
Machine Learning-Driven Biomarker Discovery for Skeletal Complications in Type 1 Gaucher Disease Patients

 , , and
, , and
Abstract
:1. Introduction
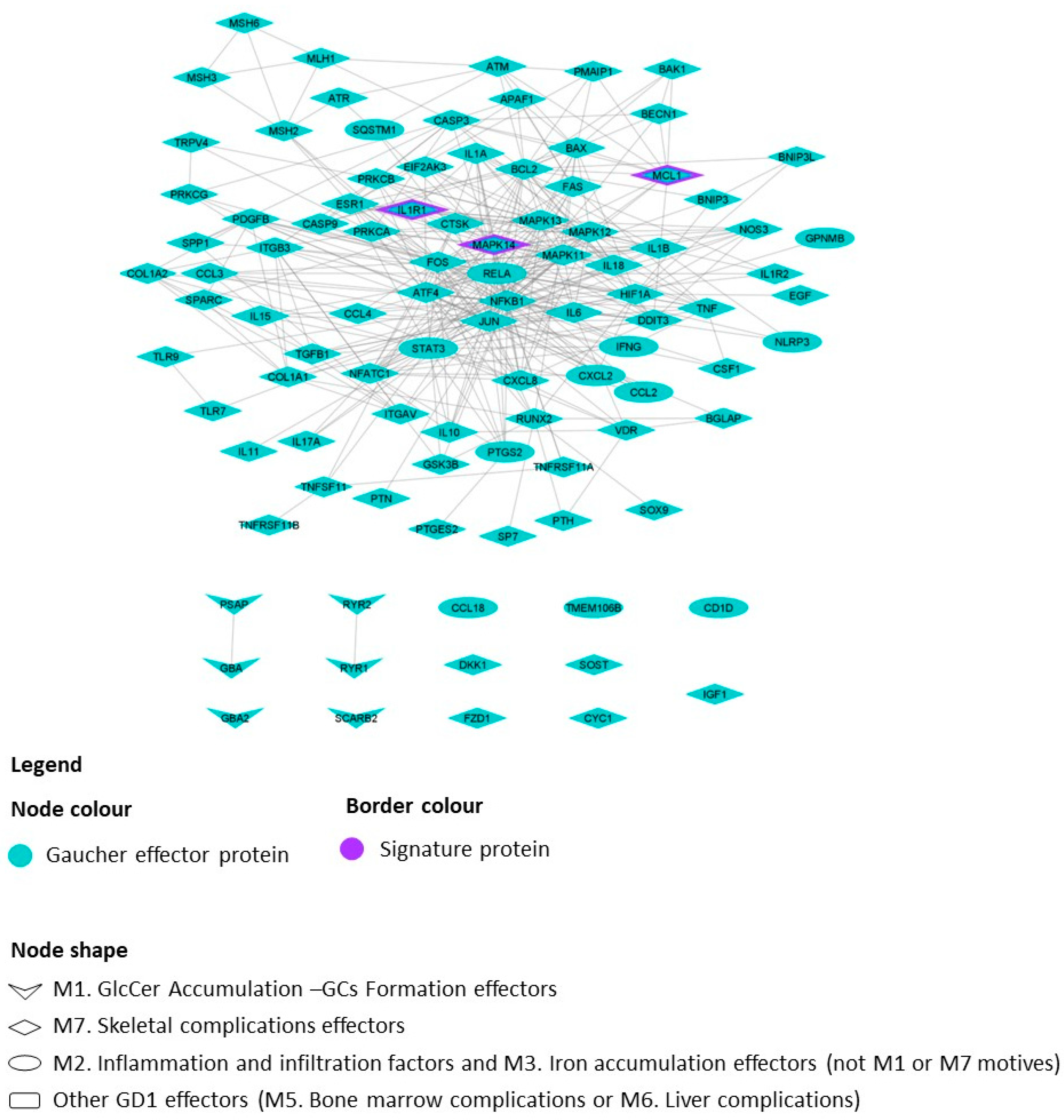
2. Results
2.1. GD1 Disease Characterization, Interactome and Mathematical Models
- -
- Biomarker candidates for skeletal complications: GD1 with skeletal complications model vs. the other models.
- -
- Biomarker candidates for bone marrow complications: GD1 with bone marrow complication models vs. the other models.
- -
- Biomarker candidates for liver complications: GD1 with liver complication models vs. the other models.
2.2. Evaluation of Previously Described Biomarkers
2.3. Identification of Potential Biomarkers of GD1-Related Skeletal Complications
3. Discussion
4. Materials and Methods
4.1. Data Collection and Gaucher Disease Interactome Characterization
4.2. Generation of Mathematical Models
4.3. Results Prediction
4.3.1. Statistical Analysis: Classifier Identification
4.3.2. Measurability Information
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saudubray, J.-M.; Mochel, F.; Lamari, F.; Garcia-Cazorla, A. Proposal for a simplified classification of IMD based on a pathophysiological approach: A practical guide for clinicians. J. Inherit. Metab. Dis. 2019, 42, 706–727. [Google Scholar] [CrossRef]
- Hughes, D.; Mikosch, P.; Belmatoug, N.; Carubbi, F.; Cox, T.; Goker-Alpan, O.; Kindmark, A.; Mistry, P.; Poll, L.; Weinreb, N.; et al. Gaucher Disease in Bone: From Pathophysiology to Practice. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2019, 34, 996–1013. [Google Scholar] [CrossRef]
- Castillon, G.; Chang, S.-C.; Moride, Y. Global Incidence and Prevalence of Gaucher Disease: A Targeted Literature Review. J. Clin. Med. 2022, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Sidransky, E. Gaucher disease: Complexity in a “simple” disorder. Mol. Genet. Metab. 2004, 83, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Mistry, P.K.; Belmatoug, N.; vom Dahl, S.; Giugliani, R. Understanding the natural history of Gaucher disease. Am. J. Hematol. 2015, 90 (Suppl. 1), S6–S11. [Google Scholar] [CrossRef] [PubMed]
- Stirnemann, J.; Belmatoug, N.; Camou, F.; Serratrice, C.; Froissart, R.; Caillaud, C.; Levade, T.; Astudillo, L.; Serratrice, J.; Brassier, A.; et al. A Review of Gaucher Disease Pathophysiology, Clinical Presentation and Treatments. Int. J. Mol. Sci. 2017, 18, 441. [Google Scholar] [CrossRef] [PubMed]
- Mikosch, P.; Hughes, D. An overview on bone manifestations in Gaucher disease. Wien. Med. Wochenschr. 2010, 160, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Charrow, J.; Andersson, H.C.; Kaplan, P.; Kolodny, E.H.; Mistry, P.; Pastores, G.; Rosenbloom, B.E.; Scott, C.R.; Wappner, R.S.; Weinreb, N.J.; et al. The Gaucher registry: Demographics and disease characteristics of 1698 patients with Gaucher disease. Arch. Intern. Med. 2000, 160, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Adar, T.; Ilan, Y.; Elstein, D.; Zimran, A. Liver involvement in Gaucher disease—Review and clinical approach. Blood Cells Mol. Dis. 2018, 68, 66–73. [Google Scholar] [CrossRef]
- Mistry, P.K.; Liu, J.; Yang, M.; Nottoli, T.; McGrath, J.; Jain, D.; Zhang, K.; Keutzer, J.; Chuang, W.-L.; Mehal, W.Z.; et al. Glucocerebrosidase gene-deficient mouse recapitulates Gaucher disease displaying cellular and molecular dysregulation beyond the macrophage. Proc. Natl. Acad. Sci. USA 2010, 107, 19473–19478. [Google Scholar] [CrossRef]
- Rozenfeld, P.A.; Crivaro, A.N.; Ormazabal, M.; Mucci, J.M.; Bondar, C.; Delpino, M.V. Unraveling the mystery of Gaucher bone density pathophysiology. Mol. Genet. Metab. 2021, 132, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Baris, H.N.; Cohen, I.J.; Mistry, P.K. Gaucher disease: The metabolic defect, pathophysiology, phenotypes and natural history. Pediatr. Endocrinol. Rev. 2014, 12 (Suppl. 1), 72–81. [Google Scholar] [PubMed]
- Simpson, W.L.; Hermann, G.; Balwani, M. Imaging of Gaucher disease. World J. Radiol. 2014, 6, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Kuter, D.J.; Salek, S.S.; Belmatoug, N.; Bembi, B.; Bright, J.; vom Dahl, S.; Deodato, F.; Di Rocco, M.; Göker-Alpan, O.; et al. Presenting signs and patient co-variables in Gaucher disease: Outcome of the Gaucher Earlier Diagnosis Consensus (GED-C) Delphi initiative. Intern. Med. J. 2019, 49, 578–591. [Google Scholar] [CrossRef] [PubMed]
- Lafforgue, P.; Trijau, S. Bone infarcts: Unsuspected gray areas? Jt. Bone Spine 2016, 83, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Gervas-Arruga, J.; Cebolla, J.J.; de Blas, I.; Roca, M.; Pocovi, M.; Giraldo, P. The influence of genetic variability and proinflammatory status on the development of bone disease in patients with Gaucher disease. PLoS ONE 2015, 10, e0126153. [Google Scholar] [CrossRef]
- Davidson, B.A.; Hassan, S.; Garcia, E.J.; Tayebi, N.; Sidransky, E. Exploring genetic modifiers of Gaucher disease: The next horizon. Hum. Mutat. 2018, 39, 1739–1751. [Google Scholar] [CrossRef]
- Mistry, P.K.; Sadan, S.; Yang, R.; Yee, J.; Yang, M. Consequences of diagnostic delays in type 1 Gaucher disease: The need for greater awareness among hematologists-oncologists and an opportunity for early diagnosis and intervention. Am. J. Hematol. 2007, 82, 697–701. [Google Scholar] [CrossRef]
- Andrade-Campos, M.M.; de Frutos, L.L.; Cebolla, J.J.; Serrano-Gonzalo, I.; Medrano-Engay, B.; Roca-Espiau, M.; Gomez-Barrera, B.; Pérez-Heredia, J.; Iniguez, D.; Giraldo, P. Identification of risk features for complication in Gaucher’s disease patients: A machine learning analysis of the Spanish registry of Gaucher disease. Orphanet J. Rare Dis. 2020, 15, 256. [Google Scholar] [CrossRef]
- Biegstraaten, M.; Cox, T.M.; Belmatoug, N.; Berger, M.G.; Collin-Histed, T.; Vom Dahl, S.; Di Rocco, M.; Fraga, C.; Giona, F.; Giraldo, P.; et al. Management goals for type 1 Gaucher disease: An expert consensus document from the European working group on Gaucher disease. Blood Cells. Mol. Dis. 2018, 68, 203–208. [Google Scholar] [CrossRef]
- Drelichman, G.; Fernández Escobar, N.; Basack, N.; Aversa, L.; Larroude, M.S.; Aguilar, G.; Szlago, M.; Schenone, A.; Fynn, A.; Cuello, M.F.; et al. Skeletal involvement in Gaucher disease: An observational multicenter study of prognostic factors in the Argentine Gaucher disease patients. Am. J. Hematol. 2016, 91, E448–E453. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, P.; Andrade-Campos, M.; Morales, M. Recommendations on the follow-up of patients with Gaucher disease in Spain: Results from a Delphi survey. JIMD Rep. 2023, 64, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Piran, S.; Amato, D. Gaucher disease: A systematic review and meta-analysis of bone complications and their response to treatment. J. Inherit. Metab. Dis. 2010, 33, 271–279. [Google Scholar] [CrossRef]
- Fleshner, P.R.; Aufses, A.H.J.; Grabowski, G.A.; Elias, R. A 27-year experience with splenectomy for Gaucher’s disease. Am. J. Surg. 1991, 161, 69–75. [Google Scholar] [CrossRef] [PubMed]
- DeMayo, R.F.; Haims, A.H.; McRae, M.C.; Yang, R.; Mistry, P.K. Correlation of MRI-Based bone marrow burden score with genotype and spleen status in Gaucher’s disease. AJR Am. J. Roentgenol. 2008, 191, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Utz, J.; Whitley, C.B.; van Giersbergen, P.L.M.; Kolb, S.A. Comorbidities and pharmacotherapies in patients with Gaucher disease type 1: The potential for drug-drug interactions. Mol. Genet. Metab. 2016, 117, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, P.; López de Frutos, L.; Cebolla, J.J. Biomarker combination is necessary for the assessment of Gaucher disease? Ann. Transl. Med. 2018, 6, S81. [Google Scholar] [CrossRef] [PubMed]
- Maas, M.; Poll, L.W.; Terk, M.R. Imaging and quantifying skeletal involvement in Gaucher disease. Br. J. Radiol. 2002, 75 (Suppl. 1), A13–A24. [Google Scholar] [CrossRef]
- Jefferies, J.L.; Spencer, A.K.; Lau, H.A.; Nelson, M.W.; Giuliano, J.D.; Zabinski, J.W.; Boussios, C.; Curhan, G.; Gliklich, R.E.; Warnock, D.G. A new approach to identifying patients with elevated risk for Fabry disease using a machine learning algorithm. Orphanet J. Rare Dis. 2021, 16, 518. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.; Artigas, L.; Valls, R.; Gervas-Arruga, J. An in silico approach to identify early damage biomarker candidates in metachromatic leukodystrophy. Mol. Genet. Metab. Rep. 2023, 35, 100974. [Google Scholar] [CrossRef]
- You, H.; Dong, M. Prediction of diagnostic gene biomarkers for hypertrophic cardiomyopathy by integrated machine learning. J. Int. Med. Res. 2023, 51, 3000605231213781. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.B.; Robertson, D.D.; Laney, D.A.; Gambello, M.J.; Terk, M. Machine learning based analytics of micro-MRI trabecular bone microarchitecture and texture in type 1 Gaucher disease. J. Biomech. 2016, 49, 1961–1968. [Google Scholar] [CrossRef] [PubMed]
- Echle, A.; Rindtorff, N.T.; Brinker, T.J.; Luedde, T.; Pearson, A.T.; Kather, J.N. Deep learning in cancer pathology: A new generation of clinical biomarkers. Br. J. Cancer 2021, 124, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-H.; Lin, C.-H.; Lane, H.-Y. Machine Learning and Novel Biomarkers for the Diagnosis of Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 2761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Itan, Y. Biological Network Approaches and Applications in Rare Disease Studies. Genes 2019, 10, 797. [Google Scholar] [CrossRef] [PubMed]
- Segú-Vergés, C.; Coma, M.; Kessel, C.; Smeets, S.; Foell, D.; Aldea, A. Application of systems biology-based in silico tools to optimize treatment strategy identification in Still’s disease. Arthritis Res. Ther. 2021, 23, 126. [Google Scholar] [CrossRef]
- Tebani, A.; Mauhin, W.; Abily-Donval, L.; Lesueur, C.; Berger, M.G.; Nadjar, Y.; Berger, J.; Benveniste, O.; Lamari, F.; Laforêt, P.; et al. A Proteomics-Based Analysis Reveals Predictive Biological Patterns in Fabry Disease. J. Clin. Med. 2020, 9, 1325. [Google Scholar] [CrossRef] [PubMed]
- Parolo, S.; Tomasoni, D.; Bora, P.; Ramponi, A.; Kaddi, C.; Azer, K.; Domenici, E.; Neves-Zaph, S.; Lombardo, R. Reconstruction of the Cytokine Signaling in Lysosomal Storage Diseases by Literature Mining and Network Analysis. Front. Cell Dev. Biol. 2021, 9, 703489. [Google Scholar] [CrossRef]
- GEO Accession Viewer. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE21899 (accessed on 1 September 2023).
- Clough, E.; Barrett, T. The Gene Expression Omnibus Database. Methods Mol. Biol. 2016, 1418, 93–110. [Google Scholar] [CrossRef]
- Jorba, G.; Aguirre-Plans, J.; Junet, V.; Segú-Vergés, C.; Ruiz, J.L.; Pujol, A.; Fernández-Fuentes, N.; Mas, J.M.; Oliva, B. In-silico simulated prototype-patients using TPMS technology to study a potential adverse effect of sacubitril and valsartan. PLoS ONE 2020, 15, e0228926. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef]
- Nanjappa, V.; Thomas, J.K.; Marimuthu, A.; Muthusamy, B.; Radhakrishnan, A.; Sharma, R.; Ahmad Khan, A.; Balakrishnan, L.; Sahasrabuddhe, N.A.; Kumar, S.; et al. Plasma Proteome Database as a resource for proteomics research: 2014 update. Nucleic Acids Res. 2014, 42, D959–D965. [Google Scholar] [CrossRef]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Kentsis, A.; Monigatti, F.; Dorff, K.; Campagne, F.; Bachur, R.; Steen, H. Urine proteomics for profiling of human disease using high accuracy mass spectrometry. Proteom. Clin. Appl. 2009, 3, 1052–1061. [Google Scholar] [CrossRef]
- Viceconti, M.; Pappalardo, F.; Rodriguez, B.; Horner, M.; Bischoff, J.; Musuamba Tshinanu, F. In silico trials: Verification, validation and uncertainty quantification of predictive models used in the regulatory evaluation of biomedical products. Methods 2021, 185, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Simats, A.; Ramiro, L.; Valls, R.; de Ramón, H.; García-Rodríguez, P.; Orset, C.; Artigas, L.; Sardon, T.; Rosell, A.; Montaner, J. Ceruletide and Alpha-1 Antitrypsin as a Novel Combination Therapy for Ischemic Stroke. Neurother. J. Am. Soc. Exp. Neurother. 2022, 19, 513–527. [Google Scholar] [CrossRef]
- Bayes-Genis, A.; Iborra-Egea, O.; Spitaleri, G.; Domingo, M.; Revuelta-López, E.; Codina, P.; Cediel, G.; Santiago-Vacas, E.; Cserkóová, A.; Pascual-Figal, D.; et al. Decoding empagliflozin’s molecular mechanism of action in heart failure with preserved ejection fraction using artificial intelligence. Sci. Rep. 2021, 11, 12025. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, N.; Tripathi, R.; Giró, A.; Rosich, L.; López-Guerra, M.; López-Oreja, I.; Playa-Albinyana, H.; Arenas, F.; Mas, J.M.; Pérez-Galán, P.; et al. Systems biology drug screening identifies statins as enhancers of current therapies in chronic lymphocytic leukemia. Sci. Rep. 2020, 10, 22153. [Google Scholar] [CrossRef] [PubMed]
- Lorén, V.; Garcia-Jaraquemada, A.; Naves, J.E.; Carmona, X.; Mañosa, M.; Aransay, A.M.; Lavin, J.L.; Sánchez, I.; Cabré, E.; Manyé, J.; et al. ANP32E, a Protein Involved in Steroid-Refractoriness in Ulcerative Colitis, Identified by a Systems Biology Approach. J. Crohns. Colitis 2019, 13, 351–361. [Google Scholar] [CrossRef]
- Barberis, E.; Khoso, S.; Sica, A.; Falasca, M.; Gennari, A.; Dondero, F.; Afantitis, A.; Manfredi, M. Precision Medicine Approaches with Metabolomics and Artificial Intelligence. Int. J. Mol. Sci. 2022, 23, 11269. [Google Scholar] [CrossRef]
- Khan, F.; Siva Prasad, B.V.V.; Syed, S.A.; Ashraf, I.; Ramasamy, L.K. An Efficient, Ensemble-Based Classification Framework for Big Medical Data. Big Data 2022, 10, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.M.; Aerts, J.M.F.G.; Belmatoug, N.; Cappellini, M.D.; vom Dahl, S.; Goldblatt, J.; Grabowski, G.A.; Hollak, C.E.M.; Hwu, P.; Maas, M.; et al. Management of non-neuronopathic Gaucher disease with special reference to pregnancy, splenectomy, bisphosphonate therapy, use of biomarkers and bone disease monitoring. J. Inherit. Metab. Dis. 2008, 31, 319–336. [Google Scholar] [CrossRef] [PubMed]
- de Mello, R.A.F.; Mello, M.B.N.; Pessanha, L.B.; Fonseca, A.P.A. Skeletal involvement in Gaucher disease: Extent of bone disease, splenic volume, and quality of life. Radiol. Bras. 2021, 54, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Gaucher disease. Current issues in diagnosis and treatment. NIH Technology Assessment Panel on Gaucher Disease. JAMA 1996, 275, 548–553. [CrossRef]
- Pavlova, E.V.; Deegan, P.B.; Tindall, J.; McFarlane, I.; Mehta, A.; Hughes, D.; Wraith, J.E.; Cox, T.M. Potential biomarkers of osteonecrosis in Gaucher disease. Blood Cells. Mol. Dis. 2011, 46, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.J.; Myer, B.J.; Khokher, A.M.; Rushton, N.; Cox, T.M. Pro-inflammatory cytokines and the pathogenesis of Gaucher’s disease: Increased release of interleukin-6 and interleukin-10. QJM 1997, 90, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Barak, V.; Acker, M.; Nisman, B.; Kalickman, I.; Abrahamov, A.; Zimran, A.; Yatziv, S. Cytokines in Gaucher’s disease. Eur. Cytokine Netw. 1999, 10, 205–210. [Google Scholar] [PubMed]
- Ramazzotti, G.; Bavelloni, A.; Blalock, W.; Piazzi, M.; Cocco, L.; Faenza, I. BMP-2 Induced Expression of PLCβ1 That is a Positive Regulator of Osteoblast Differentiation. J. Cell. Physiol. 2016, 231, 623–629. [Google Scholar] [CrossRef]
- Purohit, V.; Gao, B.; Song, B.-J. Molecular mechanisms of alcoholic fatty liver. Alcohol Clin. Exp. Res. 2009, 33, 191–205. [Google Scholar] [CrossRef]
- Blumenreich, S.; Yaacobi, C.; Vardi, A.; Barav, O.B.; Vitner, E.B.; Park, H.; Wang, B.; Cheng, S.H.; Sardi, S.P.; Futerman, A.H. Substrate reduction therapy using Genz-667161 reduces levels of pathogenic components in a mouse model of neuronopathic forms of Gaucher disease. J. Neurochem. 2021, 156, 692–701. [Google Scholar] [CrossRef]
- Zhang, Y.; Yuan, L.; Meng, L.; Fang, M.; Guo, S.; Wang, D.; Ma, J.; Wang, L. Guanine and nucleotide binding protein 3 promotes odonto/osteogenic differentiation of apical papilla stem cells via JNK and ERK signaling pathways. Int. J. Mol. Med. 2019, 43, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, H.; Huang, J.; Pei, S.; Wang, L.; Feng, J.Q.; Jing, D.; Zhao, H.; Kronenberg, H.M.; Moore, D.C.; et al. Targeted Ptpn11 deletion in mice reveals the essential role of SHP2 in osteoblast differentiation and skeletal homeostasis. Bone Res. 2021, 9, 6. [Google Scholar] [CrossRef]
- Serfecz, J.C.; Saadin, A.; Santiago, C.P.; Zhang, Y.; Bentzen, S.M.; Vogel, S.N.; Feldman, R.A. C5a Activates a Pro-Inflammatory Gene Expression Profile in Human Gaucher iPSC-Derived Macrophages. Int. J. Mol. Sci. 2021, 22, 9912. [Google Scholar] [CrossRef]
- Liu, X.; Min, L.; Duan, H.; Shi, R.; Zhang, W.; Hong, S.; Tu, C. Short hairpin RNA (shRNA) of type 2 interleukin-1 receptor (IL1R2) inhibits the proliferation of human osteosarcoma U-2 OS cells. Med. Oncol. 2015, 32, 364. [Google Scholar] [CrossRef] [PubMed]
- Mehra, M.R.; Uber, P.A.; Benitez, R.M. Gene-based bio-signature patterns and cardiac allograft rejection. Heart Fail. Clin. 2010, 6, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Alan, S.; Şalva, E.; Yılmaz, İ.; Turan, S.Ö.; Akbuğa, J. The effectiveness of chitosan-mediated silencing of PDGF-B and PDGFR-β in the mesangial proliferative glomerulonephritis therapy. Exp. Mol. Pathol. 2019, 110, 104280. [Google Scholar] [CrossRef]
- Zhang, M.; Yu, W.; Niibe, K.; Zhang, W.; Egusa, H.; Tang, T.; Jiang, X. The Effects of Platelet-Derived Growth Factor-BB on Bone Marrow Stromal Cell-Mediated Vascularized Bone Regeneration. Stem Cells Int. 2018, 2018, 3272098. [Google Scholar] [CrossRef]
- Chen, W.; Baylink, D.J.; Brier-Jones, J.; Neises, A.; Kiroyan, J.B.; Rundle, C.H.; Lau, K.-H.W.; Zhang, X.-B. PDGFB-based stem cell gene therapy increases bone strength in the mouse. Proc. Natl. Acad. Sci. USA 2015, 112, E3893–E3900. [Google Scholar] [CrossRef]
- Pompili, V.J.; Gordon, D.; San, H.; Yang, Z.; Muller, D.W.; Nabel, G.J.; Nabel, E.G. Expression and function of a recombinant PDGF B gene in porcine arteries. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 2254–2264. [Google Scholar] [CrossRef]
- Roh, J.; Subramanian, S.; Weinreb, N.J.; Kartha, R.V. Gaucher disease—More than just a rare lipid storage disease. J. Mol. Med. 2022, 100, 499–518. [Google Scholar] [CrossRef]
- Beaton, B.; Hughes, D.A. Soluble mannose receptor: A potential biomarker in Gaucher disease. Eur. J. Haematol. 2024, 112, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Komninaka, V.; Kolomodi, D.; Christoulas, D.; Marinakis, T.; Papatheodorou, A.; Repa, K.; Voskaridou, E.; Revenas, K.; Terpos, E. Evaluation of bone involvement in patients with Gaucher disease: A semi-quantitative magnetic resonance imaging method (using ROI estimation of bone lesion) as an alternative method to semi-quantitative methods used so far. Eur. J. Haematol. 2015, 95, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Campeau, P.M.; Rafei, M.; Boivin, M.-N.; Sun, Y.; Grabowski, G.A.; Galipeau, J. Characterization of Gaucher disease bone marrow mesenchymal stromal cells reveals an altered inflammatory secretome. Blood 2009, 114, 3181–3190. [Google Scholar] [CrossRef]
- Yavropoulou, M.P.; Michopoulos, A.; Yovos, J.G. PTH and PTHR1 in osteocytes. New insights into old partners. Hormones 2017, 16, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Gardella, T.J.; Vilardaga, J.-P. International Union of Basic and Clinical Pharmacology. XCIII. The parathyroid hormone receptors—Family B G protein-coupled receptors. Pharmacol. Rev. 2015, 67, 310–337. [Google Scholar] [CrossRef] [PubMed]
- Gould, N.R.; Williams, K.M.; Joca, H.C.; Torre, O.M.; Lyons, J.S.; Leser, J.M.; Srikanth, M.P.; Hughes, M.; Khairallah, R.J.; Feldman, R.A.; et al. Disparate bone anabolic cues activate bone formation by regulating the rapid lysosomal degradation of sclerostin protein. Elife 2021, 10, e64393. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Hanley, D.A.; McNeil, C.; Boyd, S. Improvement in Bone Mineral Density and Architecture in a Patient with Gaucher Disease Using Teriparatide. JIMD Rep. 2015, 22, 23–28. [Google Scholar] [CrossRef]
- Mozafari, H.; Khatami, S.; Kiani, A.; Rahimi, Z.; Vaisi-Raygani, A.; Afsharnaderi, A.; Alaei, M.R. Oxidative Stress Parameters, Trace Elements, and Lipid Profile in Iranian Patients with Gaucher Disease. Biol. Trace Elem. Res. 2020, 193, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Athar, A.; Füllgrabe, A.; George, N.; Iqbal, H.; Huerta, L.; Ali, A.; Snow, C.; Fonseca, N.A.; Petryszak, R.; Papatheodorou, I.; et al. ArrayExpress update—From bulk to single-cell expression data. Nucleic Acids Res. 2019, 47, D711–D715. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, M.; da Veiga Leprevost, F.; Squizzato, S.; Park, Y.M.; Haug, K.; Carroll, A.J.; Spalding, D.; Paschall, J.; Wang, M.; et al. Discovering and linking public omics data sets using the Omics Discovery Index. Nat. Biotechnol. 2017, 35, 406–409. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, Article3. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Meltzer, P.S. GEOquery: A bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 2007, 23, 1846–1847. [Google Scholar] [CrossRef]
- Rivals, I.; Personnaz, L.; Taing, L.; Potier, M.-C. Enrichment or depletion of a GO category within a class of genes: Which test? Bioinformatics 2007, 23, 401–407. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, D199–D205. [Google Scholar] [CrossRef]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The Reactome pathway knowledgebase. Nucleic Acids Res. 2014, 42, D472–D477. [Google Scholar] [CrossRef] [PubMed]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; del-Toro, N.; et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef]
- Salwinski, L.; Licata, L.; Winter, A.; Thorneycroft, D.; Khadake, J.; Ceol, A.; Aryamontri, A.C.; Oughtred, R.; Livstone, M.; Boucher, L.; et al. Recurated protein interaction datasets. Nat. Methods 2009, 6, 860–861. [Google Scholar] [CrossRef] [PubMed]
- Keshava Prasad, T.S.; Goel, R.; Kandasamy, K.; Keerthikumar, S.; Kumar, S.; Mathivanan, S.; Telikicherla, D.; Raju, R.; Shafreen, B.; Venugopal, A.; et al. Human Protein Reference Database—2009 update. Nucleic Acids Res. 2009, 37, D767–D772. [Google Scholar] [CrossRef]
- Han, H.; Cho, J.-W.; Lee, S.; Yun, A.; Kim, H.; Bae, D.; Yang, S.; Kim, C.Y.; Lee, M.; Kim, E.; et al. TRRUST v2: An expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018, 46, D380–D386. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Artigas, L.; Coma, M.; Matos-Filipe, P.; Aguirre-Plans, J.; Farrés, J.; Valls, R.; Fernandez-Fuentes, N.; de la Haba-Rodriguez, J.; Olvera, A.; Barbera, J.; et al. In-silico drug repurposing study predicts the combination of pirfenidone and melatonin as a promising candidate therapy to reduce SARS-CoV-2 infection progression and respiratory distress caused by cytokine storm. PLoS ONE 2020, 15, e0240149. [Google Scholar] [CrossRef]
- Chow, C.; Liu, C. Approximating discrete probability distributions with dependence trees. IEEE Trans. Inf. Theory 1968, 14, 462–467. [Google Scholar] [CrossRef]
- Peng, H.; Long, F.; Ding, C. Feature selection based on mutual information: Criteria of max-dependency, max-relevance, and min-redundancy. IEEE Trans. Pattern Anal. Mach. Intell. 2005, 27, 1226–1238. [Google Scholar] [CrossRef]
- Bishop, C.M. Pattern Recoginiton and Machine Learning; Springer: New York, NY, USA, 2006; ISBN 9780387310732. [Google Scholar]
- Guyon, I.; Weston, J.; Barnhill, S.; Vapnik, V. Gene Selection for Cancer Classification using Support Vector Machines. Mach. Learn. 2002, 46, 389–422. [Google Scholar] [CrossRef]
- Ververidis, D.; Kotropoulos, C. Fast and accurate sequential floating forward feature selection with the Bayes classifier applied to speech emotion recognition. Signal Process. 2008, 88, 2956–2970. [Google Scholar] [CrossRef]
- Christin, C.; Hoefsloot, H.C.J.; Smilde, A.K.; Hoekman, B.; Suits, F.; Bischoff, R.; Horvatovich, P. A critical assessment of feature selection methods for biomarker discovery in clinical proteomics. Mol. Cell. Proteom. 2013, 12, 263–276. [Google Scholar] [CrossRef]
- Madsen, H.; Thyregod, P. Introduction to General and Generalized Linear Models; Routledge: London, UK, 2011; ISBN 9781420091557. [Google Scholar]
- Russell, S. Artificial Intelligence: A Modern Approach; Recording for the Blind & Dyslexic: Princeton, NJ, USA, 2003; ISBN 9780137903955. [Google Scholar]
- GEO Web Site—How to Use It. Available online: https://www.ncbi.nlm.nih.gov/geo/info/geo2r.html#how_to_use (accessed on 1 September 2023).



{kind=link}
{kind=link}
{kind=link}
| M ID | M Name | Number of Proteins |
|---|---|---|
| M1 | GlcCer accumulation–GC formation | 6 |
| M2 | Inflammation and infiltration factors | 24 |
| M3 | Iron accumulation in GCs | 12 |
| M4 | Splenic complications | 0 * |
| M5 | Bone marrow complications | 21 |
| M6 | Liver complications | 43 |
| M7 | Skeletal complications (Osteoblast/osteoclast uncoupling) | 43 |
| Skeletal complications (Hypoxia-induced osteonecrosis) | 42 |
| TPMS Model Name | Response (GD1 Motif Combination) |
|---|---|
| GD1 without complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs |
| GD1 with skeletal complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M7. Skeletal complications |
| GD1 with liver complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M6. Liver complications |
| GD1 with BM complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M5. BM complications motif |
| GD1 with BM and liver complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M5. BM complications M6. Liver complications |
| GD1 with liver and skeletal complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M6. Liver complications M7. Skeletal complications |
| GD1 with BM and skeletal complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M6. BM complications M7. Skeletal complications |
| GD1 with BM, liver and skeletal complications | M2. Inflammation and infiltration factors M3. Iron accumulation in GCs M5. BM complications M6. Liver complications M7. Skeletal complications |
| Protein Name | Gene Name [40] | UniProt ID [42] | crosVal Accuracy (%) | crosVal p-Value |
|---|---|---|---|---|
| MIP-1β | CCL4 | P13236 | 58.54 | 9.32 × 10−32 |
| Cat K | CTSK | P43235 | 58.4 | 6.72 × 10−41 |
| MIP-1α | CCL3 | P10147 | 53 | 0.000218 |
| OC | BGLAP | P02818 | 51.31 | 2.83 × 10−10 |
| Gene Name [40] | UniProt ID [42] | crosVal Accuracy (%) | crosVal Precision (%) | crosValSensitivity (%) | crosVal p-Value | Plasma Protein [43,44] | Urine Protein [45] |
|---|---|---|---|---|---|---|---|
| MCL1 | Q07820 | 66.69 | 65.32 | 71.14 | 3.62 × 10−89 | No | No |
| PLCB1 | Q9NQ66 | No | No | ||||
| JAK2 | O60674 | 66.80 | 65.46 | 71.14 | 2.37 × 10−90 | No | No |
| MCL1 | Q07820 | No | No | ||||
| IL1R2 | P27930 | 67.54 | 68.25 | 65.60 | 7.26 × 10−98 | Yes | No |
| PDGFB | P01127 | 68.31 | 65.04 | 79.20 | 1.97 × 10−112 | Yes | No |
| PTH | P01270 | Yes | No | ||||
| CCL3 | P10147 | Yes | No | ||||
| PDGFB | P01127 | 68.54 | 66.30 | 75.43 | 1.51 × 10−111 | Yes | No |
| PLCB1 | Q9NQ66 | No | No | ||||
| GNAI3 | P08754 | 70.11 | 72.25 | 65.31 | 4.77 × 10−130 | No | Yes |
| CXCL8 | P10145 | Yes | No | ||||
| MCL1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 70.26 | 67.18 | 79.20 | 1.62 × 10−135 | Yes | No |
| SREBF1 | P36956 | No | No | ||||
| IL1R2 | P27930 | 70.77 | 72.15 | 67.66 | 5.66 × 10−138 | Yes | No |
| MCL1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 70.97 | 71.92 | 68.80 | 2.29 × 10−140 | Yes | No |
| IL1R2 | P27930 | Yes | No | ||||
| IL1R2 | P27930 | 71.54 | 69.41 | 77.03 | 4.62 × 10−150 | Yes | No |
| SREBF1 | P36956 | No | No | ||||
| IL1R2 | P27930 | 73.14 | 70.19 | 80.46 | 6.74 × 10−176 | Yes | No |
| PLCB1 | Q9NQ66 | No | No | ||||
| JAK2 | O60674 | 73.83 | 70.44 | 82.11 | 2.45 × 10−188 | No | No |
| IL1R2 | P27930 | Yes | No | ||||
| PDGFB | P01127 | 74.00 | 76.12 | 69.94 | 3.55 × 10−186 | Yes | No |
| MCL1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 74.20 | 78.03 | 67.37 | 2.58 × 10−192 | Yes | No |
| MCL1 | Q07820 | No | No | ||||
| CCL3 | P10147 | Yes | No | ||||
| PDGFB | P01127 | 75.20 | 77.43 | 71.14 | 4.96 × 10−206 | Yes | No |
| PTPN11 | Q06124 | No | No | ||||
| MLC1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 76.26 | 79.82 | 70.29 | 8.24 × 10−227 | Yes | No |
| SREBF1 | P36956 | No | No | ||||
| MCL1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 76.54 | 77.86 | 74.17 | 2.31 × 10−228 | Yes | No |
| CXCL8 | P10145 | Yes | No | ||||
| MCL1 | Q07820 | No | No | ||||
| PDGFB | P01127 | 77.26 | 79.96 | 72.74 | 2.15 × 10−243 | Yes | No |
| GNAI3 | P08754 | No | Yes | ||||
| MCL1 | Q07820 | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cebolla, J.J.; Giraldo, P.; Gómez, J.; Montoto, C.; Gervas-Arruga, J. Machine Learning-Driven Biomarker Discovery for Skeletal Complications in Type 1 Gaucher Disease Patients. Int. J. Mol. Sci. 2024, 25, 8586. https://doi.org/10.3390/ijms25168586
Cebolla JJ, Giraldo P, Gómez J, Montoto C, Gervas-Arruga J. Machine Learning-Driven Biomarker Discovery for Skeletal Complications in Type 1 Gaucher Disease Patients. International Journal of Molecular Sciences. 2024; 25(16):8586. https://doi.org/10.3390/ijms25168586
Chicago/Turabian StyleCebolla, Jorge J., Pilar Giraldo, Jessica Gómez, Carmen Montoto, and Javier Gervas-Arruga. 2024. "Machine Learning-Driven Biomarker Discovery for Skeletal Complications in Type 1 Gaucher Disease Patients" International Journal of Molecular Sciences 25, no. 16: 8586. https://doi.org/10.3390/ijms25168586