex is a great threat to rapeseed crop cultivation worldwide [43], especially after warm and wet autumns [44]. The breakdown of resistance in commercial rapeseed cultivars emphasizes the importance of searching for novel resistance sources in rapeseed breeding. Molecular markers are becoming an essential tool for the selection of resistant plants. Currently, the trends in modern farming and Integrated Pest Management programs require innovative and environmentally friendly solutions, allowing effective protection against diseases and pests [45,46,47]. Plant resistance breeding combined with MAS is one of such solutions. Developing molecular markers linked with blackleg resistance is an extremely complex and tremendous task.
The recently observed reduction in the cost of sequencing platforms has increased the efficiency of GWAS. Moreover, the ability of obtaining a large number of markers in relatively short time makes GWAS an incredibly useful tool for association mapping and for uncovering the genetic architecture of agronomically important phenotype characteristics.
The GWAS technique was successfully used in numerous Brassica studies to discover genome regions associated with both quality and resistance traits. For instance, GWAS has been used in the identification of SNP markers connected to glucosinolate and hemicellulose seed content [48], the identification of flowering-related traits [49] and for associating SNPs with resistance to Plasmodiophora brassicae [50]. Furthermore, the combination of GWAS, QTL mapping and expression analysis led to the recognition of a candidate gene controlling the branch number in rapeseed [51]. For blackleg resistance, several markers connected to major R genes and QTLs in Brassicas were also discovered, e.g., candidate Rlm6 gene in B. juncea [52], QTL resistance loci in B. napus [53,54] and candidate LepR1 genes in B. napus [55]. The recent discoveries deepened the knowledge of the genetic architecture of the blackleg resistance and offered new perspectives on complex pathogen–host relationships.
As previously mentioned, the L. maculans–B. napus interaction employs two types of resistance. The first one, qualitative resistance mediated by major R genes, is relatively well recognized and studied. In this type, it is believed that the L. maculans–B. napus pathosystem follows Flor’s gene-for-gene theory [56]. Thus, the successful deployment of resistant cultivars and the control of blackleg disease relies on the recognition of host and pathogen genotype. The characterization of single R genes is possible through phenotypic tests that use inoculation [57]. However, such tests are often very laborious, time consuming and require an extensive collection of precisely described pathogen isolates. The use of molecular markers enables the exact characterization of deployed varieties and the establishment of a rotation strategy in a shorter and more convenient way [58]. On the other hand, the second type of resistance, quantitative resistance (QTL), is poorly understood. It is mediated by several minor genes with small effect. Additionally, QTLs have low heritability and a non-Mendelian and accumulative character. It is challenging to pinpoint the exact genes or molecular markers linked with qualitative blackleg resistance because of the strong genotype x environment effect. Moreover, the method of evaluation, localization and the selection of winter, semi-winter and spring type rapeseed may affect the outcome of the analysis [59]. To date, multiple QTLs have been located on all 19 chromosomes of B. napus, but few have proved to be effective in various environmental conditions [60,61,62,63].
In this work, we evaluated the resistance of 183 rapeseed DH lines to blackleg via the observation of disease symptoms in adult plants (Figure 1). The analysis of variance showed a significant effect in their genotype on the observed degree of resistance to blackleg.
Among 12,948 markers selected for association mapping, 15 markers located on the B. napus chromosomes A06, A07, A08, C02, C03, C06 and C08 were found to be linked to blackleg resistance at the highest significance level, p < 0.001. Their greatest concentration was observed on chromosome A07 (Figure 3). This finding aligns with previous estimations indicating that this chromosome harbors the most R genes, which are responsible for disease resistance in plants [64]. However, it is crucial to emphasize that the A07 chromosome is not the sole repository of R genes against fungal infections. Rather, this group is dispersed throughout the rapeseed genome, as evidenced by the presence of the selected markers on several other chromosomes.
Out of these 15 markers, 11 were found within 10 B. napus genes (BnaC02g41800D, BnaA07g17660D, BnaA07g18190D, BnaA06g11460D, BnaC06g36400D, BnaC06g19150D, BnaA07g19340D, BnaA08g16950D, BnaC03g02160D and BnaA08g17000D) (Table 2). While SilicoDArT marker sequences were more evenly distributed between introns and exons (three exonic, one intronic and two across both these gene elements), most of the identified SNPs (four of all five) were located in exons (Table 2). This discrepancy can be related to the more complex structure of SilicoDArT markers that often contain indels and SNPs, sometimes in combination, and span a larger genomic region than single SNPs (https://www.diversityarrays.com/ (accessed on 17 November 2023)). The SNP in marker 100058 was responsible for the occurrence of alanine (GCT in gene sequence) in relation to the valine (GTT) in the non-marker sequence that makes a missense mutation. The SNP in marker 12232 also contributes to a missense change encoding for serine (TCT) as an alternative to threonine (ACT). A change in SNP in marker 11720 to a non-marker sequence causes a nonsense mutation with a switch of cysteine-encoding TGT to a STOP codon. Thereby, these three SNPs can have a direct impact on the length and properties of the encoded proteins. There was also found a synonymic mutation where the ATC codon for isoleucine in the 11667 SNP marker is changed to another codon for this amino acid, ATT. In light of recent reports, despite the absence of an amino acid change in the protein, such a mutation may also not be neutral [65]. This would explain the observed linkage of the 11667 marker with rapeseed resistance to fungal pathogens.
Interestingly, the expression of half of these 10 genes showed statistically significant changes following L. maculans inoculation (Figure 4), suggesting their potential involvement in rapeseed’s defense against fungal pathogens. Similarly, according to the literature review, several of their orthologues in Arabidopsis thaliana were verified to take part in plant immune response to pathogen infection (Table 3). For example, A. thaliana At5g08130 (orthologue of BnaC02g41800D) encodes a basic helix–loop–helix transcription factor that is involved in brassinosteroid signaling upon stress conditions. Brassinosteroids appear to modulate plant interactions with pathogens. Their effect is dependent on plant and pathogen species [66]. In transgenic B. napus plants, the overexpression of the brassinosteroid biosynthetic gene AtDWF4 results in increased tolerance to environmental stresses, including a resistance to L. maculans and Sclerotinia sclerotiorum [67,68] (Table 3).
Table 3.
Annotation of Arabidopsis thaliana orthologous genes. Data obtained from NCBI (https://www.ncbi.nlm.nih.gov/ (accessed on 15 March 2024)), BrassicaDB (http://www.brassicadb.org (accessed on 15 March 2024)) and TAIR (https://www.arabidopsis.org/ (accessed on 15 March 2024)) databases.
At1g16980 (orthologue of BnaA06g11460D) encodes trehalose-6-phosphate synthase. The involvement of several trehalose-6-phosphate synthases and trehalose in increasing plant resistance to pathogens has been proved a number of times [69,70,71,72] (Table 3). Shi et al. (2019) suggested that trehalose may trigger signal transduction pathways [73].
At3g62600 (orthologue of BnaA07g19340D) encodes the DnaJ heat shock protein (HSP40) family member that has been extensively studied both in plants and animals. It takes part in protein folding and is involved in regulatory pathways of many physiological processes. The latter confer salt stress tolerance, drought tolerance, temperature stress and development [74,75,76,77] (Table 3). Liu et al. (2022) found that 16 out of 17 TaDnaJs were upregulated after wheat yellow mosaic virus infection, suggesting a role for DnaJs in plant defense responses [78]. It was also observed that the overexpression of the chloroplast-targeted DnaJ gene, LeCDJ2, confers a higher resistance to Pseudomonas solanacearum in tomato plants [79]. Moreover, Zhong et al. (2017) established that the silencing of OsDjA6 results in a higher resistance of rice to rice blast caused by Magnoporthe oryzae (Pyriculoria oryzae). It was concluded that this protein negatively regulates rice immunity to the pathogen [80].
At4g38690 (orthologue of BnaA08g16950D) encodes the phospholipase C-like (PLC) type of proteins that are commonly associated with plant immune pathways (Table 3). They are multidomain proteins with enzyme activity, involved in phosphoinositide metabolism and calcium signaling. Phosphoinositide-specific phospholipase C (PI-PLC) enzymes act downstream of the recognition of pathogens via immune receptors [81]. It was found that PLCs are crucial for hypersensitive (HR) immune response in plants. HR is attributed to resistance response in rapeseed mediated by major R genes [55]. The effectors of several plant pathogens such as Cladosporum fulvum, Pseudomonas syringae and Xanthomonas oryzae induced the accumulation of phosphatidic acid (PA). The activation of PI-PLC leads to the formation of PA as the final product [82,83]. PA is considered to be a messenger in the immune response pathway [84,85]. Due to high homogenicity of B. napus and A. thaliana genes, functional information can be considered as consolidated across the two species.
In addition to the chromosomes reported in this work, similar GWAS studies also found SNP and DArT markers related to blackleg resistance in other B. napus genomic localizations. The selected markers were linked to both R genes and QTLs [54,60,86,87]. Raman et al. (2016) identified 36 molecular markers located on 13 B. napus chromosomes. One major QTL observed on chromosome A01 explained as much as 14.7% of the phenotypic variation. Another study reported the identification of Rlm12 loci on chromosome A01 [86]. Most recent studies combine GWAS and Whole Genome Sequencing (WGS) methods in multi-trait, multi-year analyses. While most of localized regions are linked to known Rlm regions, tens of new potential resistance regions are detected. Several of these regions hold putative genes with functions related to stress response and disease defense [54,63,87,88].
In conclusion, this study employed a high-throughput approach to identify fifteen molecular markers (SilicoDArT and SNP type) associated with B. napus resistance to blackleg at the significance level of p < 0.001. Eleven of them were localized within ten genes. To evaluate the potential involvement of these genes in B. napus immune responses against fungal pathogens, a preliminary characterization was conducted based on either available B. napus transcriptomic data or known functions of A. thaliana orthologues described in the literature. All of them were found to take part in stress as well as immune response pathways to pathogen infection. This suggests that the identified molecular markers are indeed linked to blackleg resistance in rapeseed. They provide an invaluable insight into genetically mediated rapeseed resistance to blackleg and have the potential to become a powerful tool in resistance breeding through MAS. Additionally, this research also confirmed that DArTseq-based GWAS is an efficient method for marker selection and mapping.
Author Contributions
Conceptualization E.S., T.J., J.S. and J.N.; data curation: E.S., T.J. and J.B.; formal analysis J.B., E.S. and T.J.; investigation: E.S., J.S. and M.J.; methodology: E.S., J.S., J.B., M.J. and M.G., supervision J.N., visualization: J.B., T.J. and E.S., writing—original draft: E.S., T.J., J.S. and J.B.; writing—review and editing: J.S., M.J. and J.N. All authors have read and agreed to the published version of the manuscript.
Funding
The research presented in this publication was financed as part of the research project “Identification of molecular markers linked to genes conditioning resistance to blackleg (Leptosphaeria spp.), using advanced molecular techniques” (project number 27). The project is implemented with a grant from the Ministry of Agriculture and Rural Development, Poland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. Magdalena Grynia is an employee of the Strzelce Plant Breeding Company. Strzelce Plant Breeding Company had no role in the design, collection, analysis, or interpretation of the data, the writing of the manuscript, or the decision to publish the results.
References
- Friedt, W.; Tu, J.; Fu, T. Academic and Economic Importance of Brassica napus Rapeseed. In Compendium of Plant Genomes; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Zheng, X.; Koopmann, B.; Ulber, B.; von Tiedemann, A. A Global Survey on Diseases and Pests in Oilseed Rape—Current Challenges and Innovative Strategies of Control. Front. Agron. 2020, 2, 590908. [Google Scholar] [CrossRef]
- Abbadi, A.; Leckband, G. Rapeseed Breeding for Oil Content, Quality, and Sustainability. Eur. J. Lipid Sci. Technol. 2011, 113, 1198–1206. [Google Scholar] [CrossRef]
- Galanty, A.; Grudzińska, M.; Paździora, W.; Paśko, P. Erucic Acid—Both Sides of the Story: A Concise Review on Its Beneficial and Toxic Properties. Molecules 2023, 28, 1924. [Google Scholar] [CrossRef]
- Miklavčič Višnjevec, A.; Tamayo Tenorio, A.; Steenkjær Hastrup, A.C.; Hansen, N.M.L.; Peeters, K.; Schwarzkopf, M. Glucosinolates and Isothiocyantes in Processed Rapeseed Determined by Hplc-dad-qtof. Plants 2021, 10, 2548. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Erucic Acid in Feed and Food. EFSA J. 2016, 14, e04593. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of Rapeseed Powder from Brassica rapa L. and Brassica napus L. as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2020, 18, e06197. [Google Scholar] [CrossRef]
- Raymer, P.L. Canola: An Emerging Oilseed Crop. In Trends in New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 2002. [Google Scholar]
- Orlovius, K.; Kirkby, E.A. Fertilizing for High Yield and Quality Oilseed Rape. Int. Potash Inst. Bull. 2003, 16, 125. [Google Scholar]
- Van De Wouw, A.P.; Marcroft, S.J.; Howlett, B.J. Blackleg Disease of Canola in Australia. Crop Pasture Sci. 2016, 67, 273–283. [Google Scholar] [CrossRef]
- Hwang, S.F.; Strelkov, S.E.; Peng, G.; Ahmed, H.; Zhou, Q.; Turnbull, G. Blackleg (Leptosphaeria maculans) Severity and Yield Loss in Canola in Alberta, Canada. Plants 2016, 5, 31. [Google Scholar] [CrossRef]
- Gwiazdowski, R. Hamowanie Wzrostu Leptosphaeria maculans i Leptosphaeria biglobosa Przez Wybrane Fungicydy w Testach Płytkowych. Rośliny Oleiste-Oilseed Crops 2008, 19, 67–73. [Google Scholar]
- Marcroft, S.J.; Sosnowski, M.R.; Scott, E.S.; Ramsey, M.D.; Salisbury, P.A.; Howlett, B.J. Brassica napus Plants Infected by Leptosphaeria maculans after the Third to Fifth Leaf Growth Stage in South-Eastern Australia Do Not Develop Blackleg Stem Canker. Eur. J. Plant Pathol. 2005, 112, 289–292. [Google Scholar] [CrossRef]
- Savage, D.; Barbetti, M.J.; MacLeod, W.J.; Salam, M.U.; Renton, M. Temporal Patterns of Ascospore Release in Leptosphaeria maculans Vary Depending on Geographic Region and Time of Observation. Microb. Ecol. 2013, 65, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Travadon, R.; Sache, I.; Dutech, C.; Stachowiak, A.; Marquer, B.; Bousset, L. Absence of Isolation by Distance Patterns at the Regional Scale in the Fungal Plant Pathogen Leptosphaeria maculans. Fungal Biol. 2011, 115, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Aubertot, J.N.; Pinochet, X.; Doré, T. The Effects of Sowing Date and Nitrogen Availability during Vegetative Stages on Leptosphaeria maculans Development on Winter Oilseed Rape. Crop Prot. 2004, 23, 635–645. [Google Scholar] [CrossRef]
- Kutcher, H.R.; Fernando, W.G.D.; Turkington, T.K.; Mclaren, D.L. Best Management Practices for Blackleg Disease of Canola. Prairie Soils Crops J. 2011, 4, 122–134. [Google Scholar]
- Van de Wouw, A.P.; Marcroft, S.J.; Sprague, S.J.; Scanlan, J.L.; Vesk, P.A.; Idnurm, A. Epidemiology and Management of Blackleg of Canola in Response to Changing Farming Practices in Australia. Australas. Plant Pathol. 2021, 50, 137–149. [Google Scholar] [CrossRef]
- Haddadi, P.; Larkan, N.J.; Van deWouw, A.; Zhang, Y.; Xiang Neik, T.; Beynon, E.; Bayer, P.; Edwards, D.; Batley, J.; Borhan, M.H. Brassica napus Genes Rlm4 and Rlm7, Conferring Resistance to Leptosphaeria maculans, Are Alleles of the Rlm9 Wall-associated Kinase-like Resistance Locus. Plant Biotechnol. J. 2022, 20, 1229. [Google Scholar] [CrossRef] [PubMed]
- Larkan, N.J.; Ma, L.; Haddadi, P.; Buchwaldt, M.; Parkin, I.A.P.; Djavaheri, M.; Borhan, M.H. The Brassica napus Wall-Associated Kinase-like (WAKL) Gene Rlm9 Provides Race-Specific Blackleg Resistance. Plant J. 2020, 104, 892–900. [Google Scholar] [CrossRef]
- Larkan, N.J.; Lydiate, D.J.; Parkin, I.A.P.; Nelson, M.N.; Epp, D.J.; Cowling, W.A.; Rimmer, S.R.; Borhan, M.H. The Brassica Napus Blackleg Resistance Gene LepR3 Encodes a Receptor-like Protein Triggered by the Leptosphaeria maculans Effector AVRLM1. New Phytol. 2013, 197, 595–605. [Google Scholar] [CrossRef]
- Larkan, N.J.; Ma, L.; Borhan, M.H. The Brassica napus Receptor-like Protein RLM2 Is Encoded by a Second Allele of the LepR3/Rlm2 Blackleg Resistance Locus. Plant Biotechnol. J. 2015, 13, 983–992. [Google Scholar] [CrossRef]
- Balesdent, M.H.; Attard, A.; Kühn, M.L.; Rouxel, T. New avirulence genes in the phytopathogenic fungus Leptosphaeria maculans. Phytopathology 2002, 92, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Marcroft, S.J.; Elliott, V.L.; Cozijnsen, A.J.; Salisbury, P.A.; Howlett, B.J.; Van de Wouw, A.P. Identifying resistance genes to Leptosphaeria maculans in Australian Brassica napus cultivars based on reactions to isolates with known avirulence genotypes. Crop Pasture Sci. 2012, 63, 338–350. [Google Scholar] [CrossRef]
- Brun, H.; Chèvre, A.M.; Fitt, B.D.; Powers, S.; Besnard, A.L.; Ermel, M.; Huteau, V.; Marquer, B.; Eber, F.; Renard, M.; et al. Quantitative Resistance Increases the Durability of Qualitative Resistance to Leptosphaeria maculans in Brassica napus. New Phytol. 2010, 185, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Delourme, R.; Bousset, L.; Ermel, M.; Duffé, P.; Besnard, A.L.; Marquer, B.; Fudal, I.; Linglin, J.; Chadoeuf, J.; Brun, H. Quantitative Resistance Affects the Speed of Frequency Increase but Not the Diversity of the Virulence Alleles Overcoming a Major Resistance Gene to Leptosphaeria Maculans in Oilseed Rape. Infect. Genet. Evol. 2014, 27, 490–499. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Mitrousia, G.K.; Sidique, S.N.M.; Qi, A.; Fitt, B.D.L. Combining R Gene and Quantitative Resistance Increases Effectiveness of Cultivar Resistance against Leptosphaeria maculans in Brassica napus in Different Environments. PLoS ONE 2018, 13, e0197752. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.T.; Chawla, H.S.; Obermeier, C.; Dreyer, F.; Abbadi, A.; Snowdon, R. Chromosome-Scale Assembly of Winter Oilseed Rape Brassica napus. Front. Plant Sci. 2020, 11, 514081. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Jing, J.; Snowdon, R.J.; Mason, A.S.; Shen, J.; Meng, J.; Zou, J. Exploring the Gene Pool of Brassica napus by Genomics-Based Approaches. Plant Biotechnol. J. 2021, 19, 1693–1712. [Google Scholar] [CrossRef]
- Starosta, E.; Szwarc, J.; Niemann, J.; Szewczyk, K.; Weigt, D. Brassica napus Haploid and Double Haploid Production and Its Latest Applications. Curr. Issues Mol. Biol. 2023, 45, 4431–4450. [Google Scholar] [CrossRef] [PubMed]
- Bocianowski, J. Using NGS Technology and Association Mapping to Identify Candidate Genes Associated with Fusarium Stalk Rot Resistance. Genes 2024, 15, 106. [Google Scholar] [CrossRef]
- Alam, M.; Neal, J.; O’Connor, K.; Kilian, A.; Topp, B. Ultra-High-Throughput DArTseq-Based SilicoDArT and SNP Markers for Genomic Studies in Macadamia. PLoS ONE 2018, 13, e0203465. [Google Scholar] [CrossRef]
- Egea, L.A.; Mérida-García, R.; Kilian, A.; Hernandez, P.; Dorado, G. Assessment of Genetic Diversity and Structure of Large Garlic (Allium aativum) Germplasm Bank, by Diversity Arrays Technology “Genotyping-by-Sequencing” Platform (DArTseq). Front. Genet. 2017, 8, 272084. [Google Scholar] [CrossRef]
- Kilian, A.; Sanewski, G.; Ko, L. The Application of DArTseq Technology to Pineapple. Acta Hortic. 2016, 1111, 181–188. [Google Scholar] [CrossRef]
- Jedryczka, M. Epidemiology and Damage Caused by Stem Canker of Oilseed Rape in Poland. Ph.D. Thesis, Dissertations and Monographs, Abstract of Habilitation Thesis Institute of Plant Genetics, Polish Academy of Sciences, Poznań, Poland, 2006; pp. 42, 150, (In Polish, with English Abstract). [Google Scholar]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity Arrays Technology: A Generic Genome Profiling Technology on Open Platforms. Methods Mol. Biol. 2012, 888, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, S.; Wei, L.; Huang, Y.; Liu, D.; Jia, Y.; Luo, C.; Lin, Y.; Liang, C.; Hu, Y.; et al. BnIR: A Multi-Omics Database with Various Tools for Brassica napus Research and Breeding. Mol. Plant 2023, 16, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Malosetti, M.; Ribaut, J.M.; van Eeuwijk, F.A. The Statistical Analysis of Multi-Environment Data: Modeling Genotype-by-Environment Interaction and Its Genetic Basis. Front. Physiol. 2013, 4, 37433. [Google Scholar] [CrossRef] [PubMed]
- van Eeuwijk, F.A.; Bink, M.C.; Chenu, K.; Chapman, S.C. Detection and Use of QTL for Complex Traits in Multiple Environments. Curr. Opin. Plant Biol. 2010, 13, 193–205. [Google Scholar] [CrossRef] [PubMed]
- VSN International. Genstat for Windows, 23rd ed.; VSN International: Hemel Hempstead, UK, 2023. [Google Scholar]
- Patterson, N.; Price, A.L.; Reich, D. Population Structure and Eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.G.; Haddadi, P.; Wan, J.; Adam, L.; Walker, P.; Larkan, N.J.; Daayf, F.; Hossein Borhan, M.; Belmonte, M.F. Transcriptome Analysis of Rlm2-Mediated Host Immunity in the Brassica napus–Leptosphaeria maculans Pathosystem. Mol. Plant-Microbe Interact. 2019, 32, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Brachaczek, A.; Kaczmarek, J.; Jedryczka, M. Warm and Wet Autumns Favour Yield Losses of Oilseed Rape Caused by Phoma Stem Canker. Agronomy 2021, 11, 1171. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-Wide Importance of Phoma Stem Canker (Leptosphaeria maculans and L. biglobosa) on Oilseed Rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Dara, S.K. The New Integrated Pest Management Paradigm for the Modern Age. J. Integr. Pest Manag. 2019, 10, 12. [Google Scholar] [CrossRef]
- Ha, T.M. A Review on the Development of Integrated Pest Management and Its Integration in Modern Agriculture. Asian J. Agric. Food Sci. 2014, 2, 336–340. [Google Scholar]
- Dhawan, A.K.; Peshin, R. Integrated Pest Management: Concept, Opportunities and Challenges. Integr. Pest Manag. 2009, 1, 51–81. [Google Scholar] [CrossRef]
- Gajardo, H.A.; Wittkop, B.; Soto-Cerda, B.; Higgins, E.E.; Parkin, I.A.P.; Snowdon, R.J.; Federico, M.L.; Iniguez-Luy, F.L. Association Mapping of Seed Quality Traits in Brassica napus L. Using GWAS and Candidate QTL Approaches. Mol. Breed. 2015, 35, 143. [Google Scholar] [CrossRef]
- Helal, M.M.U.; Gill, R.A.; Tang, M.; Yang, L.; Hu, M.; Yang, L.; Xie, M.; Zhao, C.; Cheng, X.; Zhang, Y.; et al. SNP-and Haplotype-Based GWAS of Flowering-Related Traits in Brassica napus. Plants 2021, 10, 2475. [Google Scholar] [CrossRef]
- Dakouri, A.; Lamara, M.; Karim, M.M.; Wang, J.; Chen, Q.; Gossen, B.D.; Strelkov, S.E.; Hwang, S.F.; Peng, G.; Yu, F. Identification of Resistance Loci against New Pathotypes of Plasmodiophora brassicae in Brassica napus Based on Genome-Wide Association Mapping. Sci. Rep. 2021, 11, 6599. [Google Scholar] [CrossRef]
- He, Y.; Wu, D.; Wei, D.; Fu, Y.; Cui, Y.; Dong, H.; Tan, C.; Qian, W. GWAS, QTL Mapping and Gene Expression Analyses in Brassica napus Reveal Genetic Control of Branching Morphogenesis. Sci. Rep. 2017, 7, 15971. [Google Scholar] [CrossRef]
- Yang, H.; Mohd Saad, N.S.; Ibrahim, M.I.; Bayer, P.E.; Neik, T.X.; Severn-Ellis, A.A.; Pradhan, A.; Tirnaz, S.; Edwards, D.; Batley, J. Candidate Rlm6 Resistance Genes against Leptosphaeria maculans Identified through a Genome-Wide Association Study in Brassica juncea (L.) Czern. Theor. Appl. Genet. 2021, 134, 2035–2050. [Google Scholar] [CrossRef]
- Rahman, M.; Mamidi, S.; del Rio, L.; Ross, A.; Kadir, M.M.; Rahaman, M.M.; Arifuzzaman, M. Association Mapping in Brassica napus (L.) Accessions Identifies a Major QTL for Blackleg Disease Resistance on Chromosome A01. Mol. Breed. 2016, 36, 90. [Google Scholar] [CrossRef]
- Raman, H.; McVittie, B.; Pirathiban, R.; Raman, R.; Zhang, Y.; Barbulescu, D.M.; Qiu, Y.; Liu, S.; Cullis, B. Genome-Wide Association Mapping Identifies Novel Loci for Quantitative Resistance to Blackleg Disease in Canola. Front. Plant Sci. 2020, 11, 559815. [Google Scholar] [CrossRef] [PubMed]
- Cantila, A.Y.; Saad, N.S.M.; Amas, J.C.; Edwards, D.; Batley, J. Recent Findings Unravel Genes and Genetic Factors Underlying Leptosphaeria maculans Resistance in Brassica napus and Its Relatives. Int. J. Mol. Sci. 2020, 22, 313. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. Current Status of the Gene-For-Gene Concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Liban, S.H.; Cross, D.J.; Kutcher, H.R.; Peng, G.; Fernando, W.G.D. Race Structure and Frequency of Avirulence Genes in the Western Canadian Leptosphaeria maculans Pathogen Population, the Causal Agent of Blackleg in Brassica Species. Plant Pathol. 2016, 65, 1161–1169. [Google Scholar] [CrossRef]
- Van de Wouw, A.P.; Zhang, Y.; Saad, N.S.M.; Yang, H.; Sheedy, E.; Elliott, C.E.; Batley, J. Molecular Markers for Identifying Resistance Genes in Brassica napus. Agronomy 2022, 12, 985. [Google Scholar] [CrossRef]
- Raman, R.; Diffey, S.; Barbulescu, D.M.; Coombes, N.; Luckett, D.; Salisbury, P.; Cowley, R.; Marcroft, S.; Raman, H. Genetic and Physical Mapping of Loci for Resistance to Blackleg Disease in Canola (Brassica napus L.). Sci. Rep. 2020, 10, 4416. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Jestin, C.; Welham, S.J.; King, G.J.; Manzanares-Dauleux, M.J.; Fitt, B.D.L.; Delourme, R. Identification of Environmentally Stable QTL for Resistance against Leptosphaeria maculans in Oilseed Rape (Brassica Napus). Theor. Appl. Genet. 2016, 129, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Larkan, N.J.; Raman, H.; Lydiate, D.J.; Robinson, S.J.; Yu, F.; Barbulescu, D.M.; Raman, R.; Luckett, D.J.; Burton, W.; Wratten, N.; et al. Multi-Environment QTL Studies Suggest a Role for Cysteine-Rich Protein Kinase Genes in Quantitative Resistance to Blackleg Disease in Brassica napus. BMC Plant Biol. 2016, 16, 183. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; Raman, R.; Diffey, S.; Qiu, Y.; McVittie, B.; Barbulescu, D.M.; Salisbury, P.A.; Marcroft, S.; Delourme, R. Stable Quantitative Resistance Loci to Blackleg Disease in Canola (Brassica napus L.) over Continents. Front. Plant Sci. 2018, 871, 412957. [Google Scholar] [CrossRef]
- Amas, J.; Anderson, R.; Edwards, D.; Cowling, W.; Batley, J. Status and Advances in Mining for Blackleg (Leptosphaeria maculans) Quantitative Resistance (QR) in Oilseed Rape (Brassica napus). Theor. Appl. Genet. 2021, 134, 3123–3145. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Bayer, P.E.; Tirnaz, S.; Hurgobin, B.; Edwards, D.; Batley, J. Characterization of Disease Resistance Genes in the Brassica napus Pangenome Reveals Significant Structural Variation. Plant Biotechnol. J. 2020, 18, 969–982. [Google Scholar] [CrossRef]
- Shen, X.; Song, S.; Li, C.; Zhang, J. Synonymous Mutations in Representative Yeast Genes Are Mostly Strongly Non-Neutral. Nature 2022, 606, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.-H.; Zhao, Z.-Z.; He, J.-X. Molecular Sciences Brassinosteroid Signaling in Plant-Microbe Interactions. Int. J. Mol. Sci. 2018, 19, 4091. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Prasad, B.D.; Liu, Q.; Grbic, V.; Sharpe, A.; Singh, S.P.; Krishna, P. Overexpression of the Brassinosteroid Biosynthetic Gene DWF4 in Brassica napus Simultaneously Increases Seed Yield and Stress Tolerance. Sci. Rep. 2016, 6, 28298. [Google Scholar] [CrossRef] [PubMed]
- Kagale, S.; Divi, U.K.; Krochko, J.E.; Keller, W.A.; Krishna, P. Brassinosteroid Confers Tolerance in Arabidopsis thaliana and Brassica napus to a Range of Abiotic Stresses. Planta 2007, 225, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Reignault, P.; Cogan, A.; Muchembled, J.; Lounes-Hadj Sahraoui, A.; Durand, R.; Sancholle, M. Trehalose Induces Resistance to Powdery Mildew in Wheat. New Phytol. 2001, 149, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Tayeh, C.; Randoux, B.; Vincent, D.; Bourdon, N.; Reignault, P. Exogenous Trehalose Induces Defenses in Wheat Before and During a Biotic Stress Caused by Powdery Mildew. Phytopathol 2014, 104, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Du, Y.; Yu, D. Trehalose Phosphate Synthase 5-Dependent Trehalose Metabolism Modulates Basal Defense Responses in Arabidopsis thaliana. J. Integr. Plant Biol. 2019, 61, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Govind, S.R.; Jogaiah, S.; Abdelrahman, M.; Shetty, H.S.; Tran, L.S.P. Exogenous Trehalose Treatment Enhances the Activities of Defense-Related Enzymes and Triggers Resistance against Downy Mildew Disease of Pearl Millet. Front. Plant Sci. 2016, 7, 227073. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Sun, H.; Wang, X.; Jin, W.; Chen, Q.; Yuan, Z.; Yu, H. Physiological and Transcriptomic Analyses Reveal the Molecular Networks of Responses Induced by Exogenous Trehalose in Plant. PLoS ONE 2019, 14, e0217204. [Google Scholar] [CrossRef]
- Sable, A.R.; Danforth, D.; Profile, S.; Agarwal, S.K. Plant Heat Shock Protein Families: Essential Machinery for Development and Defense. J. Biol. Sci. Med. 2018, 4, 51–64. [Google Scholar]
- Shen, L.; Kang, Y.G.G.; Liu, L.; Yu, H. The J-Domain Protein J3 Mediates the Integration of Flowering Signals in Arabidopsis. Plant Cell 2011, 23, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Zhang, X.; Li, J.; Su, X.; Liu, J. Overexpression of a Tobacco J-Domain Protein Enhances Drought Tolerance in Transgenic Arabidopsis. Plant Physiol. Biochem. 2014, 83, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Salas-Muñoz, S.; Rodríguez-Hernández, A.A.; Ortega-Amaro, M.A.; Salazar-Badillo, F.B.; Jiménez-Bremont, J.F. Arabidopsis AtDjA3 Null Mutant Shows Increased Sensitivity to Abscisic Acid, Salt, and Osmotic Stress in Germination and Post-Germination Stages. Front. Plant Sci. 2016, 7, 168225. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Xu, M.Z.; Gao, S.Q.; Zhang, Y.; Yang, H.U.; Peng, J.I.N.; Cai, L.N.; Cheng, Y.; Chen, J.P.; Yang, J.; et al. Genome-Wide Identification and Analysis of the Regulation Wheat DnaJ Family Genes Following Wheat Yellow Mosaic Virus Infection. J. Integr. Agric. 2022, 21, 153–169. [Google Scholar] [CrossRef]
- Wang, G.; Cai, G.; Kong, F.; Deng, Y.; Ma, N.; Meng, Q. Overexpression of Tomato Chloroplast-Targeted DnaJ Protein Enhances Tolerance to Drought Stress and Resistance to Pseudomonas solanacearum in Transgenic Tobacco. Plant Physiol. Biochem. 2014, 82, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Yang, J.; Shi, Y.; Wang, X.; Wang, G.L. The DnaJ Protein OsDjA6 Negatively Regulates Rice Innate Immunity to the Blast Fungus Magnaporthe oryzae. Mol. Plant Pathol. 2018, 19, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Abd-El-Haliem, A.M.; Joosten, M.H.A.J. Plant Phosphatidylinositol-Specific Phospholipase C at the Center of Plant Innate Immunity. J. Integr. Plant Biol. 2017, 59, 164–179. [Google Scholar] [CrossRef]
- Vossen, J.H.; Abd-El-Haliem, A.; Fradin, E.F.; Van Den Berg, G.C.M.; Ekengren, S.K.; Meijer, H.J.G.; Seifi, A.; Bai, Y.; Ten Have, A.; Munnik, T.; et al. Identification of Tomato Phosphatidylinositol-Specific Phospholipase-C (PI-PLC) Family Members and the Role of PLC4 and PLC6 in HR and Disease Resistance. Plant J. 2010, 62, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Munnik, T.; Testerink, C. Plant Phospholipid Signaling: “In a Nutshell”. J. Lipid Res. 2009, 50, S260–S265. [Google Scholar] [CrossRef]
- Laxalt, A.M.; Munnik, T. Phospholipid Signalling in Plant Defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef]
- Mansouripour, S.; Oladzad, A.; Shahoveisi, F.; Rahman, M.M.; del Río Mendoza, L.E.; Mamidi, S.; Moghaddam, S.M. Identification of Genomic Regions Associated with Resistance to Blackleg (Leptosphaeria maculans) in Canola Using Genome Wide Association Study. Eur. J. Plant Pathol. 2021, 161, 693–707. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Coombes, N.; Song, J.; Diffey, S.; Kilian, A.; Lindbeck, K.; Barbulescu, D.M.; Batley, J.; Edwards, D.; et al. Genome-Wide Association Study Identifies New Loci for Resistance to Leptosphaeria maculans in Canola. Front. Plant Sci. 2016, 7, 1513. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Paillard, S.; Fopa-Fomeju, B.; Falentin, C.; Deniot, G.; Baron, C.; Vallée, P.; Manzanares-Dauleux, M.J.; Delourme, R. Multi-Year Linkage and Association Mapping Confirm the High Number of Genomic Regions Involved in Oilseed Rape Quantitative Resistance to Blackleg. Theor. Appl. Genet. 2018, 131, 1627–1643. [Google Scholar] [CrossRef]
- Fikere, M.; Barbulescu, D.M.; Malmberg, M.M.; Spangenberg, G.C.; Cogan, N.O.I.; Daetwyler, H.D. Meta-Analysis of GWAS in Canola Blackleg (Leptosphaeria maculans) Disease Traits Demonstrates Increased Power from Imputed Whole-Genome Sequence. Sci. Rep. 2020, 10, 14300. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).