
Human Leukocyte Antigen and microRNAs as Key Orchestrators of Mild Cognitive Impairment and Alzheimer’s Disease: A Systematic Review

 , and
, and
Abstract
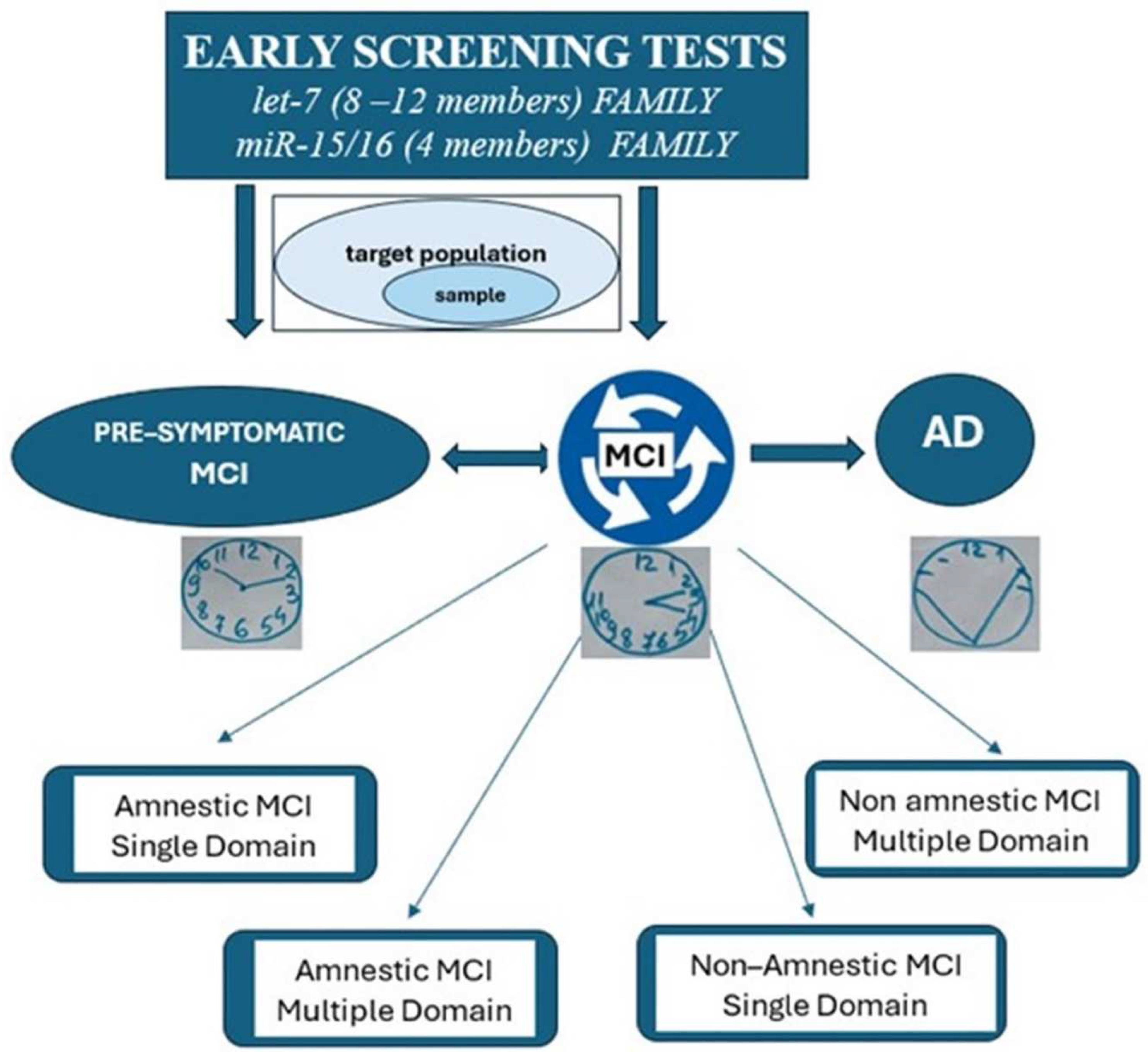
:1. Introduction
2. Inflammatory microRNAs as New Players in MCI and AD
3. Human Leukocyte Antigen (HLA) Genetic Variants in MCI and AD
4. Systematic Review Methods
4.1. Information Sources
4.2. Search Strategy
4.3. Eligibility Criteria
4.4. Selection Process
4.5. Data Collection Process and Data Items
5. Meta-Analysis Methods
6. Results

{kind=link}
{kind=link}
| HLA Type | Effects on Cognitive Impairment | References |
|---|---|---|
| HLA class II | ||
| HLA-DR15 haplotype |
| [40] |
| [46] | |
| [47] | |
| [48] | |
| TAP2 rs241448 HLA-DRB1 (linkage disequilibrium) in Caucasian populations |
| [49,50,51] |
| HLA-DQB1*06:01 (Asian populations) |
| [42,52,53] |
| HLA-DQB1*03:01 (all populations) |
| [42,52,53] |
| Single-nucleotide polymorphisms (SNP) on HLA-DRB1/DQB1: rs9271192, rs35445101, rs1130399, rs2854275, and rs28746809 |
| [42,43,52,54,55,56] |
| HLA-DRB1*09:01 |
| [42,52,54,57,58,59,60] |
| HLA-DRB1∗13:02 |
| [59] |
| HLA-DRB1*04:04 | [61] | |
| HLA-DRB1*04:01 | [62] | |
| HLA-DQA1 |
| [57,60,62,63,64,65] |
| HLA class I | ||
| HLA-B HLA-B*4402 |
| [66,67,68] |
| HLA-A2 HLA-A*33:01 |
| [42,69,70] |
| HLA-A1 HLA-A1*01 |
| [69,71,72] |
| Homeostatic iron regulator (HFE) (HLA-H)-C282Y HFE (HLA-H)-H63D mutation (high iron) |
| [73,74,75,76,77] |
| HLA-DPB1 |
| [59] |
7. Discussion
Limitations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Catana, C.-S.; Crișan, C.-A.; Opre, D.; Berindan-Neagoe, I. Implications of Long Non-Coding RNAs in Age-Altered Proteostasis. Aging Dis. 2020, 11, 692–704. [Google Scholar] [CrossRef] [PubMed]
- Cătană, C.S.; Berindan-Neagoe, I. Aging and Immunity; Lambert Academic Publishing: Saarbrucken, Germany, 2012; pp. 6–10. [Google Scholar]
- Cătană, C.S.; Pichler, M.; Giannelli, G.; Mader, R.M.; Berindan-Neagoe, I. Non-coding RNAs, the Trojan horse in two-way communication between tumor and stroma in colorectal and hepatocellular carcinoma. Oncotarget 2017, 8, 29519–29534. [Google Scholar] [CrossRef] [PubMed]
- CDC-Alzheimer’s Disease and Healthy Aging. Available online: http://www.cdc.gov/aging/data/ (accessed on 18 June 2024).
- World Health Organization. Dementia. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 26 February 2020).
- Frisoni, G.B.; Altomare, D.; Ribaldi, F.; Villain, N.; Brayne, C.; Mukadam, N.; Abramowicz, M.; Barkhof, F.; Berthier, M.; Bieler-Aeschlimann, M.; et al. Dementia prevention in memory clinics: Recommendations from the European task force for brain health services. Lancet Reg. Health-Eur. 2023, 26, 100576. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.; Kim, S.E.; Kim, J.P.; Jang, H.; Park, C.J.; Kim, H.J.; Na, D.L.; Seo, S.W.; Suk, H.-I. Deep learning model for individualized trajectory prediction of clinical outcomes in mild cognitive impairment. Front. Aging Neurosci. 2024, 16, 1356745. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, G.; Crescioli, G.; Cavedo, E.; Lucenteforte, E.; Casazza, G.; Bellatorre, A.G.; Lista, C.; Costantino, G.; Frisoni, G.; Virgili, G.; et al. Structural magnetic resonance imaging for the early diagnosis of dementia due to Alzheimer’s disease in people with mild cognitive impairment. Cochrane Database Syst. Rev. 2020, 3, CD009628. [Google Scholar] [PubMed]
- Bradfield, N.I. Mild Cognitive Impairment: Diagnosis and Subtypes. Clin. EEG Neurosci. 2021, 54, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Schoo, C. Mild Cognitive Impairment; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK599514/ (accessed on 30 June 2024).
- Ogonowski, N.; Salcidua, S.; Leon, T.; Chamorro-Veloso, N.; Valls, C.; Avalos, C.; Bisquertt, A.; Rentería, M.E.; Orellana, P.; Duran-Aniotz, C. Systematic Review: microRNAs as Potential Biomarkers in Mild Cognitive Impairment Diagnosis. Front. Aging Neurosci. 2022, 13, 807764. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Dolui, S.; Das, S.R.; Stockbower, G.E.; Daffner, M.; Rao, H.; Yushkevich, P.A.; Detre, J.A.; Wolk, D.A. A brain stress test: Cerebral perfusion during memory encoding in mild cognitive impairment. NeuroImage Clin. 2016, 11, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Stoicea, N.; Du, A.; Lakis, D.C.; Tipton, C.; Arias-Morales, C.E.; Bergese, S.D. The MiRNA Journey from Theory to Practice as a CNS Biomarker. Front. Genet. 2016, 7, 11. [Google Scholar] [CrossRef]
- Frisoni, G.B.; Hansson, O. Clinical validity of CSF biomarkes for Alzheimer’s disease: Necessary indeed, but sufficient? Lancet Neurol. 2016, 15, 650–651. [Google Scholar] [CrossRef]
- Snyder, P.J.; Jackson, C.E.; Petersen, R.C.; Khachaturian, A.S.; Kaye, J.; Albert, M.S.; Weintraub, S. Assessment of cognition in mild cognitive impairment: A comparative study. Alzheimer’s Dement. 2011, 7, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Sabahi, Z.; Farhoudi, M.; Naseri, A.; Talebi, M. Working memory assessment using cambridge neuropsychological test automated battery can help in the diagnosis of mild cognitive impairment: A systematic review and meta-analysis. Dement. Neuropsychol. 2022, 16, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Libon, D.; Lippa, C. Mild Cognitive Impairment: A Brief Review and Suggested Clinical Algorithm. Am. J. Alzheimers Dis. Other Demen 2014, 29, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C. Mild Cognitive Impairment. Continuum 2016, 22, 404–418. [Google Scholar] [PubMed]
- Petersen, R.C. Mild cognitive impairment as a diagnostic entity. J. Intern. Med. 2004, 256, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.; Wang, Z.; Initiative, F.T.A.D.N. Longitudinal Cerebral Blood Flow Changes in Normal Aging and the Alzheimer’s Disease Continuum Identified by Arterial Spin Labeling MRI. J. Alzheimer’s Dis. 2021, 81, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wang, J.; Dove, A.; Chen, H.; Yuan, C.; Bennett, D.A.; Xu, W. Body Mass Index Trajectories Preceding Incident Mild Cognitive Impairment and Dementia. JAMA Psychiatry 2022, 79, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Stern, Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef]
- Stern, Y. Cognitive Reserve: Implications for Assessment and Intervention. Folia Phoniatr. Logop. 2013, 65, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Amieva, H.; Mokri, H.; Le Goff, M.; Meillon, C.; Jacqmin-Gadda, H.; Foubert-Samier, A.; Orgogozo, J.-M.; Stern, Y.; Dartigues, J.-F. Compensatory mechanisms in higher-educated subjects with Alzheimer’s disease: A study of 20 years of cognitive decline. Brain 2014, 137, 1167–1175. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Koros, C.; Hatzimanolis, A.; Stefanis, L.; Scarmeas, N.; Papageorgiou, S.G. Exploring the Genetic Landscape of Mild Behavioral Impairment as an Early Marker of Cognitive Decline: An Updated Review Focusing on Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 2645. [Google Scholar] [CrossRef] [PubMed]
- Abuelezz, N.Z.; Nasr, F.E.; AbdulKader, M.A.; Bassiouny, A.R.; Zaky, A. MicroRNAs as Potential Orchestrators of Alzheimer’s Disease-Related Pathologies: Insights on Current Status and Future Possibilities. Front. Aging Neurosci. 2021, 13, 743573. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wang, L. Inflamma-MicroRNAs in Alzheimer’s Disease: From Disease Pathogenesis to Therapeutic Potentials. Front. Cell. Neurosci. 2021, 15, 785433. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Catana, C.; Calin, G.; Berindan-Neagoe, I. NCRNA Combined Therapy as Future Treatment Option for Cancer. Curr. Pharm. Des. 2014, 20, 6565–6574. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Tudoran, O.; Balacescu, L.; Cătană, C.S.; Neagoe, E.; Berindan-Neagoe, I.; Ionescu, C. The significance of PDGF expression in serum of colorectal carcinoma patients-correlation with Dukes classification. Can PDGF become a potential biomarker. Chirurgia 2013, 108, 849–854. [Google Scholar] [PubMed]
- Cătană, C.S.; Berindan Neagoe, I.; Opre, D. The role of circulating inflamma-microRNAs in memory performance, age-related diseases and tumorigenesis. In Proceedings of the International Conference on Cognitive and Neural Mechanisms of Human Memory, Cluj-Napoca, Romania, 18–20 May 2016. [Google Scholar]
- Grinkevich, L.N. The role of microRNAs in learning and long-term memory. Vavilov J. Genet. Breed. 2020, 24, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Liu, R.; Terpstra, E.; Wang, Y.; Qiao, F.; Wang, J.; Tong, Y.; Pan, B. Dysregulation and Diagnostic Potential of microRNA in Alzheimer’s Disease. J. Alzheimer’s Dis. 2015, 49, 1–12. [Google Scholar] [CrossRef]
- Li, W.; Zheng, Y. MicroRNAs in Extracellular Vesicles of Alzheimer’s Disease. Cells 2023, 12, 1378. [Google Scholar] [CrossRef]
- Hicks, S.D.; Ignacio, C.; Gentile, K.; Middleton, F.A. Salivary miRNA profiles identify children with autism spectrum disorder, correlate with adaptive behavior, and implicate ASD candidate genes involved in neurodevelopment. BMC Pediatr. 2016, 16, 52. [Google Scholar] [CrossRef]
- Garrido-Torres, N.; Guzmán-Torres, K.; García-Cerro, S.; Bermúdez, G.P.; Cruz-Baquero, C.; Ochoa, H.; García-González, D.; Canal-Rivero, M.; Crespo-Facorro, B.; Ruiz-Veguilla, M. miRNAs as biomarkers of autism spectrum disorder: A systematic review and meta-analysis. Eur. Child. Adolesc. Psychiatry 2023, 1–34. [Google Scholar] [CrossRef]
- Xu, H.; Song, K.; Da, L.-T. Dynamics of peptide loading into major histocompatibility complex class I molecules chaperoned by TAPBPR. Phys. Chem. Chem. Phys. 2022, 24, 12397–12409. [Google Scholar] [CrossRef]
- Choo, S.Y. The HLA system: Genetics, immunology, clinical testing, and clinical implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Wan, Q.; Xing, A. HLA in Alzheimer’s Disease: Genetic Association and Possible Pathogenic Roles. NeuroMolecular Med. 2020, 22, 464–473. [Google Scholar] [CrossRef]
- Kim, M.-S.; Cho, K.; Cho, M.-H.; Kim, N.-Y.; Kim, K.; Kim, D.-H.; Yoon, S.-Y. Neuronal MHC-I complex is destabilized by amyloid-β and its implications in Alzheimer’s disease. Cell Biosci. 2023, 13, 181. [Google Scholar] [CrossRef]
- Kunkle, B.W.; Grenier-Boley, B.; Sims, R.; Bis, J.C.; Damotte, V.; Naj, A.C. Genetic meta-analysis of diagnosed Alzheimer’s disease identifies new risk loci and implicates Aβ, tau, immunity and lipid processing. Nat. Genet. 2019, 51, 414–430. [Google Scholar] [CrossRef]
- Quan, M.; Cao, S.; Wang, Q.; Wang, S.; Jia, J. Genetic Phenotypes of Alzheimer’s Disease: Mechanisms and Potential Therapy. Phenomics 2023, 3, 333–349. [Google Scholar] [CrossRef]
- Wang, Z.X.; Wang, H.F.; Tan, L.; Liu, J.; Wan, Y.; Sun, F.R.; Tan, M.S.; Tan, C.C.; Jiang, T.; Tan, L.; et al. Effects of HLA-DRB1/DQB1 Genetic Variants on Neuroimaging in Healthy, Mild Cognitive Impairment, and Alzheimer’s Disease Cohorts. Mol. Neurobiol. 2017, 54, 3181–3188. [Google Scholar] [CrossRef]
- Lu, R.C.; Yang, W.; Tan, L.; Sun, F.R.; Tan, M.S.; Zhang, W.; Wang, H.F.; Tan, L. Association of HLA-DRB1 polymorphism with Alzheimer’s disease: A replication and meta-analysis. Oncotarget 2017, 8, 93219–93226. [Google Scholar] [CrossRef]
- Naidoo, L.; Arumugam, T.; Ramsuran, V. Narrative Review Explaining the Role of HLA-A, -B, and -C Molecules in COVID-19 Disease in and around Africa. Infect. Dis. Rep. 2024, 16, 380–406. [Google Scholar] [CrossRef]
- Van Kirk, C.A. Characterization and Regulation of the Major Histocompatibility Complex Class I in The Cns: Functional Implications for Brain Aging and Sexually Dimorphic Differences in Neuroinflammation. Ph.D. Thesis, The Pennsylvania State University, University Park, PA, USA, 2013. [Google Scholar]
- Gu, D.; Wang, L.; Zhang, N.; Wang, H.; Yu, X. Decrease in naturally occurring antibodies against epitopes of Alzheimer’s disease (AD) risk gene products is associated with cognitive decline in AD. J. Neuroinflamm. 2023, 20, 74. [Google Scholar] [CrossRef]
- Zota, V.; Nemirovsky, A.; Baron, R.; Fisher, Y.; Selkoe, D.J.; Altmann, D.M.; Weiner, H.L.; Monsonego, A. HLA-DR alleles in amyloid beta-peptide autoimmunity: A highly immunogenic role for the DRB1*1501 allele. J. Immunol. 2009, 183, 3522–3530. [Google Scholar] [CrossRef]
- Masterman, T.; Zhang, Z.; Hellgren, D.; Salter, H.; Anvret, M.; Lilius, L.; Lannfelt, L.; Hillert, J. APOE genotypes and disease severity in multiple sclerosis. Mult. Scler. J. 2002, 8, 98–103. [Google Scholar] [CrossRef]
- Trieu, C.; van Harten, A.C.; Leeuwis, A.E.; Exalto, L.G.; Hooghiemstra, A.M.; Verberk, I.M.; Allaart, C.P.; Rocca, H.-P.B.-L.; Kappelle, L.J.; van Oostenbrugge, R.J.; et al. Alzheimer’s Disease and Cognitive Decline in Patients with Cardiovascular Diseases Along the Heart-Brain Axis. J. Alzheimer’s Dis. 2024, 98, 987–1000. [Google Scholar] [CrossRef]
- Tissot, C.; Initiative, F.T.A.D.N.; Benedet, A.L.; Therriault, J.; Pascoal, T.A.; Lussier, F.Z.; Saha-Chaudhuri, P.; Chamoun, M.; Savard, M.; Mathotaarachchi, S.S.; et al. Plasma pTau181 predicts cortical brain atrophy in aging and Alzheimer’s disease. Alzheimer’s Res. Ther. 2021, 13, 69. [Google Scholar] [CrossRef]
- Bullido, M.J.; Martínez-García, A.; Artiga, M.J.; Aldudo, J.; Sastre, I.; Gil, P.; Coria, F.; Muñoz, D.G.; Hachinski, V.; Frank, A.; et al. A TAP2 genotype associated with Alzheimer’s disease in APOE4 carriers. Neurobiol. Aging 2007, 28, 519–523. [Google Scholar] [CrossRef]
- Shigemizu, D.; Fukunaga, K.; Yamakawa, A.; Suganuma, M.; Fujita, K.; Kimura, T.; Watanabe, K.; Mushiroda, T.; Sakurai, T.; Niida, S.; et al. The HLA-DRB1* 09: 01-DQB1* 03: 03 haplotype is associated with the risk for late-onset Alzheimer’s disease in APOE ε 4–negative Japanese adults. Aging 2024, 10, 3. [Google Scholar] [CrossRef]
- Jemimah, S.; AlShehhi, A. Alzheimer’s Disease Neuroimaging Initiative. c-Diadem: A constrained dual-input deep learning model to identify novel biomarkers in Alzheimer’s disease. BMC Med. Genom. 2023, 16, 244. [Google Scholar] [CrossRef]
- Huang, J.; Stein, T.D.; Wang, Y.; Ang, T.F.A.; Tao, Q.; Lunetta, K.L. Blood levels of MCP-1 modulate the genetic risks of Alzheimer’s disease mediated by HLA-DRB1 and APOE for Alzheimer’s disease. Alzheimers Dement. 2023, 19, 1925–1937. [Google Scholar] [CrossRef]
- Jiao, B.; Liu, X.; Zhou, L.; Wang, M.H.; Zhou, Y.; Xiao, T.; Zhang, W.; Sun, R.; Waye, M.M.Y.; Tang, B.; et al. Polygenic Analysis of Late-Onset Alzheimer’s Disease from Mainland China. PLoS ONE 2015, 10, e0144898. [Google Scholar] [CrossRef]
- Pandey, J.P.; Nietert, P.J.; Kothera, R.T.; Barnes, L.L.; Bennett, D.A. Interactive Effects of HLA and GM Alleles on the Development of Alzheimer Disease. Neurol. Genet. 2021, 7, e565. [Google Scholar] [CrossRef]
- Steele, N.Z.R.; Carr, J.S.; Bonham, L.W.; Geier, E.G.; Damotte, V.; Miller, Z.A.; Desikan, R.S.; Boehme, K.L.; Mukherjee, S.; Crane, P.K.; et al. Fine-mapping of the human leukocyte antigen locus as a risk factor for Alzheimer disease: A case–control study. PLOS Med. 2017, 14, e1002272. [Google Scholar] [CrossRef]
- Mansouri, L.; Klai, S.; Gritli, N.; Fekih-Mrissa, N.; Messalmani, M.; Bedoui, I.; Derbali, H.; Mrissa, R. Association of HLA-DR/DQ Polymorphism with Alzheimer’s Disease. Am. J. Med. Sci. 2015, 349, 334–337. [Google Scholar] [CrossRef]
- Le Guen, Y.; Luo, G.; Ambati, A.; Damotte, V.; Jansen, I.; Yu, E.; Nicolas, A.; De Rojas, I.; Peixoto Leal, T.; Miyashita, A.; et al. Multiancestry analysis of the HLA locus in Alzheimer’s and Parkinson’s diseases uncovers a shared adaptive immune response mediated by HLA-DRB1*04 subtypes. Proc. Natl. Acad. Sci. USA 2023, 120, e2302720120. [Google Scholar] [CrossRef]
- Zhang, X.; Zou, M.; Wu, Y.; Jiang, D.; Wu, T.; Zhao, Y.; Wu, D.; Cui, J.; Li, G. Regulation of the Late Onset Alzheimer’s Disease Associated HLA-DQA1/DRB1 Expression. Am. J. Alzheimers Dis. Other Dement. 2022, 37, 15333175221085066. [Google Scholar] [CrossRef]
- James, L.M.; Georgopoulos, A.P. Human Leukocyte Antigen as a Key Factor in Preventing Dementia and Associated Apolipoprotein E4 Risk. Front. Aging Neurosci. 2019, 11, 82. [Google Scholar] [CrossRef]
- Guo, F.; Kang, J.; Xu, J.; Wei, S.; Tao, J.; Dong, Y.; Ma, Y.; Tian, H.; Guo, X.; Bi, S.; et al. Genome-wide identification of m6A-associated single nucleotide polymorphisms in complex diseases of nervous system. Neurosci. Lett. 2023, 817, 137513. [Google Scholar] [CrossRef]
- Yokoyama, J.S.; Wang, Y.; Schork, A.J.; Thompson, W.K.; Karch, C.M.; Cruchaga, C.; McEvoy, L.K.; Witoelar, A.; Chen, C.-H.; Holland, D.; et al. Association between Genetic Traits for Immune-Mediated Diseases and Alzheimer Disease. JAMA Neurol. 2016, 73, 691–697. [Google Scholar] [CrossRef]
- Lin, K.; Yu, Y. Causal associations between neuroinflammation and Alzheimer’s disease risk. In Proceedings of the 2022 11th International Conference on Bioinformatics and Biomedical Science, Nanning, China, 28–30 October 2022. [Google Scholar]
- Panitch, R.; Hu, J.; Xia, W.; Bennett, D.A.; Stein, T.D.; Farrer, L.A.; Jun, G.R. Blood and brain transcriptome analysis reveals APOE genotype-mediated and immune-related pathways involved in Alzheimer disease. Alzheimer’s Res. Ther. 2022, 14, 30. [Google Scholar] [CrossRef]
- Vavougios, G.D.; Mavridis, T.; Doskas, T.; Papaggeli, O.; Foka, P.; Hadjigeorgiou, G. SARS-CoV-2-Induced Type I Interferon Signaling Dysregulation in Olfactory Networks Implications for Alzheimer’s Disease. Curr. Issues Mol. Biol. 2024, 46, 4565–4579. [Google Scholar] [CrossRef]
- Duan, T.; Chu, J.; Hu, F. Identification of Peripheral Blood GZMK+ CD8 + T Cells as Biomarkers of Alzheimer’s Disease Based on Single-Cell Transcriptome. Sichuan Da Xue Xue Bao Yi Xue Ban. 2023, 54, 863–873. [Google Scholar]
- Dai, Y.; Jia, P.; Zhao, Z.; Gottlieb, A. A Method for Bridging Population-Specific Genotypes to Detect Gene Modules Associated with Alzheimer’s Disease. Cells 2022, 11, 2219. [Google Scholar] [CrossRef]
- Feingold, K.R. Lipid and Lipoprotein Levels in Patients with COVID-19 Infections. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2022. [Google Scholar]
- Rosset, M.B.; Lui, G.; Dansokho, C.; Chaigneau, T.; Dorothée, G. Vaccine-induced Aβ-specific CD8+ T cells do not trigger autoimmune neuroinflammation in a murine model of Alzheimer’s disease. J. Neuroinflamm. 2015, 12, 95. [Google Scholar] [CrossRef]
- Walker, D.G.; Lue, L.-F. Immune phenotypes of microglia in human neurodegenerative disease: Challenges to detecting microglial polarization in human brains. Alzheimer’s Res. Ther. 2015, 7, 56. [Google Scholar] [CrossRef]
- Guerini, F.R.; Tinelli, C.; Calabrese, E.; Agliardi, C.; Zanzottera, M.; De Silvestri, A.; Franceschi, M.; Grimaldi, L.M.E.; Nemni, R.; Clerici, M. HLA-A*01 is associated with late onset of Alzheimer’s disease in Italian patients. Int. J. Immunopathol. Pharmacol. 2009, 22, 991–999. [Google Scholar] [CrossRef]
- Moscon, S.L.M.; Connor, J.R. HFE Mutations in Neurodegenerative Disease as a Model of Hormesis. Int. J. Mol. Sci. 2024, 25, 3334. [Google Scholar] [CrossRef]
- Ali-Rahmani, F.; Schengrund, C.L.; Connor, J.R. HFE gene variants, iron, and lipids: A novel connection in Alzheimer’s disease. Front. Pharmacol. 2014, 8, 165. [Google Scholar] [CrossRef]
- Casanova, F.; Tian, Q.; Williamson, D.S.; Qian, Y.; Zweibaum, D.; Ding, J.; Atkins, J.L.; Melzer, D.; Ferrucci, L.; Pilling, L.C. MRI-derived brain iron, grey matter volume, and risk of dementia and Parkinson’s disease: Observational and genetic analysis in the UK Biobank cohort. Neurobiol. Dis. 2024, 197, 106539. [Google Scholar] [CrossRef]
- Tisato, V.; Zuliani, G.; Vigliano, M.; Longo, G.; Franchini, E.; Secchiero, P.; Zauli, G.; Paraboschi, E.M.; Singh, A.V.; Serino, M.L.; et al. Gene-gene interactions among coding genes of iron-homeostasis proteins and APOE-alleles in cognitive impairment diseases. PLoS ONE 2018, 13, e0193867. [Google Scholar] [CrossRef]
- Kim, Y.; Connor, J.R. The roles of iron and HFE genotype in neurological diseases. Mol. Asp. Med. 2020, 75, 100867. [Google Scholar] [CrossRef]
- Cătană, C.-S.; Crișan, C.-A.; Opre, D.; Berindan-Neagoe, I. Diagnostic and prognostic value of microRNAs for Alzheimer’s disease: A comprehensive meta-analysis. Med. Pharm. Rep. 2020, 93, 53–61. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J.; Xia, Y.; Zhang, L.; Jiang, Y.; Liu, M.; Gao, Q.; Zhang, C. Harnessing the potential of HLA-G in cancer therapy: Advances, challenges, and prospects. J. Transl. Med. 2024, 22, 130. [Google Scholar] [CrossRef]
- Scavuzzi, B.M.; van Drongelen, V.; Holoshitz, J. HLA-G and the MHC Cusp Theory. Front. Immunol. 2022, 13, 814967. [Google Scholar] [CrossRef]
- Müller, M.; Kuiperij, H.B.; Claassen, J.A.; Küsters, B.; Verbeek, M.M. MicroRNAs in Alzheimer’s disease: Differential expression in hippocampus and cell-free cerebrospinal fluid. Neurobiol. Aging 2014, 35, 152–158. [Google Scholar] [CrossRef]
- Guo, S.; Jin, Y.; Zhou, J.; Zhu, Q.; Jiang, T.; Bian, Y.; Zhang, R.; Chang, C.; Xu, L.; Shen, J.; et al. MicroRNA Variants and HLA-miRNA Interactions are Novel Rheumatoid Arthritis Susceptibility Factors. Front. Genet. 2021, 12, 747274. [Google Scholar] [CrossRef]
- Friedrich, M.; Vaxevanis, C.K.; Biehl, K.; Mueller, A.; Seliger, B. Targeting the coding sequence: Opposing roles in regulating classical and non-classical MHC class I molecules by miR-16 and miR-744. J. Immunother. Cancer 2020, 8, e000396. [Google Scholar] [CrossRef]
- Mohsin, M.; Singh, P.; Khan, S.; Verma, A.K.; Jha, R.; Alsahli, M.A.; Rahmani, A.H.; Almatroodi, S.A.; Alrumaihi, F.; Kaprwan, N.; et al. Integrated transcriptomic and regulatory network analyses uncovers the role of let-7b-5p, SPIB, and HLA-DPB1 in sepsis. Sci. Rep. 2022, 12, 11963. [Google Scholar] [CrossRef]
- Li, G.; Wakao, S.; Kitada, M.; Dezawa, M. Tumor suppressor let-7 acts as a key regulator for pluripotency gene expression in Muse cells. Cell. Mol. Life Sci. 2024, 81, 54. [Google Scholar] [CrossRef]
- Kulski, J.K.; Pfaff, A.L.; Marney, L.D.; Fröhlich, A.; Bubb, V.J.; Quinn, J.P.; Koks, S. al. Regulation of expression quantitative trait loci by SVA retrotransposons within the major histocompatibility complex. Exp. Biol. Med. 2023, 248, 2304–2318. [Google Scholar]
- Kulski, J.K.; Suzuki, S.; Shiina, T.; Pfaff, A.L.; Kõks, S. Regulatory SVA retrotransposons and classical HLA genotyped-transcripts associated with Parkinson’s disease. Front. Immunol. 2024, 15, 1349030. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cătană, C.S.; Marta, M.M.; Văleanu, M.; Dican, L.; Crișan, C.A. Human Leukocyte Antigen and microRNAs as Key Orchestrators of Mild Cognitive Impairment and Alzheimer’s Disease: A Systematic Review. Int. J. Mol. Sci. 2024, 25, 8544. https://doi.org/10.3390/ijms25158544
Cătană CS, Marta MM, Văleanu M, Dican L, Crișan CA. Human Leukocyte Antigen and microRNAs as Key Orchestrators of Mild Cognitive Impairment and Alzheimer’s Disease: A Systematic Review. International Journal of Molecular Sciences. 2024; 25(15):8544. https://doi.org/10.3390/ijms25158544
Chicago/Turabian StyleCătană, Cristina Sorina, Monica Mihaela Marta, Mădălina Văleanu, Lucia Dican, and Cătălina Angela Crișan. 2024. "Human Leukocyte Antigen and microRNAs as Key Orchestrators of Mild Cognitive Impairment and Alzheimer’s Disease: A Systematic Review" International Journal of Molecular Sciences 25, no. 15: 8544. https://doi.org/10.3390/ijms25158544