
Quercetin Attenuates Acetaldehyde-Induced Cytotoxicity via the Heme Oxygenase-1-Dependent Antioxidant Mechanism in Hepatocytes

 , and
, and
Abstract
1. Introduction
2. Results
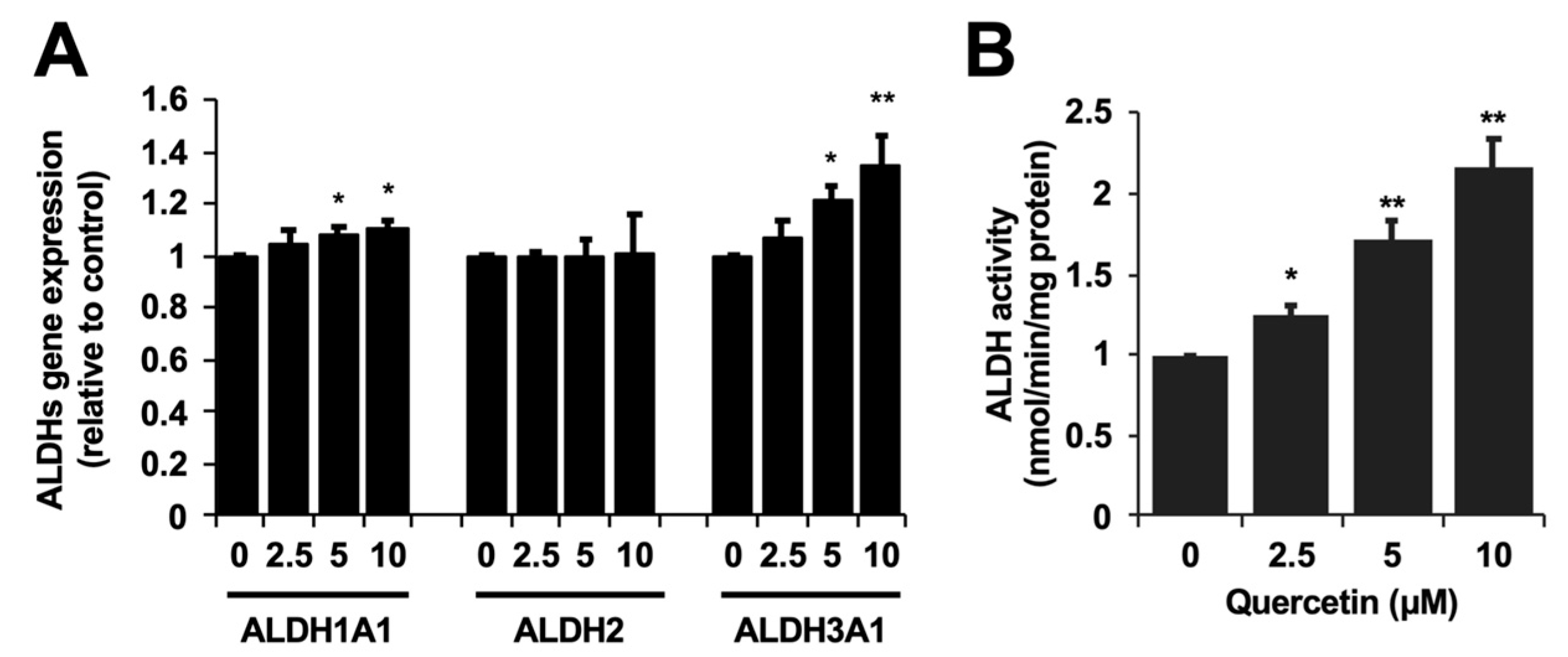
2.1. Cytoprotection against the Acetaldehyde Toxicity and ALDH Activity Enhancement by Quercetin
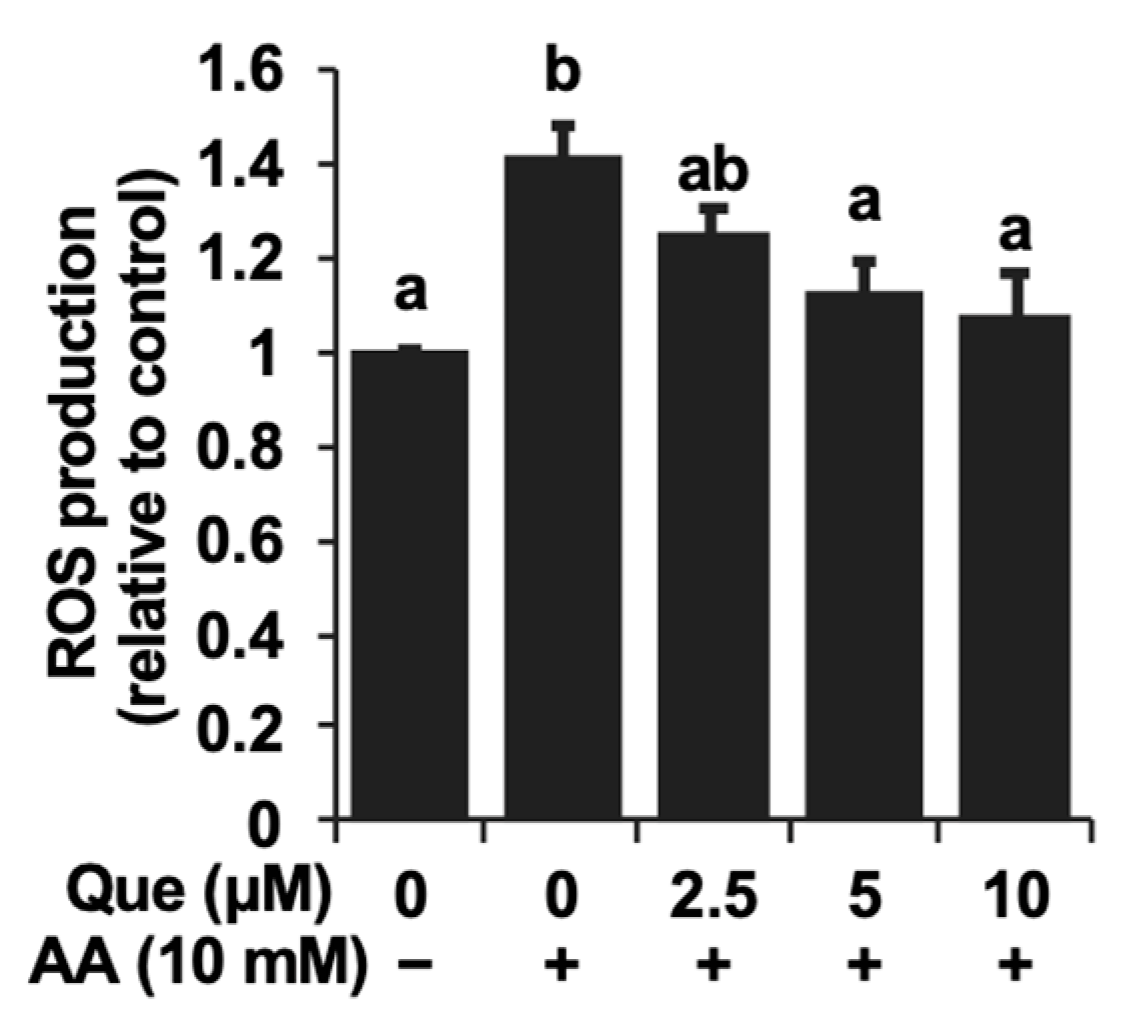
2.2. Inhibitory Effect of Quercetin on the Reactive Oxygen Species Accumulation Induced by Acetaldehyde
2.3. Up-Regulation of the Intracellular Antioxidant-Related Gene Expression by Quercetin
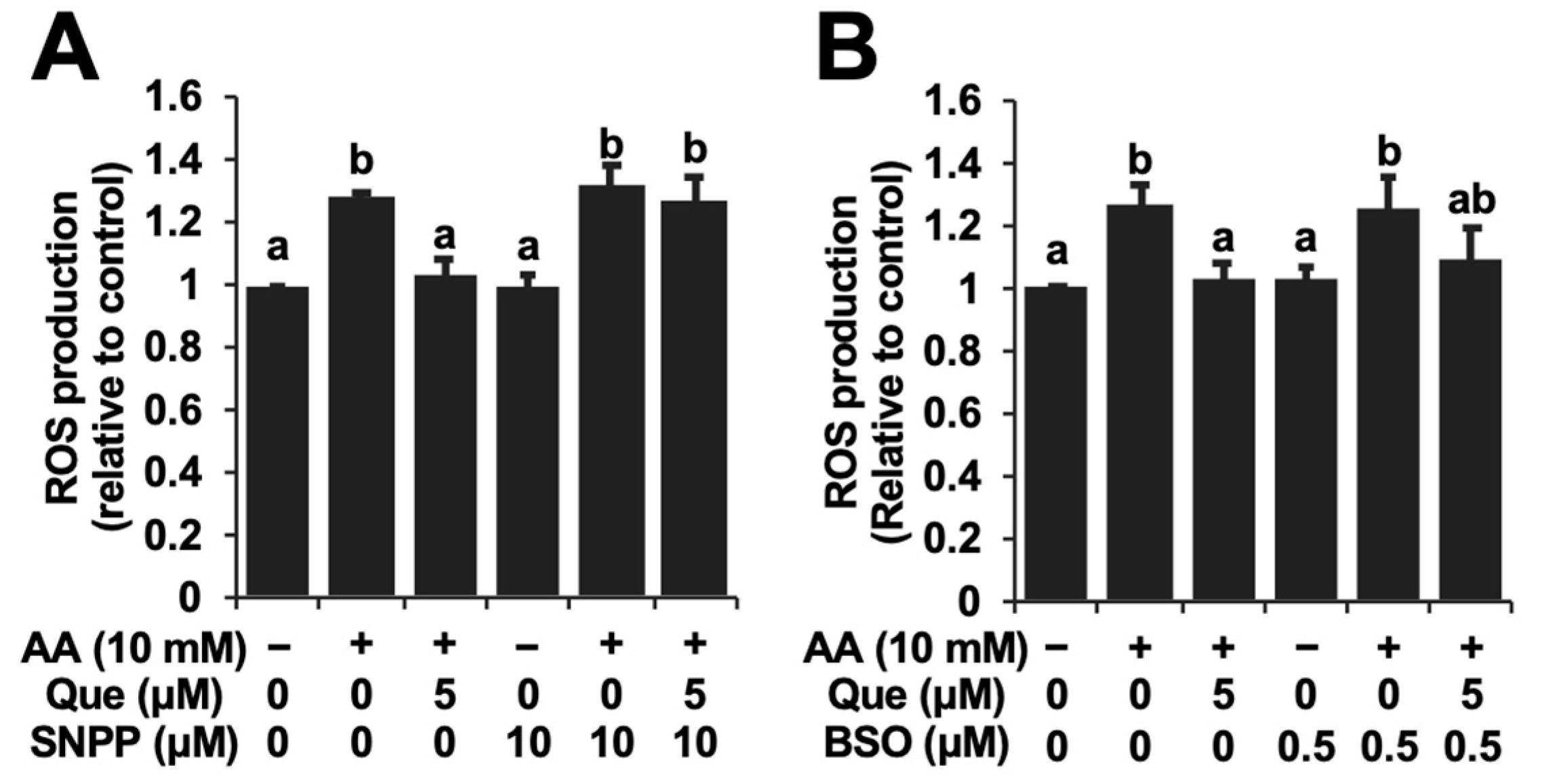
2.4. HO-1-Denpedent Inhibition of the Acetaldehyde-Induced ROS Elevation and Cytotoxicity by Quercetin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures and Viability Determination
4.3. Intracellular Reactive Oxygen Species Measurement and Image Analysis
4.4. RNA Extraction and Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.5. Glutathione Titration
4.6. ALDH Activity Assay
4.7. Western Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rocco, A.; Compare, D.; Angrisani, D.; Sanduzzi Zamparelli, M.; Nardone, G. Alcoholic disease: Liver and beyond. World J. Gastroenterol. 2014, 20, 14652–14659. [Google Scholar] [CrossRef] [PubMed]
- Ceni, E.; Mello, T.; Galli, A. Pathogenesis of alcoholic liver disease: Role of oxidative metabolism. World J. Gastroenterol. 2014, 20, 17756–17772. [Google Scholar] [CrossRef]
- Jackson, B.; Brocker, C.; Thompson, D.C.; Black, W.; Vasiliou, K.; Nebert, D.W.; Vasiliou, V. Update on the aldehyde dehydrogenase gene (ALDH) superfamily. Hum. Genom. 2011, 5, 283–303. [Google Scholar] [CrossRef]
- Lind, P.A.; Eriksson, C.J.; Wilhelmsen, K.C. The role of aldehyde dehydrogenase-1 (ALDH1A1) polymorphisms in harmful alcohol consumption in a Finnish population. Hum. Genom. 2008, 3, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, C.; Xu, H.; Gao, Y. Aldehyde Dehydrogenase, Liver Disease and Cancer. Int. J. Biol. Sci. 2020, 16, 921–934. [Google Scholar] [CrossRef]
- Kruk, J.; Aboul-Enein, H.Y.; Kładna, A.; Bowser, J.E. Oxidative stress in biological systems and its relation with pathophysi-ological functions: The effect of physical activity on cellular redox homeostasis. Free Radic. Res. 2019, 53, 497–521. [Google Scholar] [CrossRef]
- Tsermpini, E.E.; Plemenitaš Ilješ, A.; Dolžan, V. Alcohol-Induced Oxidative Stress and the Role of Antioxidants in Alcohol Use Disorder: A Systematic Review. Antioxidants 2022, 11, 1374. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Y.; Zhao, X.; Pang, J.; Pan, C.; Wang, J.; Wei, S.; Yu, X.; Zhang, C.; Chen, Y.; et al. The role of aldehyde dehydrogenase 2 in cardiovascular disease. Nat. Rev. Cardiol. 2023, 20, 495–509. [Google Scholar] [CrossRef]
- Wang, X.; Chang, X.; Zhan, H.; Zhang, Q.; Li, C.; Gao, Q.; Yang, M.; Luo, Z.; Li, S.; Sun, Y. Curcumin and Baicalin ameliorate ethanol-induced liver oxidative damage via the Nrf2/HO-1 pathway. J. Food Biochem. 2020, 44, e13425. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Ryter, S.W. Heme Oxygenase-1: An Anti-Inflammatory Effector in Cardiovascular, Lung, and Related Metabolic Disorders. Antioxidants 2022, 11, 555. [Google Scholar] [CrossRef] [PubMed]
- Haodang, L.; Lianmei, Q.; Ranhui, L.; Liesong, C.; Jun, H.; Yihua, Z.; Cuiming, Z.; Yimou, W.; Xiaoxing, Y. HO-1 mediates the anti-inflammatory actions of Sulforaphane in monocytes stimulated with a mycoplasmal lipopeptide. Chem. Biol. Interact. 2019, 306, 10–18. [Google Scholar] [CrossRef]
- Nakamura, Y.; Miyoshi, N. Electrophiles in foods: The current status of isothiocyanates and their chemical biology. Biosci. Biotechnol. Biochem. 2010, 74, 242–255. [Google Scholar] [CrossRef]
- Terao, J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants 2023, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Qi, W.; Xiong, D.; Long, M. Quercetin: Its Antioxidant Mechanism, Antibacterial Properties and Potential Application in Prevention and Control of Toxipathy. Molecules 2022, 27, 6545. [Google Scholar] [CrossRef]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhou, X.; Zhao, J. Quercetin prevents alcohol-induced liver injury through targeting of PI3K/Akt/nuclear factor-kappaB and STAT3 signaling pathway. Exp. Ther. Med. 2017, 14, 6169–6175. [Google Scholar]
- Jelski, W.; Szmitkowski, M. Alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) in the cancer diseases. Clin. Chim. Acta 2008, 395, 1–5. [Google Scholar] [CrossRef]
- Chen, C.H.; Cruz, L.A.; Mochly-Rosen, D. Pharmacological recruitment of aldehyde dehydrogenase 3A1 (ALDH3A1) to assist ALDH2 in acetaldehyde and ethanol metabolism in vivo. Proc. Natl. Acad. Sci. USA 2015, 112, 3074–3079. [Google Scholar] [CrossRef]
- Tamura, M.; Ito, H.; Matsui, H. Acetaldehyde is an oxidative stressor for gastric epithelial cells. J. Clin. Biochem. Nutr. 2014, 55, 26–31. [Google Scholar] [CrossRef]
- Wu, H.; Nakamura, T.; Guo, Y.; Matsumoto, R.; Munemasa, S.; Murata, Y.; Nakamura, Y. Cycloartenyl Ferulate Is the Predominant Compound in Brown Rice Conferring Cytoprotective Potential against Oxidative Stress-Induced Cytotoxicity. Int. J. Mol. Sci. 2023, 24, 822. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wu, H.; Kidawara, M.; Lin, Y.; Satoh, A.; Zhang, G.; Munemasa, S.; Murata, Y.; Nakamura, T.; Nakamura, Y. The microbiota catabolites of quercetin glycosides concertedly enhance the resistance against acetaldehyde-induced oxidative stress. Free Radic. Res. 2022, 56, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Kitson, T.M.; Kitson, K.E. The effect of quercetin, a widely distributed flavonoid in food and drink, on cytosolic aldehyde dehydrogenase: A comparison with the effect of diethylstilboestrol. Biochim. Biophys. Acta. 2000, 1481, 247–254. [Google Scholar] [CrossRef]
- Liu. Y.; Kurita, A.; Nakashima, S.; Zhu, B.; Munemasa, S.; Nakamura, T.; Murata, Y.; Nakamura, Y. 3,4-Dihydroxyphenylacetic acid is a potential aldehyde dehydrogenase inducer in murine hepatoma Hepa1c1c7 cells. Biosci. Biotechnol. Biochem. 2017, 81, 1978–1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Myojin, T.; Li, K.; Kurita, A.; Seto, M.; Motoyama, A.; Liu, X.; Satoh, A.; Munemasa, S.; Murata, Y.; et al. A Major Intestinal Catabolite of Quercetin Glycosides, 3-Hydroxyphenylacetic Acid, Protects the Hepatocytes from the Acetaldehyde-Induced Cytotoxicity through the Enhancement of the Total Aldehyde Dehydrogenase Activity. Int. J. Mol. Sci. 2022, 23, 1762. [Google Scholar] [CrossRef]
- Liu, Y.; Yamanaka, M.; Abe-Kanoh, N.; Liu, X.; Zhu, B.; Munemasa, S.; Nakamura, T.; Murata, Y.; Nakamura, Y. Benzyl isothiocyanate ameliorates acetaldehyde-induced cytotoxicity by enhancing aldehyde dehydrogenase activity in murine hepatoma Hepa1c1c7 cells. Food Chem. Toxicol. 2017, 108, 305–313. [Google Scholar] [CrossRef]
- Xu, Y.; Sawamoto, T.; Sun, R.; Ishikura, A.; Munemasa, S.; Murata, Y.; Satoh, A.; Matsumoto, A.; Nakamura, T.; Nakamura, Y. Evaluation of quercetin as a potential cytoprotector against acetaldehyde using the cultured hepatocyte model with aldehyde dehydrogenase isozyme deficiency. Biosci. Biotechnol. Biochem. 2024, zbae100. [Google Scholar] [CrossRef] [PubMed]
- Vrba, J.; Kren, V.; Vacek, J.; Papouskova, B.; Ulrichova, J. Quercetin, quercetin glycosides and taxifolin differ in their ability to induce AhR activation and CYP1A1 expression in HepG2 cells. Phytother. Res. 2012, 26, 1746–1752. [Google Scholar] [CrossRef]
- Yao, P.; Nussler, A.; Liu, L.; Hao, L.; Song, F.; Schirmeier, A.; Nussler, N. Quercetin protects human hepatocytes from ethanol-derived oxidative stress by inducing heme oxygenase-1 via the MAPK/Nrf2 pathways. J. Hepatol. 2007, 47, 253–261. [Google Scholar] [CrossRef]
- Rayamajhi, N.; Kim, S.K.; Go, H.; Joe, Y.; Callaway, Z.; Kang, J.G.; Ryter, S.W.; Chung, H.T. Quercetin induces mitochondrial biogenesis through activation of HO-1 in HepG2 cells. Oxid. Med. Cell Longev. 2013, 2013, 154279. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Nakashima, S.; Saiki, S.; Myoi, Y.; Abe, N.; Kuwazuru, S.; Zhu, B.; Ashida, H.; Murata, Y.; Nakamura, Y. 3,4-Dihydroxyphenylacetic acid is a predominant biologically-active catabolite of quercetin glycosides. Food Res. Int. 2016, 89, 716–723. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, F.; Echeverry, C.; Abin-Carriquiry, J.A.; Blasina, F.; Antúnez, K.; Jones, D.P.; Go, Y.M.; Liang, Y.L.; Dajas, F. After cellular internalization, quercetin causes Nrf2 nuclear translocation, increases glutathione levels, and prevents neuronal death against an oxidative insult. Free Radic. Biol. Med. 2010, 49, 738–747. [Google Scholar] [CrossRef]
- Averilla, J.N.; Oh, J.; Kim, J.S. Carbon Monoxide Partially Mediates Protective Effect of Resveratrol Against UVB-Induced Oxidative Stress in Human Keratinocytes. Antioxidants 2019, 8, 432. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, D.Y.; Kang, S.; Miao, W.; Kim, H.; Lee, Y.; Jon, S. Bilirubin nanoparticle preconditioning protects against hepatic ischemia-reperfusion injury. Biomaterials 2017, 133, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Keum, H.; Kim, T.W.; Kim, Y.; Seo, C.; Son, Y.; Kim, J.; Kim, D.; Jung, W.; Whang, C.H.; Jon, S. Bilirubin nanomedicine alleviates psoriatic skin inflammation by reducing oxidative stress and suppressing pathogenic signaling. J. Control Release 2020, 325, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Funes, S.C.; Rios, M.; Fernández-Fierro, A.; Covián, C.; Bueno, S.M.; Riedel, C.A.; Mackern-Oberti, J.P.; Kalergis, A.M. Naturally Derived Heme-Oxygenase 1 Inducers and Their Therapeutic Application to Immune-Mediated Diseases. Front. Immunol. 2020, 11, 1467. [Google Scholar] [CrossRef]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [CrossRef]
- Baker, M.A.; Cerniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Cycles and Product Size |
|---|---|---|---|
| mβ-actin | 5′-GTCACCCACACTGTGCCCATCTA-3′ | 5′-GCAATGCCAGGGTACATGGTGGT-3′ | 16, 455 bp |
| mALDH1A1 | 5′-GACAGGCTTTCCAGATTGGCTC-3′ | 5′-AAGACTTTCCCACCATTGAGTGC-3′ | 26, 142 bp |
| mALDH2 | 5′-TGAAGACGGTTACTGTCAAAGTGC-3′ | 5′-AGTGTGTGTGGCGGTTTTTCTC-3′ | 26, 115 bp |
| mALDH3 | 5′-GATGCCCATTGTGTGTGTTCG-3′ | 5′-CCACCGCTTGATGTCTCTGC-3′ | 26, 138 bp |
| mHO-1 | 5′-ACATCGACAGCCCCACCAAGTTCAA-3′ | 5′-CTGACGAAGTGACGCCATCTGTGAG-3′ | 22, 203 bp |
| mGCLC | 5′-GGCGATGTTCTTGAGACTCTGC-3′ | 5′-TTCCTTCGATCATGTAACTCCCATA-3′ | 26, 100 bp |
| mxCT | 5′-CCTGGCATTTGGACGCTACAT-3′ | 5′-TGAGAATTGCTGTGAGCTTGCA-3′ | 25, 182 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Kidawara, M.; Chen, Q.; Munemasa, S.; Murata, Y.; Nakamura, T.; Nakamura, Y. Quercetin Attenuates Acetaldehyde-Induced Cytotoxicity via the Heme Oxygenase-1-Dependent Antioxidant Mechanism in Hepatocytes. Int. J. Mol. Sci. 2024, 25, 9038. https://doi.org/10.3390/ijms25169038
Li K, Kidawara M, Chen Q, Munemasa S, Murata Y, Nakamura T, Nakamura Y. Quercetin Attenuates Acetaldehyde-Induced Cytotoxicity via the Heme Oxygenase-1-Dependent Antioxidant Mechanism in Hepatocytes. International Journal of Molecular Sciences. 2024; 25(16):9038. https://doi.org/10.3390/ijms25169038
Chicago/Turabian StyleLi, Kexin, Minori Kidawara, Qiguang Chen, Shintaro Munemasa, Yoshiyuki Murata, Toshiyuki Nakamura, and Yoshimasa Nakamura. 2024. "Quercetin Attenuates Acetaldehyde-Induced Cytotoxicity via the Heme Oxygenase-1-Dependent Antioxidant Mechanism in Hepatocytes" International Journal of Molecular Sciences 25, no. 16: 9038. https://doi.org/10.3390/ijms25169038
APA StyleLi, K., Kidawara, M., Chen, Q., Munemasa, S., Murata, Y., Nakamura, T., & Nakamura, Y. (2024). Quercetin Attenuates Acetaldehyde-Induced Cytotoxicity via the Heme Oxygenase-1-Dependent Antioxidant Mechanism in Hepatocytes. International Journal of Molecular Sciences, 25(16), 9038. https://doi.org/10.3390/ijms25169038