
Recombinant Subunit Vaccine Candidate against the Bovine Viral Diarrhea Virus

 , , ,
, , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. BVDV Viral Survey
2.2. Production of E2 Proteins from Different BVDV Subgenotypes
2.3. Dose Response Evaluation in a Sheep Model
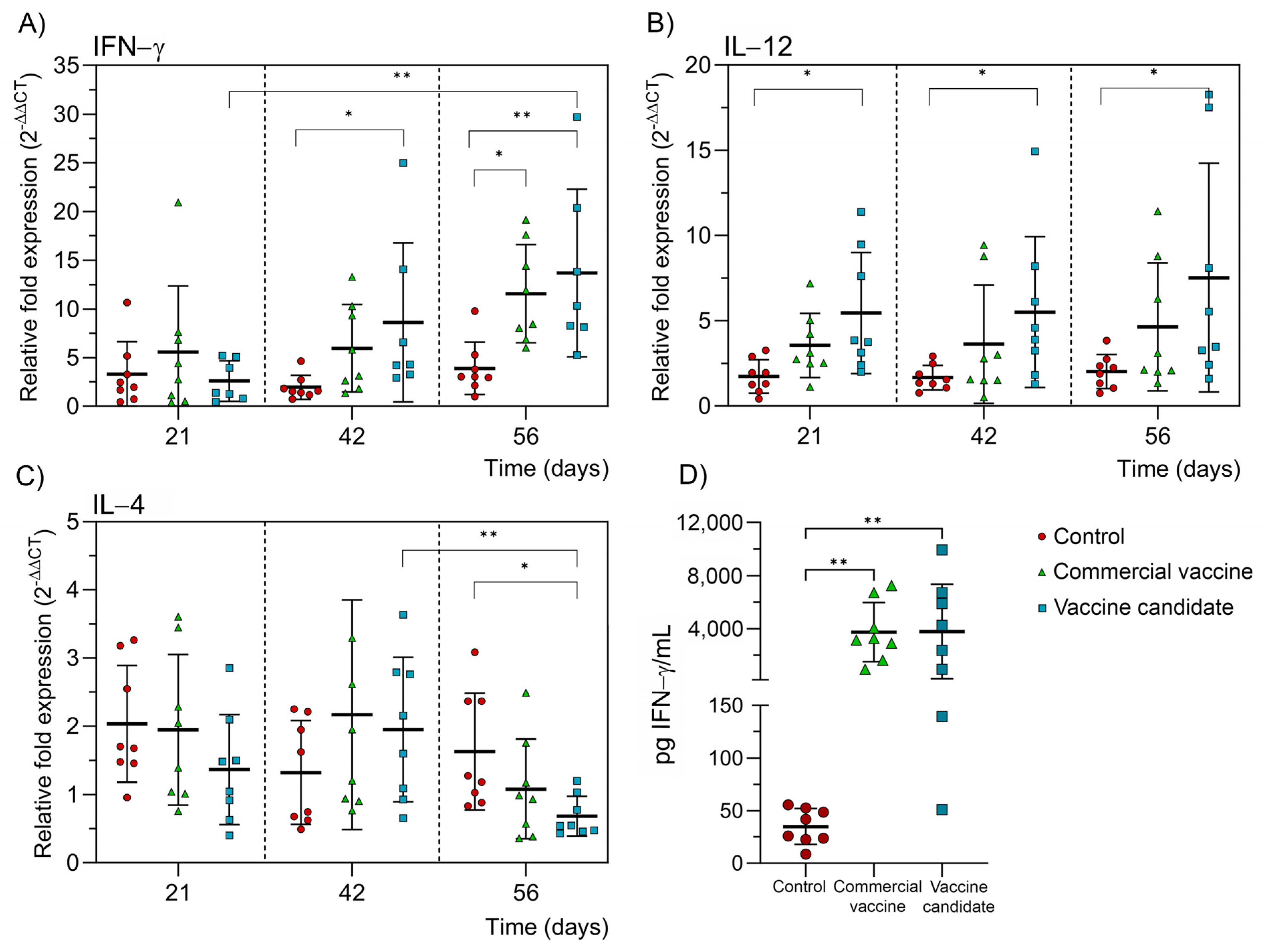
2.4. Immune Response Induced by the Vaccine Candidate in Bovines
2.4.1. Humoral Immune Response
2.4.2. Cellular Immune Response
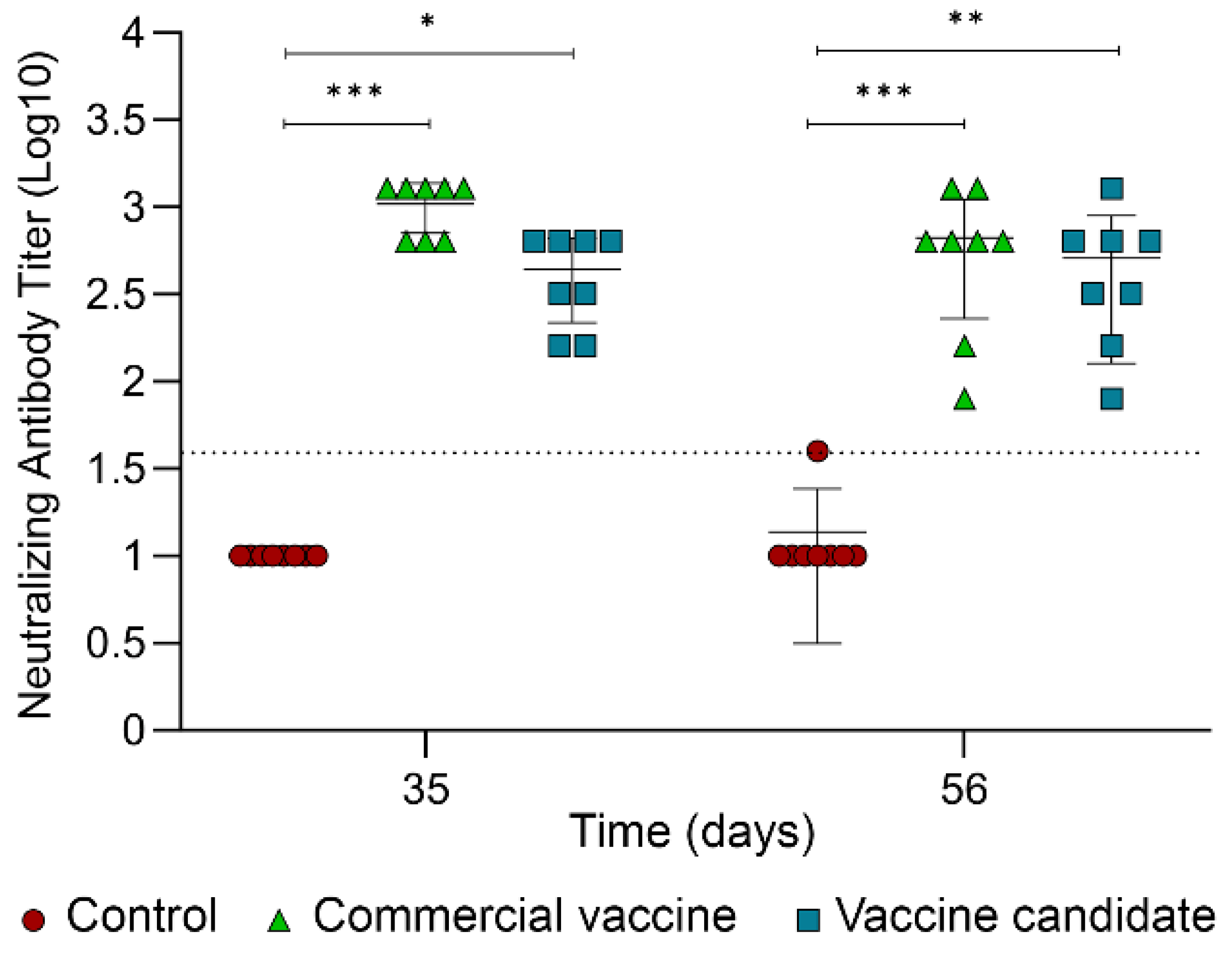
2.5. Determination of Neutralizing Antibodies
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Animals
4.2. BVDV Subgenotype Screening
4.3. Generation of CHO-K1 Clones Expressing Different E2 Proteins
4.4. Purification of E2 Proteins
4.5. SDS-PAGE and Western-Blot
4.6. Vaccine Formulation
4.7. Evaluation of the Vaccine Candidate in Sheep
4.8. Evaluation of the Vaccine Candidate in Bovines
4.9. Evaluation of Humoral and Cellular Immune Responses
4.9.1. Humoral Immune Response
4.9.2. Isolation of Peripheral Blood Mononuclear Cells
4.9.3. Stimulation of Peripheral Blood Mononuclear Cells
4.9.4. Evaluation of Cellular Response Markers Using RT-qPCR
4.9.5. ELISA for Detecting IFN-γ from PBMCs
4.9.6. Neutralization Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyal, S.M.; Ridpath, J.F. Bovine Viral Diarrhea Virus: Diagnosis, Management and Control; Blackwell Publishing Professional: Ames, IA, USA, 2005. [Google Scholar]
- Giammarioli, M.; Ceglie, L.; Rossi, E.; Bazzucchi, M.; Casciari, C.; Petrini, S.; De Mia, G.M. Increased Genetic Diversity of Bvdv-1: Recent Findings and Implications Thereof. Virus Genes 2015, 50, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Yesilbag, K.; Alpay, G.; Becher, P. Variability and Global Distribution of Subgenotypes of Bovine Viral Diarrhea Virus. Viruses 2017, 9, 128. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Chen, N.; Guidarini, C.; Xu, Z.; Zhang, J.; Cai, L.; Yuan, S.; Sun, Y.; Metcalfe, L. Prevalence and Genetic Diversity of Bovine Viral Diarrhea Virus in Dairy Herds of China. Vet. Microbiol. 2020, 242, 108565. [Google Scholar] [CrossRef] [PubMed]
- Santman-Berends, I.; Mars, M.H.; Van Duijn, L.; Van den Broek, K.W.H.; Van Schaik, G. A Quantitative Risk-Analysis for Introduction of Bovine Viral Diarrhoea Virus in the Netherlands through Cattle Imports. Prev. Vet. Med. 2017, 146, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Kelling, C.L. Evolution of Bovine Viral Diarrhea Virus Vaccines. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Bolin, S.R.; Ridpath, J.F. Specificity of Neutralizing and Precipitating Antibodies Induced in Healthy Calves by Monovalent Modified-Live Bovine Viral Diarrhea Virus Vaccines. Am. J. Vet. Res. 1989, 50, 817–821. [Google Scholar] [PubMed]
- Cortese, V.S.; Whittaker, R.; Ellis, J.; Ridpath, J.F.; Bolin, S.R. Specificity and Duration of Neutralizing Antibodies Induced in Healthy Cattle after Administration of a Modified-Live Virus Vaccine against Bovine Viral Diarrhea. Am. J. Vet. Res. 1998, 59, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Fulton, R.W.; Burge, L.J. Bovine Viral Diarrhea Virus Types 1 and 2 Antibody Response in Calves Receiving Modified Live Virus or Inactivated Vaccines. Vaccine 2000, 19, 264–274. [Google Scholar] [CrossRef]
- McClurkin, A.W.; Littledike, E.T.; Cutlip, R.C.; Frank, G.H.; Coria, M.F.; Bolin, S.R. Production of Cattle Immunotolerant to Bovine Viral Diarrhea Virus. Can. J. Comp. Med. 1984, 48, 156–161. [Google Scholar] [PubMed]
- Ridpath, J.F. Immunology of Bvdv Vaccines. Biologicals 2013, 41, 14–19. [Google Scholar] [CrossRef]
- Falkenberg, S.M.; Dassanayake, R.P.; Crawford, L.; Sarlo Davila, K.; Boggiatto, P. Response to Bovine Viral Diarrhea Virus in Heifers Vaccinated with a Combination of Multivalent Modified Live and Inactivated Viral Vaccines. Viruses 2023, 15, 703. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Walz, P.H.; Givens, M.D.; Wilson, A.E. Efficacy of Bovine Viral Diarrhea Virus Vaccination to Prevent Reproductive Disease: A Meta-Analysis. Theriogenology 2015, 83, 360–365.e1. [Google Scholar] [CrossRef]
- Patel, J.R.; Shilleto, R.W.; Williams, J.; Alexander, D.C. Prevention of Transplacental Infection of Bovine Foetus by Bovine Viral Diarrhoea Virus through Vaccination. Arch. Virol. 2002, 147, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Antos, A.; Miroslaw, P.; Rola, J.; Polak, M.P. Vaccination Failure in Eradication and Control Programs for Bovine Viral Diarrhea Infection. Front. Vet. Sci. 2021, 8, 688911. [Google Scholar] [CrossRef] [PubMed]
- Newcomer, B.W.; Givens, D. Diagnosis and Control of Viral Diseases of Reproductive Importance: Infectious Bovine Rhinotracheitis and Bovine Viral Diarrhea. Vet. Clin. N. Am. Food Anim. Pract. 2016, 32, 425–441. [Google Scholar] [CrossRef]
- Brownlie, J.; Clarke, M.C.; Hooper, L.B.; Bell, G.D. Protection of the Bovine Fetus from Bovine Viral Diarrhoea Virus by Means of a New Inactivated Vaccine. Vet. Rec. 1995, 137, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Al-Kubati, A.A.G.; Hussen, J.; Kandeel, M.; Al-Mubarak, A.I.A.; Hemida, M.G. Recent Advances on the Bovine Viral Diarrhea Virus Molecular Pathogenesis, Immune Response, and Vaccines Development. Front. Vet. Sci. 2021, 8, 665128. [Google Scholar] [CrossRef]
- Laureyns, J.; Ribbens, S.; de Kruif, A. Control of Bovine Virus Diarrhoea at the Herd Level: Reducing the Risk of False Negatives in the Detection of Persistently Infected Cattle. Vet. J. 2010, 184, 21–26. [Google Scholar] [CrossRef]
- Aitkenhead, H.; Riedel, C.; Cowieson, N.; Rumenapf, H.T.; Stuart, D.I.; El Omari, K. Structural Comparison of Typical and Atypical E2 Pestivirus Glycoproteins. Structure 2024, 32, 273–281.E4. [Google Scholar] [CrossRef]
- Donofrio, G.; Bottarelli, E.; Sandro, C.; Flammini, C.F. Expression of Bovine Viral Diarrhea Virus Glycoprotein E2 as a Soluble Secreted form in a Mammalian Cell Line. Clin. Vaccine Immunol. 2006, 13, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Nogarol, C.; Decaro, N.; Bertolotti, L.; Colitti, B.; Iotti, B.; Petrini, S.; Lucente, M.S.; Elia, G.; Perona, G.; Profiti, M.; et al. Pestivirus Infection in Cattle Dairy Farms: E2 Glycoprotein Elisa Reveals the Presence of Bovine Viral Diarrhea Virus Type 2 in Northwestern Italy. BMC Vet. Res. 2017, 13, 377. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, B.; Letellier, C.; Olivier, F.; Dehan, P.; Elouahabi, A.; Vandenbranden, M.; Ruysschaert, J.M.; Hamers, C.; Pastoret, P.P.; Kerkhofs, P. Sequence-Optimised E2 Constructs from Bvdv-1b and Bvdv-2 for DNA Immunisation in Cattle. Vet. Res. 2007, 38, 819–834. [Google Scholar] [CrossRef]
- van Drunen Littel-van den Hurk, S.; Lawman, Z.; Wilson, D.; Luxembourg, A.; Ellefsen, B.; van den Hurk, J.V.; Hannaman, D. Electroporation Enhances Immune Responses and Protection Induced by a Bovine Viral Diarrhea Virus DNA Vaccine in Newborn Calves with Maternal Antibodies. Vaccine 2010, 28, 6445–6454. [Google Scholar] [CrossRef] [PubMed]
- El-Attar, L.M.R.; Thomas, C.; Luke, J.; Williams, J.A.; Brownlie, J. Enhanced Neutralising Antibody Response to Bovine Viral Diarrhoea Virus (Bvdv) Induced by DNA Vaccination in Calves. Vaccine 2015, 33, 4004–4012. [Google Scholar] [CrossRef] [PubMed]
- Leng, D.; Yamada, S.; Chiba, Y.; Yoneyama, S.; Sakai, Y.; Hikono, H.; Murakami, K. Co-Administration of a Plasmid Encoding Cd40 or Cd63 Enhances the Immune Responses to a DNA Vaccine against Bovine Viral Diarrhea Virus in Mice. J. Vet. Med. Sci. 2022, 84, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Cheng, L.T.; Zhang, J.Y.; Wu, Y.J.; Liu, S.S.; Chu, C.Y. Recombinant E2 Protein Enhances Protective Efficacy of Inactivated Bovine Viral Diarrhea Virus 2 Vaccine in a Goat Model. BMC Vet. Res. 2018, 14, 194. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Young, N.J.; Heaney, J.; Collins, M.E.; Brownlie, J. Evaluation of Efficacy of Mammalian and Baculovirus Expressed E2 Subunit Vaccine Candidates to Bovine Viral Diarrhoea Virus. Vaccine 2009, 27, 2387–2393. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Marconi, P.; Periolo, O.; La Torre, J.; Alvarez, M.A. Immunocompetent Truncated E2 Glycoprotein of Bovine Viral Diarrhea Virus (Bvdv) Expressed in Nicotiana Tabacum Plants: A Candidate Antigen for New Generation of Veterinary Vaccines. Vaccine 2012, 30, 4499–4504. [Google Scholar] [CrossRef]
- Pecora, A.; Malacari, D.A.; Perez Aguirreburualde, M.S.; Bellido, D.; Nunez, M.C.; Dus Santos, M.J.; Escribano, J.M.; Wigdorovitz, A. Development of an Apc-Targeted Multivalent E2-Based Vaccine against Bovine Viral Diarrhea Virus Types 1 and 2. Vaccine 2015, 33, 5163–5171. [Google Scholar] [CrossRef]
- Lo, Y.T.; Ryan, M.D.; Luke, G.A.; Chang, W.C.; Wu, H.C. Immunogenicity of a Secreted, C-Terminally Truncated, Form of Bovine Viral Diarrhea Virus E2 Glycoprotein as a Potential Candidate in Subunit Vaccine Development. Sci. Rep. 2023, 13, 296. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, B.; Niu, C.; Jia, S.; Sun, C.; Wang, Z.; Jiang, Y.; Cui, W.; Wang, L.; Xu, Y. Dendritic Cell Targeting of Bovine Viral Diarrhea Virus E2 Protein Expressed by Lactobacillus Casei Effectively Induces Antigen-Specific Immune Responses via Oral Vaccination. Viruses 2019, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Montesino, R.; Maura, R.; Hugues, F.; Cabezas, O.; Altamirano, C.; Sánchez, O.; Toledo, J.R. Characterisation of a New Molecule Based on Two E2 Sequences from Bovine Viral Diarrhoea-Mucosal Disease Virus Fused to the Human Immunoglobulin Fc Fragment. J. Vet. Res. 2021, 65, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Hugues, F.; Cabezas, I.; Garigliany, M.; Rivas, F.; Casanova, T.; Gonzalez, E.E.; Sanchez, O.; Castillo, R.; Parra, N.C.; Inostroza-Michael, O.; et al. First Report of Bovine Viral Diarrhea Virus Subgenotypes 1d and 1e in Southern Chile. Virol. J. 2023, 20, 205. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Lucero, J.; Celedon, M.O.; Aguilera, M.; de Calisto, A. Molecular Characterization of Pestiviruses Isolated from Bovines in Chile. Vet. Microbiol. 2006, 115, 208–217. [Google Scholar] [CrossRef]
- Fernandez, F.; Costantini, V.; Barrandeguy, M.; Parreno, V.; Schiappacassi, G.; Maliandi, F.; Leunda, M.; Odeon, A. Evaluation of Experimental Vaccines for Bovine Viral Diarrhea in Bovines, Ovines and Guinea Pigs. Rev. Argent. Microbiol. 2009, 41, 86–91. [Google Scholar] [PubMed]
- Abdisa, T. Review on Practical Guidance of Veterinary Clinical Diagnostic Approach. Int. J. Vet. Sci. Res. 2017, 3, 030–049. [Google Scholar] [CrossRef]
- Reece, W.O.; Erickson, H.H.; Goff, J.P.; Uemura, E.E. (Eds.) Dukes’ Physiology of Domestic Animals, 13th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2015. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Harpin, S.; Hurley, D.J.; Mbikay, M.; Talbot, B.; Elazhary, Y. Vaccination of Cattle with a DNA Plasmid Encoding the Bovine Viral Diarrhoea Virus Major Glycoprotein E2. J. Gen. Virol. 1999, 80 Pt 12, 3137–3144. [Google Scholar] [CrossRef]
- Liang, R.; Babiuk, L.A.; Hurk, S.v.D.L.-V.D. Compatibility of Plasmids Encoding Bovine Viral Diarrhea Virus Type 1 and Type 2 E2 in a Single DNA Vaccine Formulation. Vaccine 2007, 25, 5994–6006. [Google Scholar] [CrossRef]
- Nobiron, I.; Thompson, I.; Brownlie, J.; Collins, M.E. Cytokine Adjuvancy of Bvdv DNA Vaccine Enhances both Humoral and Cellular Immune Responses in Mice. Vaccine 2001, 19, 4226–4235. [Google Scholar] [CrossRef]
- Hurk, S.v.D.L.-V.D.; Lawman, Z.; Snider, M.; Wilson, D.; Hurk, J.V.v.D.; Ellefsen, B.; Hannaman, D. Two Doses of Bovine Viral Diarrhea Virus DNA Vaccine Delivered by Electroporation Induce Long-Term Protective Immune Responses. Clin. Vaccine Immunol. 2013, 20, 166–173. [Google Scholar] [CrossRef]
- Liang, R.; van den Hurk, J.V.; Landi, A.; Lawman, Z.; Deregt, D.; Townsend, H.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. DNA Prime Protein Boost Strategies Protect Cattle from Bovine Viral Diarrhea Virus Type 2 Challenge. J. Gen. Virol. 2008, 89, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Song, Q.; Duan, C.; Wang, S.; Wang, J.; Zhu, Y. Enhanced Immune Responses to E2 Protein and DNA Formulated with Isa 61 Vg Administered as a DNA Prime-Protein Boost Regimen against Bovine Viral Diarrhea Virus. Vaccine 2018, 36, 5591–5599. [Google Scholar] [CrossRef] [PubMed]
- Perez Aguirreburualde, M.S.; Gomez, M.C.; Ostachuk, A.; Wolman, F.; Albanesi, G.; Pecora, A.; Odeon, A.; Ardila, F.; Escribano, J.M.; Dus Santos, M.J.; et al. Efficacy of a Bvdv Subunit Vaccine Produced in Alfalfa Transgenic Plants. Vet. Immunol. Immunopathol. 2013, 151, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhang, J.; Xu, M.; Yi, J.; Wang, Z.; Wang, Y.; Chen, C. Virus-Like Particles Vaccines Based on Glycoprotein E0 and E2 of Bovine Viral Diarrhea Virus Induce Humoral Responses. Front. Microbiol. 2022, 13, 1047001. [Google Scholar] [CrossRef] [PubMed]
- Katsura, M.; Fukushima, M.; Kameyama, K.I.; Kokuho, T.; Nakahira, Y.; Takeuchi, K. Novel Bovine Viral Diarrhea Virus (Bvdv) Virus-Like Particle Vaccine Candidates Presenting the E2 Protein Using the Spytag/Spycatcher System Induce a Robust Neutralizing Antibody Response in Mice. Arch. Virol. 2023, 168, 49. [Google Scholar] [CrossRef] [PubMed]
- Bruschke, C.J.; Moormann, R.J.; van Oirschot, J.T.; van Rijn, P.A. A Subunit Vaccine Based on Glycoprotein E2 of Bovine Virus Diarrhea Virus Induces Fetal Protection in Sheep against Homologous Challenge. Vaccine 1997, 15, 1940–1945. [Google Scholar] [CrossRef] [PubMed]
- Bruschke, C.J.; van Oirschot, J.T.; van Rijn, P.A. An Experimental Multivalent Bovine Virus Diarrhea Virus E2 Subunit Vaccine and Two Experimental Conventionally Inactivated Vaccines Induce Partial Fetal Protection in Sheep. Vaccine 1999, 17, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.I.; Pannhorst, K.; Sangewar, N.; Pavulraj, S.; Wen, X.; Stout, R.W.; Mwangi, W.; Paulsen, D.B. Bohv-1-Vectored Bvdv-2 Subunit Vaccine Induces Bvdv Cross-Reactive Cellular Immune Responses and Protects against Bvdv-2 Challenge. Vaccines 2021, 9, 46. [Google Scholar] [CrossRef]
- Pecora, A.; Aguirreburualde, M.S.; Aguirreburualde, A.; Leunda, M.R.; Odeon, A.; Chiavenna, S.; Bochoeyer, D.; Spitteler, M.; Filippi, J.L.; Dus Santos, M.J.; et al. Safety and Efficacy of an E2 Glycoprotein Subunit Vaccine Produced in Mammalian Cells to Prevent Experimental Infection with Bovine Viral Diarrhoea Virus in Cattle. Vet. Res. Commun. 2012, 36, 157–164. [Google Scholar] [CrossRef]
- Gomez-Romero, N.; Arias, C.F.; Verdugo-Rodriguez, A.; Lopez, S.; Valenzuela-Moreno, L.F.; Cedillo-Pelaez, C.; Basurto-Alcantara, F.J. Immune Protection Induced by E2 Recombinant Glycoprotein of Bovine Viral Diarrhea Virus in a Murine Model. Front. Vet. Sci. 2023, 10, 1168846. [Google Scholar] [CrossRef]
- Cheng, Y.; Tu, S.; Chen, T.; Zou, J.; Wang, S.; Jiang, M.; Tian, S.; Guo, Q.; Suolang, S.; Zhou, H. Evaluation of the Mucosal Immunity Effect of Bovine Viral Diarrhea Virus Subunit Vaccine E2fc and E2ft. Int. J. Mol. Sci. 2023, 24, 4172. [Google Scholar] [CrossRef] [PubMed]
- Mosena, A.C.S.; Falkenberg, S.M.; Ma, H.; Casas, E.; Dassanayake, R.P.; Walz, P.H.; Canal, C.W.; Neill, J.D. Multivariate Analysis as a Method to Evaluate Antigenic Relationships between Bvdv Vaccine and Field Strains. Vaccine 2020, 38, 5764–5772. [Google Scholar] [CrossRef] [PubMed]
- Vega, S.; Rosell, R.; Paton, D.J.; Orden, J.A.; de la Fuente, R. Antigenic Characterization of Bovine Viral Diarrhoea Virus Isolates from Spain with a Panel of Monoclonal Antibodies. J. Vet. Med. B Infect. Dis. Vet. Public. Health 2000, 47, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.; Avalos Ramirez, R.; Orlich, M.; Cedillo Rosales, S.; Konig, M.; Schweizer, M.; Stalder, H.; Schirrmeier, H.; Thiel, H.J. Genetic and Antigenic Characterization of Novel Pestivirus Genotypes: Implications for Classification. Virology 2003, 311, 96–104. [Google Scholar] [CrossRef]
- Klimowicz-Bodys, M.D.; Polak, M.P.; Ploneczka-Janeczko, K.; Bagnicka, E.; Zbroja, D.; Rypula, K. Lack of Fetal Protection against Bovine Viral Diarrhea Virus in a Vaccinated Heifer. Viruses 2022, 14, 311. [Google Scholar] [CrossRef]
- Fulton, R.W.; Ridpath, J.F.; Confer, A.W.; Saliki, J.T.; Burge, L.J.; Payton, M.E. Bovine Viral Diarrhoea Virus Antigenic Diversity: Impact on Disease and Vaccination Programmes. Biologicals 2003, 31, 89–95. [Google Scholar] [CrossRef]
- Van Campen, H.; Vorpahl, P.; Huzurbazar, S.; Edwards, J.; Cavender, J. A case report: Evidence for Type 2 Bovine Viral Diarrhea Virus (Bvdv)-Associated Disease in Beef Herds Vaccinated with a Modified-Live Type 1 Bvdv Vaccine. J. Vet. Diagn. Investig. 2000, 12, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Downey-Slinker, E.D.; Ridpath, J.F.; Sawyer, J.E.; Skow, L.C.; Herring, A.D. Antibody Titers to Vaccination Are Not Predictive of Level of Protection against a Bvdv Type 1b Challenge in Bos Indicus-Bos Taurus Steers. Vaccine 2016, 34, 5053–5059. [Google Scholar] [CrossRef]
- Beer, M.; Hehnen, H.R.; Wolfmeyer, A.; Poll, G.; Kaaden, O.R.; Wolf, G. A New Inactivated Bvdv Genotype I and II Vaccine. An Immunisation and Challenge Study with Bvdv Genotype I. Vet. Microbiol. 2000, 77, 195–208. [Google Scholar] [CrossRef]
- Bolin, S.R.; Ridpath, J.F. Assessment of Protection from Systemic Infection or Disease Afforded by Low to Intermediate Titers of Passively Acquired Neutralizing Antibody against Bovine Viral Diarrhea Virus in Calves. Am. J. Vet. Res. 1995, 56, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.F. Practical significance of heterogeneity among Bvdv strains: Impact of Biotype and Genotype on U.S. Control Programs. Prev. Vet. Med. 2005, 72, 17–30; discussion 215–219. [Google Scholar] [CrossRef] [PubMed]
- Platt, R.; Widel, P.W.; Kesl, L.D.; Roth, J.A. Comparison of Humoral and Cellular Immune Responses to a Pentavalent Modified Live Virus Vaccine in Three Age Groups of Calves with Maternal Antibodies, before and after Bvdv Type 2 Challenge. Vaccine 2009, 27, 4508–4519. [Google Scholar] [CrossRef] [PubMed]
- Vilcek, S.; Herring, A.J.; Herring, J.A.; Nettleton, P.F.; Lowings, J.P.; Paton, D.J. Pestiviruses Isolated from Pigs, Cattle and Sheep Can Be Allocated into at Least Three Genogroups Using Polymerase Chain Reaction and Restriction Endonuclease Analysis. Arch. Virol. 1994, 136, 309–323. [Google Scholar] [CrossRef]
- Birnboim, H.C.; Doly, J. A Rapid Alkaline Extraction Procedure for Screening Recombinant Plasmid DNA. Nucleic Acids Res. 1979, 7, 1513–1523. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Number of Samples | Ab. Positive | Ab. Negative | Ab. Doubtful | Ag. Positive | Seq. | Viral Subtype |
|---|---|---|---|---|---|---|---|
| Maule | 21 | 0 | 21 | 0 | 21 | 7 | BVDV1b |
| Ñuble | 238 | 122 | 110 | 6 | 7 | 7 | BVDV1b (6), BVDV1d (1) |
| Bio-Bio | 2 | 0 | 2 | 0 | 2 | 0 | - |
| Araucanía | 15 | 0 | 15 | 0 | 15 | 3 | BVDV1b |
| Los Ríos | 58 | 0 | 58 | 0 | 58 | 7 | BVDV1b (6), BVDV1d (1) |
| Los Lagos | 923 | 877 | 38 | 8 | 26 | 7 | BVDV1b (3), BVDV1d (4) |
| Aysén | 695 | 637 | 55 | 3 | 0 | 0 | - |
| Magallanes | 3 | 0 | 3 | 0 | 3 | 0 | - |
| Total | 1955 | 1636 | 302 | 17 | 132 | 31 |
| Genotypes | Anti-Tag Antibody | Secondary Antibody | Anti-6xHis Antibody | Secondary Antibody |
|---|---|---|---|---|
| 1aE2-cMyc-6xHis | Mouse anti-c-Myc tag, monoclonal antibody (cod. 13-2500, Invitrogen, Waltham, MA, USA). 1/1000 | Goat anti-Mouse IgG (H+L) antibody, Alexa Fluor™ 790 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 | Rabbit anti-6xHis tag polyclonal antibody (PA1-983B, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Rabbit IgG (H+L) antibody Alexa Fluor® 680 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 |
| 1bE2-HA-6xHis | Mouse anti-HA tag monoclonal antibody (cod. 26183, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Mouse IgG (H+L) antibody, Alexa Fluor™ 790 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 | Rabbit anti-6xHis tag polyclonal antibody (PA1-983B, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Rabbit IgG (H+L) antibody Alexa Fluor® 680 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 |
| 1cE2-VSV-6xHis | Rabbit anti-VSV-G tag polyclonal antibody (cod. A190-131A. Bethyl laboratories. Inc, Montgomery, TX, USA) 1/5000 | Goat anti-Rabbit IgG (H+L) antibody Alexa Fluor® 680 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 | Mouse anti-6xHis tag monoclonal antibody (cod. 631212, Clontech Laboratories, Waltham, MA, USA 1/5000 | Goat anti-Mouse IgG (H+L) antibody, Alexa Fluor™ 790 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 |
| 1dE2-V5-6xHis | Mouse anti-V5 tag monoclonal antibody (cod. R960-25, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Mouse IgG (H+L) antibody, Alexa Fluor™ 790 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 | Rabbit anti-6xHis tag polyclonal antibody (PA1-983B, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Rabbit IgG (H+L) antibody Alexa Fluor® 680 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 |
| 1eE2-E-6xHis | Goat anti-E tag Epitope Tag polyclonal antibody (NB600-518B, Novus biologicals, Centennial, CO, USA) 1/5000 | Rabbit anti-Goat IgG (H+L) antibody, Alexa Fluor™ 790 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 | Rabbit anti-6xHis tag polyclonal antibody (PA1-983B, Invitrogen, Waltham, MA, USA) 1/5000 | Goat anti-Rabbit IgG (H+L) antibody Alexa Fluor® 680 (Thermo Fischer Scientific, Waltham, MA, USA) 1/30,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avello, V.; Salazar, S.; González, E.E.; Campos, P.; Manríque, V.; Mathieu, C.; Hugues, F.; Cabezas, I.; Gädicke, P.; Parra, N.C.; et al. Recombinant Subunit Vaccine Candidate against the Bovine Viral Diarrhea Virus. Int. J. Mol. Sci. 2024, 25, 8734. https://doi.org/10.3390/ijms25168734
Avello V, Salazar S, González EE, Campos P, Manríque V, Mathieu C, Hugues F, Cabezas I, Gädicke P, Parra NC, et al. Recombinant Subunit Vaccine Candidate against the Bovine Viral Diarrhea Virus. International Journal of Molecular Sciences. 2024; 25(16):8734. https://doi.org/10.3390/ijms25168734
Chicago/Turabian StyleAvello, Verónica, Santiago Salazar, Eddy E. González, Paula Campos, Viana Manríque, Christian Mathieu, Florence Hugues, Ignacio Cabezas, Paula Gädicke, Natalie C. Parra, and et al. 2024. "Recombinant Subunit Vaccine Candidate against the Bovine Viral Diarrhea Virus" International Journal of Molecular Sciences 25, no. 16: 8734. https://doi.org/10.3390/ijms25168734