
Multiple Posterior Insula Projections to the Brainstem Descending Pain Modulatory System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Injection Sites
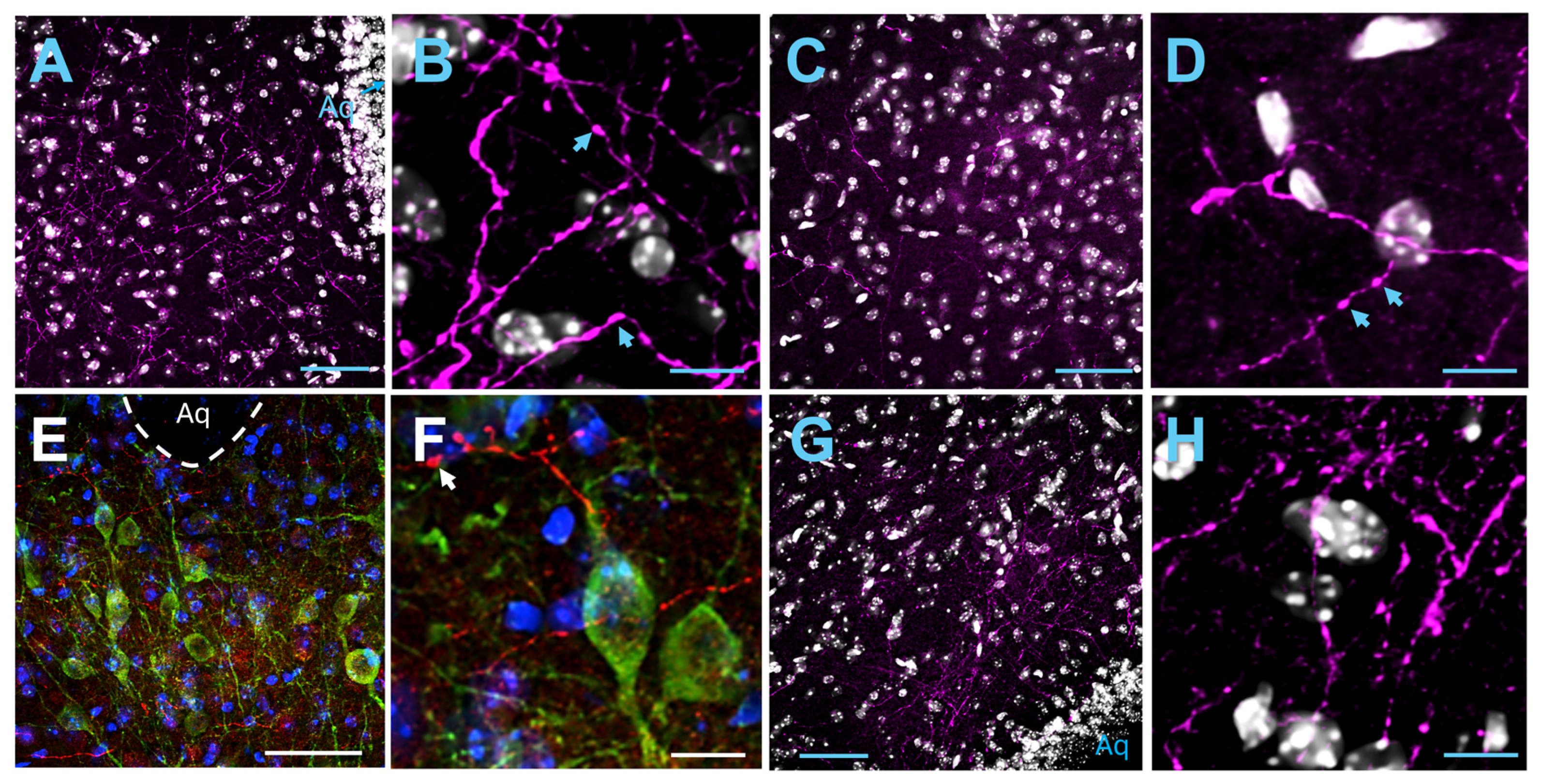
2.2. Axonal Projections from the Insula to the Brain Stem
2.2.1. Periaqueductal Gray
2.2.2. Raphe Magnus Nucleus
2.2.3. Locus Coeruleus and Other Noradrenergic Nuclei
2.2.4. Parabrachial Nucleus
2.2.5. Dorsal Horn of the Spinal Cord
2.3. Relative Projection Densities to the Different Brain Stem Areas
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Stereotactic Injections
4.3. Immunofluorescence
4.4. Imaging and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corder, G.; Castro, D.C.; Bruchas, M.R.; Scherrer, G. Endogenous and Exogenous Opioids in Pain. Annu. Rev. Neurosci. 2018, 41, 453–473. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M. Descending Facilitation. Mol. Pain 2017, 13, 174480691769921. [Google Scholar] [CrossRef] [PubMed]
- Porreca, F.; Ossipov, M.H.; Gebhart, G.F. Chronic Pain and Medullary Descending Facilitation. Trends Neurosci. 2002, 25, 319–325. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Fields, H.L. Endogenous Pain Control Systems: Brainstem Spinal Pathways and Endorphin Circuitry. Annu. Rev. Neurosci. 1984, 7, 309–338. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Fields, H.L. Endogenous Pain Control Mechanisms: Review and Hypothesis. Ann. Neurol. 1978, 4, 451–462. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Braz, J.; Ossipov, M.H.; Porreca, F. The Endogenous Neuromodulation System. In Neuromodulation; Elsevier: Amsterdam, The Netherlands, 2009; Volume 1, pp. 303–312. ISBN 9780123742483. [Google Scholar]
- Pertovaara, A. Noradrenergic Pain Modulation. Prog. Neurobiol. 2006, 80, 53–83. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.M.; Whittier, K.L.; Hegarty, D.M.; Aicher, S.A. Periaqueductal Gray Neurons Project to Spinally Projecting GABAergic Neurons in the Rostral Ventromedial Medulla. Pain 2008, 140, 376–386. [Google Scholar] [CrossRef]
- Lewis, V.A.; Gebhart, G.F. Evaluation of the Periaqueductal Central Gray (PAG) as a Morphine-Specific Locus of Action and Examination of Morphine-Induced and Stimulation-Produced Analgesia at Coincident PAG Loci. Brain Res. 1977, 124, 283–303. [Google Scholar] [CrossRef]
- Nguyen, E.; Grajales-Reyes, J.G.; Gereau, R.W.; Ross, S.E. Cell Type-Specific Dissection of Sensory Pathways Involved in Descending Modulation. Trends Neurosci. 2023, 46, 539–550. [Google Scholar] [CrossRef]
- Cheriyan, J.; Sheets, P.L. Altered Excitability and Local Connectivity of MPFC-PAG Neurons in a Mouse Model of Neuropathic Pain. J. Neurosci. 2018, 38, 4829–4839. [Google Scholar] [CrossRef]
- Huang, J.; Gadotti, V.M.; Chen, L.; Souza, I.A.; Huang, S.; Wang, D.; Ramakrishnan, C.; Deisseroth, K.; Zhang, Z.; Zamponi, G.W. A Neuronal Circuit for Activating Descending Modulation of Neuropathic Pain. Nat. Neurosci. 2019, 22, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.N.; Yousuf, H.; Dalton, S.; Sheets, P.L. Highly Differentiated Cellular and Circuit Properties of Infralimbic Pyramidal Neurons Projecting to the Periaqueductal Gray and Amygdala. Front. Cell. Neurosci. 2015, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Z.; Gambeta, E.; Chen, L.; Zamponi, G.W. An Orbitofrontal Cortex to Midbrain Projection Modulates Hypersensitivity after Peripheral Nerve Injury. Cell Rep. 2021, 35, 109033. [Google Scholar] [CrossRef] [PubMed]
- Siemian, J.N.; Arenivar, M.A.; Sarsfield, S.; Borja, C.B.; Erbaugh, L.J.; Eagle, A.L.; Robison, A.J.; Leinninger, G.; Aponte, Y. An Excitatory Lateral Hypothalamic Circuit Orchestrating Pain Behaviors in Mice. Elife 2021, 10, 1–31. [Google Scholar] [CrossRef]
- Siemian, J.N.; Borja, C.B.; Sarsfield, S.; Kisner, A.; Aponte, Y. Lateral Hypothalamic Fast-Spiking Parvalbumin Neurons Modulate Nociception through Connections in the Periaqueductal Gray Area. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Iwasaki, M.; Lefevre, A.; Althammer, F.; Clauss Creusot, E.; Łąpieś, O.; Petitjean, H.; Hilfiger, L.; Kerspern, D.; Melchior, M.; Küppers, S.; et al. An Analgesic Pathway from Parvocellular Oxytocin Neurons to the Periaqueductal Gray in Rats. Nat. Commun. 2023, 14, 1066. [Google Scholar] [CrossRef]
- Lee, J.-Y.; You, T.; Lee, C.-H.; Im, G.H.; Seo, H.; Woo, C.-W.; Kim, S.-G. Role of Anterior Cingulate Cortex Inputs to Periaqueductal Gray for Pain Avoidance. Curr. Biol. 2022, 32, 2834–2847.e5. [Google Scholar] [CrossRef]
- Zhu, X.; Xu, Y.; Shen, Z.; Zhang, H.; Xiao, S.; Zhu, Y.; Wu, M.; Chen, Y.; Wu, Z.; Xu, Y.; et al. Rostral Anterior Cingulate Cortex–Ventrolateral Periaqueductal Gray Circuit Underlies Electroacupuncture to Alleviate Hyperalgesia but Not Anxiety-Like Behaviors in Mice With Spared Nerve Injury. Front. Neurosci. 2022, 15, 757628. [Google Scholar] [CrossRef]
- Li, J.N.; Sheets, P.L. The Central Amygdala to Periaqueductal Gray Pathway Comprises Intrinsically Distinct Neurons Differentially Affected in a Model of Inflammatory Pain. J. Physiol. 2018, 596, 6289–6305. [Google Scholar] [CrossRef]
- Edwards, S.; Gilpin, N.W.; Baynes, B.B.; Middleton, J.W.; Avegno, E.M.; Whitaker, A.M.; Weera, M.M.; Lobell, T.D.; Itoga, C.A. Central Amygdala Circuits Mediate Hyperalgesia in Alcohol-Dependent Rats. J. Neurosci. 2018, 38, 7761–7773. [Google Scholar] [CrossRef]
- Wang, L.-H.; Ding, W.-Q.; Sun, Y.-G. Spinal Ascending Pathways for Somatosensory Information Processing. Trends Neurosci. 2022, 45, 594–607. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bleimeister, I.H.; Nguyen, E.K.; Li, J.; Cui, A.Y.; Stratton, H.J.; Smith, K.M.; Baccei, M.L.; Ross, S.E. The Functional and Anatomical Characterization of Three Spinal Output Pathways of the Anterolateral Tract. Cell Rep. 2024, 43, 113829. [Google Scholar] [CrossRef] [PubMed]
- Hermann, D.M.; Luppi, P.-H.; Peyron, C.; Hinckel, P.; Jouvet, M. Afferent Projections to the Rat Nuclei Raphe Magnus, Raphe Pallidus and Reticularis Gigantocellularis Pars α Demonstrated by Iontophoretic Application of Choleratoxin (Subunit B). J. Chem. Neuroanat. 1997, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Heinricher, M.M. Shifting the Balance: How Top-Down and Bottom-Up Input Modulate Pain via the Rostral Ventromedial Medulla. Front. Pain Res. 2022, 3, 932476. [Google Scholar] [CrossRef]
- Heinricher, M.M.; Tortorici, V. Interference with GABA Transmission in the Rostral Ventromedial Medulla: Disinhibition of off-Cells as a Central Mechanism in Nociceptive Modulation. Neuroscience 1994, 63, 533–546. [Google Scholar] [CrossRef]
- Neubert, M.J.; Kincaid, W.; Heinricher, M.M. Nociceptive Facilitating Neurons in the Rostral Ventromedial Medulla. Pain 2004, 110, 158–165. [Google Scholar] [CrossRef]
- Roeder, Z.; Chen, Q.; Davis, S.; Carlson, J.D.; Tupone, D.; Heinricher, M.M. Parabrachial Complex Links Pain Transmission to Descending Pain Modulation. Pain 2016, 157, 2697–2708. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Y.; Gao, P.; Pan, C.; Deng, H.; Xu, S.; Tang, D.; Xiao, J.; Jiao, Y.; Yu, W.; et al. The Locus Coeruleus Input to the Rostral Ventromedial Medulla Mediates Stress-Induced Colorectal Visceral Pain. Acta Neuropathol. Commun. 2023, 11, 65. [Google Scholar] [CrossRef]
- Jones, S.L.; Gebhart, G.F. Characterization of Coeruleospinal Inhibition of the Nociceptive Tail-Flick Reflex in the Rat: Mediation by Spinal A2-Adrenoceptors. Brain Res. 1986, 364, 315–330. [Google Scholar] [CrossRef]
- Hirschberg, S.; Li, Y.; Randall, A.; Kremer, E.J.; Pickering, A.E. Functional Dichotomy in Spinal- vs Prefrontal-Projecting Locus Coeruleus Modules Splits Descending Noradrenergic Analgesia from Ascending Aversion and Anxiety in Rats. Elife 2017, 6, 1–26. [Google Scholar] [CrossRef]
- Li, Y.; Hickey, L.; Perrins, R.; Werlen, E.; Patel, A.A.; Hirschberg, S.; Jones, M.W.; Salinas, S.; Kremer, E.J.; Pickering, A.E. Retrograde Optogenetic Characterization of the Pontospinal Module of the Locus Coeruleus with a Canine Adenoviral Vector. Brain Res. 2016, 1641, 274–290. [Google Scholar] [CrossRef]
- Lubejko, S.T.; Livrizzi, G.; Patel, J.; Yung, J.C.; Yaksh, T.L.; Banghart, M.R. Inputs to the Locus Coeruleus from the Periaqueductal Gray and Rostroventral Medulla Shape Opioid-Mediated Descending Pain Modulation. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Bajic, D.; Proudfit, H.K. Projections of Neurons in the Periaqueductal Gray to Pontine and Medullary Catecholamine Cell Groups Involved in the Modulation of Nociception. J. Comp. Neurol. 1999, 405, 359–379. [Google Scholar] [CrossRef]
- Barcomb, K.; Olah, S.S.; Kennedy, M.J.; Ford, C.P. Properties and Modulation of Excitatory Inputs to the Locus Coeruleus. J. Physiol. 2022, 600, 4897–4916. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.J.; Hervé-Minvielle, A. Inhibitory Influence of Frontal Cortex on Locus Coeruleus Neurons. Proc. Natl. Acad. Sci. USA 1995, 92, 6032–6036. [Google Scholar] [CrossRef] [PubMed]
- Labrakakis, C. The Role of the Insular Cortex in Pain. Int. J. Mol. Sci. 2023, 24, 5736. [Google Scholar] [CrossRef]
- Gogolla, N. The Insular Cortex. Curr. Biol. 2017, 27, R580–R586. [Google Scholar] [CrossRef]
- Segerdahl, A.R.; Mezue, M.; Okell, T.W.; Farrar, J.T.; Tracey, I. The Dorsal Posterior Insula Subserves a Fundamental Role in Human Pain. Nat. Neurosci. 2015, 18, 499–500. [Google Scholar] [CrossRef]
- Tan, L.L.; Pelzer, P.; Heinl, C.; Tang, W.; Gangadharan, V.; Flor, H.; Sprengel, R.; Kuner, T.; Kuner, R. A Pathway from Midcingulate Cortex to Posterior Insula Gates Nociceptive Hypersensitivity. Nat. Neurosci. 2017, 20, 1591–1601. [Google Scholar] [CrossRef]
- Sato, F.; Akhter, F.; Haque, T.; Kato, T.; Takeda, R.; Nagase, Y.; Sessle, B.J.J.; Yoshida, A. Projections from the Insular Cortex to Pain-Receptive Trigeminal Caudal Subnucleus (Medullary Dorsal Horn) and Other Lower Brainstem Areas in Rats. Neuroscience 2013, 233, 9–27. [Google Scholar] [CrossRef]
- Floyd, N.S.; Price, J.L.; Ferry, A.T.; Keay, K.A.; Bandler, R. Orbitomedial Prefrontal Cortical Projections to Distinct Longitudinal Columns of the Periaqueductal Gray in the Rat. J. Comp. Neurol. 2000, 422, 556–578. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, L.; Rabkin, S.D.; Granato, A.; Boudah, A.; Ohara, P.T. Analgesia and Hyperalgesia from GABA-Mediated Modulation of the Cerebral Cortex. Nature 2003, 424, 316–320. [Google Scholar] [CrossRef]
- Cedarbaum, J.M.; Aghajanian, G.K. Afferent Projections to the Rat Locus Coeruleus as Determined by a Retrograde Tracing Technique. J. Comp. Neurol. 1978, 178, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bruinstroop, E.; Cano, G.; Vanderhorst, V.G.J.M.; Cavalcante, J.C.; Wirth, J.; Sena-Esteves, M.; Saper, C.B. Spinal Projections of the A5, A6 (Locus Coeruleus), and A7 Noradrenergic Cell Groups in Rats. J. Comp. Neurol. 2012, 520, 1985–2001. [Google Scholar] [CrossRef] [PubMed]
- Kwiat, G.C.; Basbaum, A.I. The Origin of Brainstem Noradrenergic and Serotonergic Projections to the Spinal Cord Dorsal Horn in the Rat. Somatosens. Mot. Res. 1992, 9, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.E.; Pei, J.; Zhang, J.; Vlasov, K.Y.; Davis, T.; Taylor, E.; Wengvan Dort, F.J.J.; Solt, K.; Brown, E.N. The Role of Glutamatergic and Dopaminergic Neurons in the Periaqueductal Gray/Dorsal Raphe: Separating Analgesia and Anxiety. eNeuro 2019, 6, 1–14. [Google Scholar] [CrossRef]
- Hylden, J.L.K.; Anton, F.; Nahin, R.L. Spinal Lamina I Projection Neurons in the Rat: Collateral Innervation of Parabrachial Area and Thalamus. Neuroscience 1989, 28, 27–37. [Google Scholar] [CrossRef]
- Nakaya, Y.; Yamamoto, K.; Kobayashi, M. Descending Projections from the Insular Cortex to the Trigeminal Spinal Subnucleus Caudalis Facilitate Excitatory Outputs to the Parabrachial Nucleus in Rats. Pain 2023, 164, e157–e173. [Google Scholar] [CrossRef]
- Cai, Y.Q.; Wang, W.; Hou, Y.Y.; Pan, Z.Z. Optogenetic Activation of Brainstem Serotonergic Neurons Induces Persistent Pain Sensitization. Mol. Pain 2014, 10, 70. [Google Scholar] [CrossRef]
- Suárez-Pereira, I.; Llorca-Torralba, M.; Bravo, L.; Camarena-Delgado, C.; Soriano-Mas, C.; Berrocoso, E. The Role of the Locus Coeruleus in Pain and Associated Stress-Related Disorders. Biol. Psychiatry 2022, 91, 786–797. [Google Scholar] [CrossRef]
- Burnett, A.; Gebhart, G.F.F. Characterization of Descending Modulation of Nociception from the A5 Cell Group. Brain Res. 1991, 546, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, M.W.; Di Domenico, F.; Bannister, K. A Critical Brainstem Relay for Mediation of Diffuse Noxious Inhibitory Controls. Brain 2023, 146, 2259–2267. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, J.; Tsuruoka, M.; Maeda, M.; Hayashi, B.; Inoue, T. Involvement of a Descending Pathway from the A7 Region in Nociceptive Processing under Neuropathic Conditions in Rats. Dent. Med. Res. 2014, 34, 23–31. [Google Scholar] [CrossRef]
- Yeomans, D.C.; Clark, F.M.; Paice, J.A.; Proudfit, H.K. Antinociception Induced by Electrical Stimulation of Spinally Projecting Noradrenergic Neurons in the A7 Catecholamine Cell Group of the Rat. Pain 1992, 48, 449–461. [Google Scholar] [CrossRef]
- Gauriau, C.; Bernard, J. Pain Pathways and Parabrachial Circuits in the Rat. Exp. Physiol. 2002, 87, 251–258. [Google Scholar] [CrossRef]
- Chen, Q.; Roeder, Z.; Li, M.-H.; Zhang, Y.; Ingram, S.L.; Heinricher, M.M. Optogenetic Evidence for a Direct Circuit Linking Nociceptive Transmission through the Parabrachial Complex with Pain-Modulating Neurons of the Rostral Ventromedial Medulla (RVM). eNeuro 2017, 4, ENEURO.0202-17.2017. [Google Scholar] [CrossRef]
- Lapirot, O.; Chebbi, R.; Monconduit, L.; Artola, A.; Dallel, R.; Luccarini, P. NK1 Receptor-Expressing Spinoparabrachial Neurons Trigger Diffuse Noxious Inhibitory Controls through Lateral Parabrachial Activation in the Male Rat. Pain 2009, 142, 245–254. [Google Scholar] [CrossRef]
- Chiang, M.C.; Nguyen, E.K.; Canto-Bustos, M.; Papale, A.E.; Oswald, A.-M.M.M.; Ross, S.E.; Ross, S.E.; Canto-Bustos, M.; Papale, A.E.; Oswald, A.-M.M.M.; et al. Divergent Neural Pathways Emanating from the Lateral Parabrachial Nucleus Mediate Distinct Components of the Pain Response. Neuron 2020, 106, 927–939.e5. [Google Scholar] [CrossRef]
- Bouchatta, O.; Aby, F.; Sifeddine, W.; Bouali-Benazzouz, R.; Brochoire, L.; Manouze, H.; Fossat, P.; Ba M’Hamed, S.; Bennis, M.; Landry, M. Pain Hypersensitivity in a Pharmacological Mouse Model of Attention-Deficit/Hyperactivity Disorder. Proc. Natl. Acad. Sci. USA 2022, 119, e2114094119. [Google Scholar] [CrossRef]
- Chen, J.; Gao, Y.; Bao, S.-T.; Wang, Y.-D.; Jia, T.; Yin, C.; Xiao, C.; Zhou, C. Insula→Amygdala and Insula→Thalamus Pathways Are Involved in Comorbid Chronic Pain and Depression-Like Behavior in Mice. J. Neurosci. 2024, 44, e2062232024. [Google Scholar] [CrossRef]
- Nir, R.-R.; Yarnitsky, D. Conditioned Pain Modulation. Curr. Opin. Support. Palliat. Care 2015, 9, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Wouterlood, F.G.; Groenewegen, H.J. Neuroanatomical Tracing by Use of Phaseolus Vulgaris-Leucoagglutinin (PHA-L): Electron Microscopy of PHA-L-Filled Neuronal Somata, Dendrites, Axons and Axon Terminals. Brain Res. 1985, 326, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Desbois, C.; Le Bars, D.; Villanueva, L. Organization of Cortical Projections to the Medullary Subnucleus Reticularis Dorsalis: A Retrograde and Anterograde Tracing Study in the Rat. J. Comp. Neurol. 1999, 410, 178–196. [Google Scholar] [CrossRef]
- Bouhassira, D.; Villanueva, L.; Bing, Z.; le Bars, D. Involvement of the Subnucleus Reticularis Dorsalis in Diffuse Noxious Inhibitory Controls in the Rat. Brain Res. 1992, 595, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Koutsikou, S.; Apps, R.; Lumb, B.M. Top down Control of Spinal Sensorimotor Circuits Essential for Survival. J. Physiol. 2017, 595, 4151–4158. [Google Scholar] [CrossRef]
- Samineni, V.K.; Grajales-Reyes, J.G.; Sundaram, S.S.; Yoo, J.J.; Gereau, R.W. Cell Type-Specific Modulation of Sensory and Affective Components of Itch in the Periaqueductal Gray. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Walker, R.A.; Wright, K.M.; Jhou, T.C.; McDannald, M.A. The Ventrolateral Periaqueductal Grey Updates Fear via Positive Prediction Error. Eur. J. Neurosci. 2020, 51, 866–880. [Google Scholar] [CrossRef]
- Dampney, R.A.L. Functional Organization of Central Pathways Regulating the Cardiovascular System. Physiol. Rev. 1994, 74, 323–364. [Google Scholar] [CrossRef]
- Davern, P.J. A Role for the Lateral Parabrachial Nucleus in Cardiovascular Function and Fluid Homeostasis. Front. Physiol. 2014, 5, 436. [Google Scholar] [CrossRef]
- Alonso-Matielo, H.; Zhang, Z.; Gambeta, E.; Huang, J.; Chen, L.; de Melo, G.O.; Dale, C.S.; Zamponi, G.W. Inhibitory Insula-ACC Projections Modulate Affective but Not Sensory Aspects of Neuropathic Pain. Mol. Brain 2023, 16, 64. [Google Scholar] [CrossRef]
- Lee, A.T.; Vogt, D.; Rubenstein, J.L.; Sohal, V.S. A Class of GABAergic Neurons in the Prefrontal Cortex Sends Long-Range Projections to the Nucleus Accumbens and Elicits Acute Avoidance Behavior. J. Neurosci. 2014, 34, 11519–11525. [Google Scholar] [CrossRef]
- Masamizu, Y.; Okada, T.; Kawasaki, K.; Ishibashi, H.; Yuasa, S.; Takeda, S.; Hasegawa, I.; Nakahara, K. Local and Retrograde Gene Transfer into Primate Neuronal Pathways via Adeno-Associated Virus Serotype 8 and 9. Neuroscience 2011, 193, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, B.; Gao, Z. Lighting Up Neural Circuits by Viral Tracing. Neurosci. Bull. 2022, 38, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Gehrlach, D.A.; Weiand, C.; Gaitanos, T.N.; Cho, E.; Klein, A.S.; Hennrich, A.A.; Conzelmann, K.-K.K.; Gogolla, N.; Weiand, C.; Hennrich, A.A.; et al. A Whole-Brain Connectivity Map of Mouse Insular Cortex. Elife 2020, 9, e55585. [Google Scholar] [CrossRef] [PubMed]
- Dongyang, L.; Fernandes, A.M.; da Cunha, P.H.M.; Tibes, R.; Sato, J.; Listik, C.; Dale, C.; Kubota, G.T.; Galhardoni, R.; Teixeira, M.J.; et al. Posterior-Superior Insular Deep Transcranial Magnetic Stimulation Alleviates Peripheral Neuropathic Pain—A Pilot Double-Blind, Randomized Cross-over Study. Neurophysiol. Clin. 2021, 51, 291–302. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001; ISBN 0125476361. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, D.; Labrakakis, C. Multiple Posterior Insula Projections to the Brainstem Descending Pain Modulatory System. Int. J. Mol. Sci. 2024, 25, 9185. https://doi.org/10.3390/ijms25179185
Liang D, Labrakakis C. Multiple Posterior Insula Projections to the Brainstem Descending Pain Modulatory System. International Journal of Molecular Sciences. 2024; 25(17):9185. https://doi.org/10.3390/ijms25179185
Chicago/Turabian StyleLiang, Despoina, and Charalampos Labrakakis. 2024. "Multiple Posterior Insula Projections to the Brainstem Descending Pain Modulatory System" International Journal of Molecular Sciences 25, no. 17: 9185. https://doi.org/10.3390/ijms25179185
APA StyleLiang, D., & Labrakakis, C. (2024). Multiple Posterior Insula Projections to the Brainstem Descending Pain Modulatory System. International Journal of Molecular Sciences, 25(17), 9185. https://doi.org/10.3390/ijms25179185