

Exogenous Substances Used to Relieve Plants from Drought Stress and Their Associated Underlying Mechanisms
 , , and
, , and
Abstract
1. Introduction
2. Research and Development of Exogenous Substances to Alleviate Plant Drought Stress
2.1. Overview
2.2. Analysis of Research Hotspots and Frequently Used Exogenous Substances Worldwide
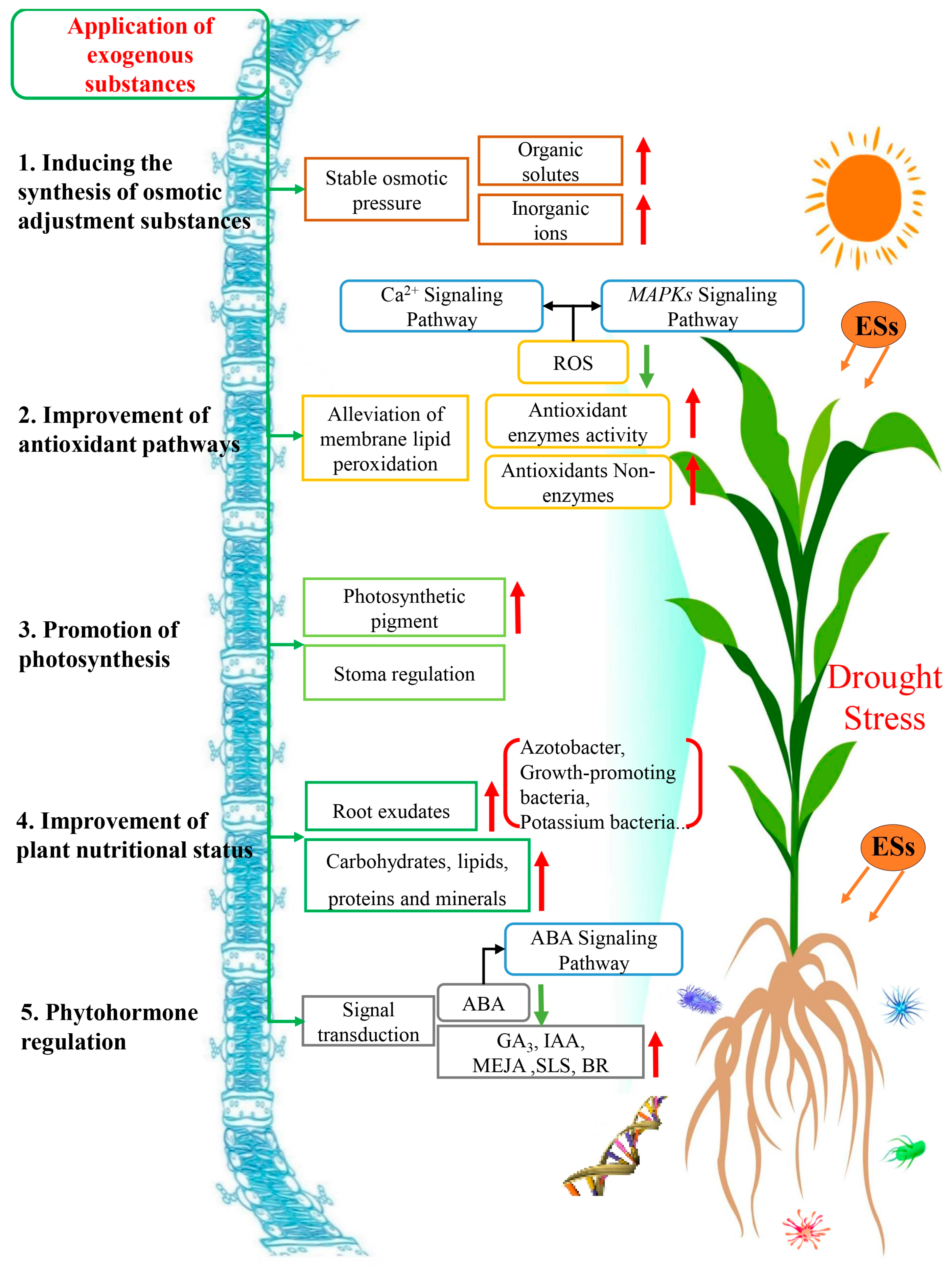
3. The Mechanisms of Exogenous Substances Involved in Alleviating Plant Drought Stress
3.1. Induced Synthesis of Osmotic Regulators
3.2. Improvement of Antioxidant Pathways
3.3. Promotion of Photosynthesis
3.4. Improvement of Plant Nutritional Status
3.5. Phytohormone Regulation
4. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Cook, B.I.; Ault, T.R.; Smerdon, J.E. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci. Adv. 2015, 1, e1400082. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.; McColl, K.A. No projected global drylands expansion under greenhouse warming. Nat. Clim. Chang. 2021, 11, 331–337. [Google Scholar] [CrossRef]
- Li, R.; Sun, R.; Wang, T.; Chen, D.; Li, R.; Li, L.; Zhao, W. Research progress on identification and evaluation methods, and mechanism of drought resistance in plants. Biotechnol. Bull. 2017, 33, 40. [Google Scholar] [CrossRef]
- Schlenker, W.; Lobell, D.B. Robust negative impacts of climate change on African agriculture. Environ. Res. Lett. 2010, 5, 014010. [Google Scholar] [CrossRef]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Siddique, Z.; Jan, S.; Imadi, S.R.; Gul, A.; Ahmad, P. Drought stress and photosynthesis in plants. In Water Stress and Crop Plants: A Sustainable Approach; Wiley Oline Library: New York, NY, USA, 2016; Volume 1, pp. 1–11. [Google Scholar] [CrossRef]
- Alipoor, B.; Moradi, F. Relationship between drought stress and some antioxidant enzymes with cell membrane and chlorophyll stability in wheat lines. Afr. J. Microbiol. Res. 2012, 6, 617–623. [Google Scholar] [CrossRef]
- Singh, A.; Roychoudhury, A. Abscisic acid in plants under abiotic stress: Crosstalk with major phytohormones. Plant Cell Rep. 2023, 42, 961–974. [Google Scholar] [CrossRef]
- Lata, C.; Muthamilarasan, M.; Prasad, M. Drought stress responses and signal transduction in plants. Elucidation Abiotic Stress Signal. Plants Funct. Genom. Perspect. 2015, 2, 195–225. [Google Scholar] [CrossRef]
- Hanaka, A.; Ozimek, E.; Reszczyńska, E.; Jaroszuk-Ściseł, J.; Stolarz, M. Plant tolerance to drought stress in the presence of supporting bacteria and fungi: An efficient strategy in horticulture. Horticulturae 2021, 7, 390. [Google Scholar] [CrossRef]
- Williams, A.; de Vries, F.T. Plant root exudation under drought: Implications for ecosystem functioning. New Phytol. 2020, 225, 1899–1905. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tyagi, S.R.; Wani, M.R.; Ahmad, P. Drought tolerance: Role of organic osmolytes, growth regulators, and mineral nutrients. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Springer: New York, NY, USA, 2013; Volume 1, pp. 25–55. [Google Scholar] [CrossRef]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought tolerance strategies in plants: A mechanistic approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Feng, D.; Gao, Q.; Liu, J.; Tang, J.; Hua, Z.; Sun, X. Categories of exogenous substances and their effect on alleviation of plant salt stress. Eur. J. Agron. 2023, 142, 126656. [Google Scholar] [CrossRef]
- Hossain, M.A.; Wani, S.H.; Bhattacharjee, S.; Burritt, D.J.; Tran, L.-S.P. Drought Stress Tolerance in Plants. In Molecular and Genetic Perspectives; Springer International Publishing: Berlin, Bermany, 2016; Volume 2. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K. Drought stress in plants: An overview. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer International Publishing: Berlin, Bermany, 2012; pp. 1–33. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2012, 172, 1321–1335. [Google Scholar] [CrossRef]
- Xu, Z.; Ma, J.; Lei, P.; Wang, Q.; Feng, X.; Xu, H. Poly-γ-glutamic acid induces system tolerance to drought stress by promoting abscisic acid accumulation in Brassica napus L. Sci. Rep. 2020, 10, 252. [Google Scholar] [CrossRef]
- Guo, H.; Cui, Y.N.; Pan, Y.Q.; Wang, S.M.; Bao, A.K. Sodium chloride facilitates the secretohalophyte Atriplex canescens adaptation to drought stress. Plant Physiol. Biochem. 2020, 150, 99–108. [Google Scholar] [CrossRef]
- Jiao, Z.; Li, Y.; Li, J.; Xu, X.; Li, H.; Lu, D.; Wang, J. Effects of exogenous chitosan on physiological characteristics of potato seedlings under drought stress and rehydration. Potato Res. 2012, 55, 293–301. [Google Scholar] [CrossRef]
- Ahmad, Z.; Anjum, S.; Waraich, E.A.; Ayub, M.A.; Ahmad, T.; Tariq, R.M.S.; Iqbal, M.A. Growth, physiology, and biochemical activities of plant responses with foliar potassium application under drought stress–a review. J. Plant Nutr. 2018, 41, 1734–1743. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. Water Stress Crop Plants A Sustain. Approach 2016, 2, 649–668. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Imran, M.; Kang, S.M.; Park, Y.S.; Wani, S.H.; Lee, I.J. Research progress in the field of microbial mitigation of drought stress in plants. Front. Plant Sci. 2022, 13, 870626. [Google Scholar] [CrossRef]
- Doneva, D.; Pál, M.; Brankova, L.; Szalai, G.; Tajti, J.; Khalil, R.; Ivanovska, B.; Velikova, V.; Misheva, S.; Janda, T.; et al. The effects of putrescine pre-treatment on osmotic stress responses in drought-tolerant and drought-sensitive wheat seedlings. Physiol. Plant. 2021, 171, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, V.; D’Amelia, V.; Esposito, M.; Amitrano, C.; Carillo, P.; Carputo, D.; Maggio, A. Anthocyanins are key regulators of drought stress tolerance in tobacco. Biology 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Thind, S.K.; Bains, N.S. Glycine betaine application modifies biochemical attributes of osmotic adjustment in drought stressed wheat. Plant Growth Regul. 2014, 72, 221–228. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A.; Wahid, M.A.; Khan, H.Z. Biochemically triggered heat and drought stress tolerance in rice by proline application. J. Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]
- Shen, J.; Guo, M.J.; Wang, Y.G.; Yuan, X.Y.; Wen, Y.Y.; Song, X.E.; Guo, P.Y. Humic acid improves the physiological and photosynthetic characteristics of millet seedlings under drought stress. Plant Signal. Behav. 2020, 15, 1774212. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.J.; Nie, Y.X.; Gao, Y.; Dai, A.H.; Bai, J.G. Exogenous cinnamic acid regulates antioxidant enzyme activity and reduces lipid peroxidation in drought-stressed cucumber leaves. Acta Physiol. Plant. 2012, 34, 641–655. [Google Scholar] [CrossRef]
- Dehghan, M.; Balouchi, H.; Yadavi, A.; Zare, E. Improve wheat (Triticum aestivum L.) performance by brassinolide application under different irrigation regimes. S. Afr. J. Bot. 2020, 130, 259–267. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Tung, S.A.; Samad, R.A.; Wang, L.; Shahzad, B. Exogenously applied methyl jasmonate improves the drought tolerance in wheat imposed at early and late developmental stages. Acta Physiol. Plant. 2016, 38, 25. [Google Scholar] [CrossRef]
- Naz, R.; Batool, S.; Shahid, M.; Keyani, R.; Yasmin, H.; Nosheen, A.; Hassan, M.N.; Mumtaz, S.; Siddiqui, M.H. Exogenous silicon and hydrogen sulfide alleviates the simultaneously occurring drought stress and leaf rust infection in wheat. Plant Physiol. Biochem. 2021, 166, 558–571. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Liu, S.; Peng, X. Exogenous application of hydrogen peroxide alleviates drought stress in cucumber seedlings. S. Afr. J. Bot. 2016, 106, 23–28. [Google Scholar] [CrossRef]
- Xu, Q.; Fu, H.; Zhu, B.; Hussain, H.A.; Zhang, K.; Tian, X.; Duan, M.; Xie, X.; Wang, L. Potassium improves drought stress tolerance in plants by affecting root morphology, root exudates, and microbial diversity. Metabolites 2021, 11, 131. [Google Scholar] [CrossRef]
- Nawaz, F.; Ahmad, R.; Ashraf, M.Y.; Waraich, E.A.; Khan, S.Z. Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.T.; Haddad, R. Study of silicon effects on antioxidant enzyme activities and osmotic adjustment of wheat under drought stress. Czech J. Genet. Plant Breed. 2011, 47, 17–27. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; & Dietz, K.J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Song, Y.; Li, J.; Liu, M.; Meng, Z.; Liu, K.; Sui, N. Nitrogen increases drought tolerance in maize seedlings. Funct. Plantbiol. 2019, 46, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Impa, S.M.; Nadaradjan, S.; Jagadish, S.V.K. Drought stress induced reactive oxygen species and anti-oxidants in plants. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Springer: Berlin/Heidelberg, Germany, 2012; pp. 131–147. [Google Scholar]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhriss, M.; Ben Abdullah, F. Exogenous proline effects on photosynthetic performance and antioxidant defense system of young olive tree. J. Agric. Food Chem. 2010, 58, 4216–4222. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Mukherjee, S.; Alamri, S.; Hu, Y.; Alamri, A.; Alsubaie, Q.; Ali, H. Hydrogen sulfide (H2S) and potassium (K+) synergistically induce drought stress tolerance through regulation of H+-ATPase activity, sugar metabolism, and antioxidative defense in tomato seedlings. Plant Cell Rep. 2021, 40, 1543–1564. [Google Scholar] [CrossRef]
- Kolupaev, Y.E.; Karpets, Y.V.; Yastreb, T.O.; Lugovaya, A.A. Combined effect of salicylic acid and nitrogen oxide donor on stress-protective system of wheat plants under drought conditions. Appl. Biochem. Microbiol. 2018, 54, 418–424. [Google Scholar] [CrossRef]
- Sher, A.; Tahira, A.S.; Sattar, A.; Nawaz, A.; Qayyum, A.; Hussain, S.; Manaf, A. Foliage application of 5-aminolevulinic acid alleviates drought stress in sunflower (Helianthus annuus L.) through improving stay green and antioxidant enzymes activities. Acta Physiol. Plant. 2021, 43, 22. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Limson, J.; Weintraub, S.T.; Qi, W. Melatonin directly scavenges hydrogen peroxide: A potentially new metabolic pathway of melatonin biotransformation. Free Radic. Biol. Med. 2000, 29, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biológica Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Ahmad, S.; Kamran, M.; Ding, R.; Meng, X.; Wang, H.; Ahmad, I.; Fahad, S.; Han, Q. Exogenous melatonin confers drought stress by promoting plant growth, photosynthetic capacity and antioxidant defense system of maize seedlings. Peer J. 2019, 7, e7793. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Y.; Li, D.Q. Drought stress signal transduction and regulation mechanism in plants. Biotechnol. Bull. 2012, 10, 1–7. [Google Scholar] [CrossRef]
- Feng, D.; Wang, X.; Gao, J.; Zhang, C.; Liu, H.; Liu, P.; Sun, X. Exogenous calcium: Its mechanisms and research advances involved in plant stress tolerance. Front. Plant Sci. 2023, 14, 1143963. [Google Scholar] [CrossRef]
- Kang, G.Z.; Li, G.Z.; Liu, G.Q.; Xu, W.; Peng, X.Q.; Wang, C.Y.; Zhu, Y.J.; Guo, T.C. Exogenous salicylic acid enhances wheat drought tolerance by influence on the expression of genes related to ascorbate-glutathione cycle. Biol. Plant. 2013, 57, 718–724. [Google Scholar] [CrossRef]
- Islam, M.J.; Uddin, M.J.; Hossain, M.A.; Henry, R.; Begum, M.K.; Sohel, M.A.T.; Cheong, E.J.; Lim, Y.S. Exogenous putrescine attenuates the negative impact of drought stress by modulating physio-biochemical traits and gene expression in sugar beet (Beta vulgaris L.). PLoS ONE 2022, 17, e0262099. [Google Scholar] [CrossRef]
- Aydin, M.; Tombuloglu, G.; Sakcali, M.S.; Hakeem, K.R.; Tombuloglu, H. Boron alleviates drought stress by enhancing gene expression and antioxidant enzyme activity. J. Soil Sci. Plant Nutr. 2019, 19, 545–555. [Google Scholar] [CrossRef]
- Kumar, M.; Mishra, S.; Dixit, V.; Kumar, M.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liang, Z.; Li, Y.; Liao, Y.; Liu, Y. Exogenous spermidine regulates starch synthesis and the antioxidant system to promote wheat grain filling under drought stress. Acta Physiol. Plant. 2020, 42, 110. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Belal Chowdhury, M.; Afrin, S.; Burritt, D.J.; Murata, Y.; Hossain, M.A.; Afzal Hossain, M. Exogenous Glutathione-Mediated Drought Stress Tolerance in Rice (Oryza sativa L.) is Associated with Lower Oxidative Damage and Favorable Ionic Homeostasis. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 955–971. [Google Scholar] [CrossRef]
- Li, J.; Zhao, M.; Liu, L.; Guo, X.; Pei, Y.; Wang, C.; Song, X. Exogenoussorbitol application confers drought tolerance to maize seedlings through upregulating antioxidant system and endogenous sorbitol biosynthesis. Plants 2023, 12, 2456. [Google Scholar] [CrossRef]
- Yildizli, A.; Çevik, S.; Ünyayar, S. Effects of exogenous myo-inositol on leaf water status and oxidative stress of Capsicum annuum under drought stress. Acta Physiol. Plant. 2018, 40, 122. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Exogenous nitric oxide donor and arginine provide protection against short-term drought stress in wheat seedlings. Physiol. Mol. Biol. Plants 2018, 24, 993–1004. [Google Scholar] [CrossRef]
- Abd El-Gawad, H.G.; Mukherjee, S.; Farag, R.; Abd Elbar, O.H.; Hikal, M.; Abou El-Yazied, A.; Abd Elhady, S.A.; Helal, N.; ElKelish, A.; El Nahhas, N.; et al. Exogenous γ-aminobutyric acid (GABA)-induced signaling events and field performance associated with mitigation of drought stress in Phaseolus vulgaris L. Plant Signal. Behav. 2021, 16, 1853384. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Fulvic acid application improves the maize performance under well-watered and drought conditions. J. Agron. Crop Sci. 2011, 197, 409–417. [Google Scholar] [CrossRef]
- Elkelish, A.; El-Mogy, M.M.; Niedbała, G.; Piekutowska, M.; Atia, M.A.M.; Hamada, M.M.A.; Shahin, M.; Mukherjee, S.; El-Yazied, A.A.; Shebl, M.; et al. Roles of Exogenous α-Lipoic Acid and Cysteine in Mitigation of Drought Stress and Restoration of Grain Quality in Wheat. Plants 2021, 10, 2318. [Google Scholar] [CrossRef]
- Xie, H.; Bai, G.; Lu, P.; Li, H.; Fei, M.; Xiao, B.G.; Chen, X.J.; Tong, Z.J.; Wang, Z.Y.; Yang, D.H. Exogenous citric acid enhances drought tolerance in tobacco (Nicotiana tabacum). Plant Biol. 2022, 24, 333–343. [Google Scholar] [CrossRef]
- Li, D.M.; Nie, Y.X.; Zhang, J.; Yin, J.S.; Li, Q.; Wang, X.J.; Bai, J.G. Ferulic acid pretreatment enhances dehydration-stress tolerance of cucumber seedlings. Biol. Plant. 2013, 57, 711–717. [Google Scholar] [CrossRef]
- Zhou, H.T.; Zhang, Y.Y.; Li, T.L.; Cao, L.X.; Jin, Y.N.; Zhang, X.J. S-excitation is the best exogenous substance for relieving drought stress of oat (Avena sativa) seed. Mol. Plant Breed. 2019, 17, 7182–7189. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Y.; Xie, Y.; Wang, Y.; Duan, L.; Zhang, M.; Li, Z. Ethephon improved drought tolerance in maize seedlings by modulating cuticular wax biosynthesis and membrane stability. J. Plant Physiol. 2017, 214, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, F.; Li, M.; Liang, D.; Zou, J. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions. Plant Growth Regul. 2011, 64, 63–74. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous melatonin improves seedling health index and drought tolerance in tomato. Plant Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Ehsanpour, A. Influence of exogenously applied paclobutrazol on some physiological traits and growth of Stevia rebaudiana under in vitro drought stress. Biologia 2013, 68, 414–420. [Google Scholar] [CrossRef]
- Da Costa, V.A.; Cothren, J.T. Drought effects on gas exchange, chlorophyll, and plant growth of 1-Methylcyclopropene treated cotton. Agron. J. 2011, 103, 1230–1241. [Google Scholar] [CrossRef]
- Xie, T.L.; Gu, W.R.; Zhang, L.G.; Li, L.J.; Qu, D.Y.; Li, C.F.; Meng, Y.; Li, J.; Wei, S.; Li, W.H. Modulating the antioxidant system by exogenous 2-(3, 4-dichlorophenoxy) triethylamine in maize seedlings exposed to polyethylene glycol-simulated drought stress. PLoS ONE 2018, 13, e0203626. [Google Scholar] [CrossRef]
- Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Nitric oxide stimulates antioxidant system and osmotic adjustment in soybean under drought stress. J. Soil Sci. Plant Nutr. 2020, 20, 1122–1132. [Google Scholar] [CrossRef]
- Lin, Y.H.; Jin, Y.K.; Chen, Z.Y.; Xiao, Z.D.; Shen, S.; Zhou, S.L. Exogenous Methylglyoxal Ameliorates Source Strength and Retrieves Yield Loss Under Drought Stress During Grain Filling in Maize. J. Plant Growth Regul. 2022, 42, 3934–3946. [Google Scholar] [CrossRef]
- Sattar, A.; Ul-Allah, S.; Ijaz, M.; Sher, A.; Butt, M.; Abbas, T.; Irfan, M.; Fatima, T.; Alfarraj, S.; Alharbi, S.A. Exogenous application of strigolactone alleviates drought stress in maize seedlings by regulating the physiological and antioxidants defense mechanisms. Cereal Res. Commun. 2022, 50, 263–272. [Google Scholar] [CrossRef]
- Li, J.A.; Tang, H.L.; Chen, H.P. Effect of exogenous carbon monoxide on antioxidative system in rice seedlings under drought stress. Acta Bot. Boreali-Occident. Sin. 2010, 30, 330–335. [Google Scholar] [CrossRef]
- Ippolito, M.P.; Fasciano, C.; d’Aquino, L.; Tommasi, F. Responses of antioxidant systems to lanthanum nitrate treatments in tomato plants during drought stress. Plant Biosyst. 2011, 145, 248–252. [Google Scholar] [CrossRef]
- Cui, W.; Kamran, M.; Song, Q.; Zuo, B.; Jia, Z.; Han, Q. Lanthanum chloride improves maize grain yield by promoting photosynthetic characteristics, antioxidants enzymes and endogenous hormone at reproductive stages. J. Rare Earths 2019, 37, 781–790. [Google Scholar] [CrossRef]
- Alabdallah, N.M.; HASAN, M.; Salih, A.M.; SS, R.; Al-Shammari, A.S.; Alsanie, S.I.; El-Zaidy, M. Silver nanoparticles improve growth and protect against oxidative damage in eggplant seedlings under drought stress. Plant Soil Environ. 2021, 67, 617–624. [Google Scholar] [CrossRef]
- Liu, Z.J.; Zhang, X.L.; Bai, J.G.; Suo, B.X.; Xu, P.L.; Wang, L. Exogenous paraquat changes antioxidant enzyme activities and lipid peroxidation in drought-stressed cucumber leaves. Sci. Hortic. 2009, 121, 138–143. [Google Scholar] [CrossRef]
- Jacomassi, L.M.; Viveiros, J.D.O.; Momesso, L.; Crusciol, C.A.C. A seaweed extract-based biostimulant mitigates drought stress in sugarcane. Front. Plant Sci. 2022, 13, 865291. [Google Scholar] [CrossRef] [PubMed]
- Cornic, G.; Massacci, A. Leaf photosynthesis under drought stress. In Photosynthesis and the Environment; Springer: Dordrecht, The Netherlands, 1996; pp. 347–366. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef]
- Zargar, S.M.; Gupta, N.; Nazir, M.; Mahajan, R.; Malik, F.A.; Sofi, N.R.; Shikari, A.B.; Salgotra, R.K. Impact of drought on photosynthesis: Molecular perspective. Plant Gene 2017, 11, 154–159. [Google Scholar] [CrossRef]
- Liang, D.; Ni, Z.; Xia, H.; Xie, Y.; Lv, X.; Wang, J.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Ma, H.; Li, P.; Liu, X.; Li, C.; Zhang, S.; Wang, X.; Tao, X. Poly-γ-glutamic acid enhanced the drought resistance of maize by improving photosynthesis and affecting the rhizosphere microbial community. BMC Plant Biol. 2022, 22, 11. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.Q.; Liu, Q. Photosynthesis product allocation and yield in sweet potato with spraying exogenous hormones under drought stress. J. Plant Physiol. 2020, 253, 153265. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Hu, W.; Tian, S.B.; Di, Q.; Duan, S.H.; Dai, K. Effects of exogenous calcium on mesophyll cell ultrastructure, gas exchange, and photosystem II in tobacco (Nicotiana tabacum Linn.) under drought stress. Photosynthetica 2018, 56, 1204–1211. [Google Scholar] [CrossRef]
- Teng, K.Q.; Li, J.Z.; Liu, L.; Han, Y.C.; Du, Y.X.; Zhang, J.; Sun, H.Z.; Zhao, Q.Z. Exogenous ABA induces drought tolerance in upland rice: The role of chloroplast and ABA biosynthesis-related gene expression on photosystem II during PEG stress. Acta Physiol. Plant. 2014, 36, 2219–2227. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, S.; Wang, J.; Su, X.; Suo, B.; Qin, F.; Zhao, H. Exogenous application of 5-aminolevulinic acid on wheat seedlings under drought stress enhances the transcription of psbA and psbD genes and improves photosynthesis. Braz. J. Bot. 2018, 41, 275–285. [Google Scholar] [CrossRef]
- Timmusk, S.; Wagner, E.G. The Plant-Growth-Promoting Rhizobacterium Paenibacillus polymyxa Induces Changes in Arabidopsis thaliana Gene Expression: A Possible Connection Between Biotic and Abiotic Stress Responses. Mol. Plant Microbe Interact. 1999, 12, 951–959. [Google Scholar] [CrossRef]
- Fonseca, M.d.C.d.; Bossolani, J.W.; de Oliveira, S.L.; Moretti, L.G.; Portugal, J.R.; Scudeletti, D.; de Oliveira, E.F.; Crusciol, C.A.C. Bacillus subtilis inoculation improves nutrient uptake and physiological activity in sugarcane under drought stress. Microorganisms 2022, 10, 809. [Google Scholar] [CrossRef]
- Han, R.; Gao, G.; Li, Z.; Dong, Z.; Guo, Z. Effects of exogenous 5-aminolevulinic acid on seed germination of alfalfa (Medicago varia Martyn.) under drought stress. Grassl. Sci. 2018, 64, 100–107. [Google Scholar] [CrossRef]
- Wasaya, A.; Abbas, T.; Yasir, T.A.; Sarwar, N.; Aziz, A.; Javaid, M.M.; Akram, S. Mitigating drought stress in sunflower (Helianthus annuus L.) through exogenous application of β-aminobutyric acid. J. Soil Sci. Plant Nutr. 2021, 21, 936–948. [Google Scholar] [CrossRef]
- Zhao, H.; Tan, J.; Qi, C. Photosynthesis of Rehmannia glutinosa subjected to drought stress is enhanced by choline chloride through alleviating lipid peroxidation and increasing proline accumulation. Plant Growth Regul. 2007, 51, 255–262. [Google Scholar] [CrossRef]
- Egilla, J.; Davies, F.; Boutton, T. Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 2005, 43, 135–140. [Google Scholar] [CrossRef]
- Kang, J.; Peng, Y.; Xu, W. Crop root responses to drought stress: Molecular mechanisms, nutrient regulations, and interactions with microorganisms in the rhizosphere. Int. J. Mol. Sci. 2022, 23, 9310. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria that confer resistance to water stress in tomatoes and peppers. Plant Sci 2004, 166, 525–530. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Réthoré, E.; Pluchon, S.; Ali, N.; Billiot, B.; Yvin, J.C. Calcium application enhances drought stress tolerance in sugar beet and promotes plant biomass and beetroot sucrose concentration. Int. J. Mol. Sci. 2019, 20, 3777. [Google Scholar] [CrossRef]
- Egilla, J.N.; Davies, F.T.; Drew, M.C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. leprechaun: Plant growth, leaf macro-and micronutrient content and root longevity. Plant Soil 2001, 229, 213–224. [Google Scholar] [CrossRef]
- Kosar, F.; Alshallash, K.S.; Akram, N.A.; Sadiq, M.; Ashraf, M.; Alkhalifah, D.H.M.; Abdel Latef, A.A.H.; Elkelish, A. Trehalose-induced regulations in nutrient status and secondary metabolites of drought-stressed sunflower (Helianthus annuus L.) plants. Plants 2022, 11, 2780. [Google Scholar] [CrossRef]
- Fiere, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Yue-Yu, S.; Zhang, X.D.; Kai, M.; Herbert, S. Spatial variability of nutrient properties in black soil of northeast China. Pedosphere 2007, 17, 19–29. [Google Scholar] [CrossRef]
- Castrillo, G.; Teixeira, P.J.P.L.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, C.D. Root microbiota drive direct integration of phosphate stress and immunity. Nature 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Gholamhoseini, M.; Ghalavand, A.; Dolatabadian, A.; Jamshidi, E.; Khodaei-Joghan, A. Effects of arbuscular mycorrhizal inoculation on growth, yield, nutrient uptake and irrigation water productivity of sunflowers grown under drought stress. Agric. Water Manag. 2013, 117, 106–114. [Google Scholar] [CrossRef]
- Liu, J.; Wang, G.; Jin, J.; Liu, J.; Liu, X. Effects of different concentrations of phosphorus on microbial communities in soybean rhizosphere grown in two types of soils. Ann. Microbiol. 2011, 61, 525–534. [Google Scholar] [CrossRef]
- Daszkowska-Golec, A. The role of abscisic acid in drought stress: How ABA helps plants to cope with drought stress. Drought Stress Tolerance in Plants, Vol 2, Molecular and Genetic Perspectives; Springer International Publishing: Berlin, Bermany, 2016; pp. 123–151. [Google Scholar] [CrossRef]
- Burgess, P.; Huang, B. Mechanisms of hormone regulation for drought tolerance in plants. In Drought Stress Tolerance in Plants; Springer: Berlin, Bermany, 2016; Volume 1, pp. 45–75. [Google Scholar] [CrossRef]
- Christmann, A.; Grill, E.; Huang, J. Hydraulic signals in long-distance signaling. Curr. Opin. Plant Biol. 2013, 16, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Testerink, C.; Zhang, Y. How roots and shoots communicate through stressful times. Trends Plant Sci. 2021, 26, 940–952. [Google Scholar] [CrossRef]
- Ding, L.; Li, Y.R.; Wang, Y.; Gao, L.M.; Wang, M., Chaumont; Shen, Q.R.; Guo, S.W. Root ABA accumulation enhances rice seedling drought tolerance under ammonium supply: Interaction with aquaporins. Front. Plant Sci. 2016, 7, 1206. [Google Scholar] [CrossRef]
- Yuan, G.F.; Jia, C.G.; Li, Z.; Sun, B.; Zhang, L.P.; Liu, N.; Wang, Q.M. Effect of brassinosteroids on drought resistance and abscisic acid concentration in tomato under water stress. Sci. Hortic. 2010, 126, 103–108. [Google Scholar] [CrossRef]
- Sedaghat, M.; Emam, Y.; Mokhtassi-Bidgoli, A.; Hazrati, S.; Lovisolo, C.; Visentin, I.; Cardinale, F.; Tahmasebi-Sarvestani, Z. The potential of the synthetic strigolactone analogue GR24 for the maintenance of photosynthesis and yield in winter wheat under drought: Investigations on the mechanisms of action and delivery modes. Plants 2021, 10, 1223. [Google Scholar] [CrossRef]
- Xing, X.H.; Xui, Z.J.; Qi, Y.J.; Wang, X.J.; Sun, D.L.; Bian, N.F.; Wang, X. Effect of exogenous α-naphthaleneacetic acid on carbon metabolism of soybean under drought stress at flowering stage. Chin. J. Appl. Ecol. 2018, 29, 1215–1224. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O.; Aremu, B.R. Genomic insights into plant growth promoting rhizobia capable of enhancing soybean germination under drought stress. BMC Microbiol. 2019, 19, 159. [Google Scholar] [CrossRef]
- Li, C.Y.; Kong, X.Q.; Luo, Z.; Li, W.J.; Tang, W.; Zhang, D.M.; Ma, C.L.; Dong, H.Z. Exogenous application of acetic acid improves the survival rate of cotton by increasing abscisic acid and jasmonic acid contents under drought stress. Acta Physiol. Plant. 2021, 43, 32. [Google Scholar] [CrossRef]
- Akter, N.; Rafiqul Islam, M.; Abdul Karim, M.; Hossain, T. Alleviation of drought stress in maize by exogenous application of gibberellic acid and cytokinin. J. Crop Sci. Biotechnol. 2014, 17, 41–48. [Google Scholar] [CrossRef]
- Chang, Z.; Liu, Y.; Dong, H.; Teng, K.; Han, L.; Zhang, X. Effects of Cytokinin and Nitrogen on Drought Tolerance of Creeping Bentgrass. PLoS ONE 2016, 11, e0154005. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Wang, S.; Li, J.; Li, X.; Zang, H.; Fang, B. The changes of wheat seedlings in drought condition by exogenous coronatine (COR). Acta Agric. Scand. Sect. B—Soil Plant Sci. 2020, 70, 467–473. [Google Scholar] [CrossRef]
- Fernández, V.; Bahamonde, H.A.; Peguero-Pina, J.J.; Gil-Pelegrín, E.; Sancho-Knapik, D.; Gil, L.; Goldbach, H.E.; Eichert, T. Physico-chemical properties of plant cuticles and their functional and ecological significance. J. Exp. Bot. 2017, 68, 5293–5306. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhang, J.; Wu, Z.; Loka, D.A.; Zhao, W.; Chen, B.; Wang, Y.; Meng, Y.; Zhou, Z.; Gao, L. Effects of single and combined exogenous application of abscisic acid and melatonin on cotton carbohydrate metabolism and yield under drought stress. Ind. Crops Prod. 2022, 176, 114302. [Google Scholar] [CrossRef]
- Tayyab, N.; Naz, R.; Yasmin, H.; Nosheen, A.; Keyani, R.; Sajjad, M.; Hassan, M.N.; Roberts, T.H. Combined seed and foliar pre-treatments with exogenous methyl jasmonate and salicylic acid mitigate drought-induced stress in maize. PLoS ONE 2020, 15, e0232269. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Polyamine | ||
|---|---|---|
| Spermidine (Spd) | Putrescine (Put) | |
| Polysaccharide | ||
| Chitosan (CTS) | Trehalose | |
| Polyphenol | ||
| Anthocyanin (AC) | ||
| Polypeptide | ||
| Glutathione (GSH) | ||
| Polyol | ||
| Sorbitol (ST) | Inositol | |
| Amino Acid | ||
| Glycine Betaine (GB) | 5-Aminolevulinic acid (5-ALA) | Arginine (Arg) |
| γ-Aminobutyric acid (GABA) | γ-polyglutamic acid (γ-PGA) | β-aminobutyric acid (BABA) |
| Proline (Pro) | ||
| Organic Acid | ||
| Humic acid (HA) | Fulvic acid (FA) | Salicylic acid (SA) |
| α-Lipoic Acid (ALA) | Cinnamic acid (CA) | Acetic acid (HAc) |
| Citric acid | Ferulic acid (FA) | |
| Phytohormone | ||
| S-excitation (S-ABA) | Ethephon (ETH) | Gibberellins (GA3) |
| Abscisic acid (ABA) | Melatonin (MT) | Brassinolide (BR) |
| Methyl jasmonate (MeJA) | Cytokinin (CTK) | Strigolactones (SLS) |
| 6-Benzylaminoadenine (6-BA) | ||
| Plant-growth Regulator | ||
| Coronatine (COR) | Paclobutrazol (PBZ) | 1-methylcyclopropene (1-MCP) |
| α-naphthaleneacetic acid (NAA) | 2-(3,4-Dichlorophenoxy) triethylamine (DCPTA) | Choline chloride (Cc) |
| Signaling Molecule | ||
| Nitric oxide (NO) | Hydrogen sulfide (H2S) | Methylglyoxal (MG) |
| Hydrogen peroxide (H2O2) | Carbon monoxide (CO) | |
| Element | ||
| Calcium (Ca) | Potassium Kalium (K) | Selenium (Se) |
| Silicon (Si) | Boron (B) | Phosphorus (P) |
| Compound | ||
| Lanthanum nitrateon | lanthanum chloride | Sodium chloride (NaCl) |
| Microorganism | ||
| Arbuscular mycorrhizal | Rhizobium | NBRIRA |
| NBRISN13 | Bacillus subtilis | |
| Other | ||
| AgNPs | Paraquat (PQ) | Seaweed extract (SWE) |
| Exogenous Substance | No. of Papers (Topic Search) | No. of Papers (Refined by MeSH) | Mechanisms Involved |
|---|---|---|---|
| Abscisic acid (ABA) | 1547 | 420 | 2, 3, 5 |
| Hydrogen peroxide (H2O2) | 855 | 147 | 1, 2 |
| Glutathione (GSH) | 379 | 44 | 2 |
| Melatonin (MT) | 346 | 124 | 2, 3 |
| Nitric oxide (NO) | 332 | 62 | 2 |
| Potassium kalium (K) | 279 | 16 | 1, 3, 4 |
| Calcium (Ca) | 273 | 28 | 3, 4 |
| Glycine betaine (GB) | 185 | 16 | 1 |
| Spermidine (Spd) | 183 | 28 | 1, 5 |
| Methyl jasmonate (MeJA) | 160 | 29 | 1, 5 |
| Exogenous Substance | Optimal Concentration | Application Method | Crop | Mechanism of Exogenous Substances | Reference |
|---|---|---|---|---|---|
| Putrescine | 100 μM | Hydroponics | Wheat (Triticum aestivum L.) | An effective reduction in the osmotic stress and increase in the photosynthetic capacity and proline concentration of crops under DS. | [26] |
| Chitosan | 100 mg L−1 | Foliar spraying | Wheat (Triticum aestivum L.) | Maintains the integrity and stability of the cell membranes of seedlings and increases the proline concentration and soluble protein content through foliar spraying at the early stage of drought to increase the soluble sugar content in leaves for a better osmotic adjustment. | [22] |
| Anthocyanin | 4 mgL−1 | Foliar spraying | Tobacco (Nicotiana tabacum L.) | Used to promote the accumulation of sucrose and amino acids, especially proline, and alleviate osmotic impairments. | [27] |
| Glycine Betaine | 100 mM | Foliar spraying | Wheat (Triticum aestivum L.) | Enhances the osmotic adjustment, increases the photosynthetic rate, upregulates the STI gene of wheat genotypes, and accumulates more proline and endogenous betaine through reducing the sucrose content in leaves. | [28] |
| γ-polyglutamic acid | 20 mgL−1 | Hydroponics | Rice (Oryza sativa L.) | An increase in the proline concentration and stabilization of plant osmotic pressure. | [20] |
| Proline | 30 mM | Foliar spraying | Rice (Oryza sativa L.) | Increases the activity of total soluble protein, proline, and glycine betaine in leaves, promotes the K+ absorption efficiency, and produces more osmotic protective agents. | [29] |
| Humic acid | 200 mg L−1 | Seed soaking | Millet (Setaria italica (L.) Beauv.) | Promotes the accumulation of soluble protein and free proline and improves the osmotic adjustment ability. | [30] |
| cinnamic acid | 50 μM | Drench | Cucumbers (Cucumis sativus L.) | Increases the contents of ascorbic acid, proline, soluble sugar, vanillic acid (VA), and CA in leaves and enhances the osmotic adjustment ability. | [31] |
| Brassinolide | 1 μM | Foliar spraying | wheat (Triticum aestivum L.) | A positive effect on maintaining photosynthetic capacity and initiating osmotic protection and other hormone induction. | [32] |
| Methyl jasmonate | 0.5 mM | Foliar spraying | Wheat (Triticum aestivum L.) | Used for promoting the accumulation of total soluble sugars, polysaccharides and carbohydrates, enhancing the accumulation of osmotic protective agents, initiating stomatal closures, escalating the water use efficiency, and inducing the transport of assimilates to increase yield. | [33] |
| Hydrogen sulfide | 0.3 mM | Foliar spraying | Cucumbers (Cucumis sativus L.) | Promotes the total soluble sugar, protein, and proline content, improves the osmotic adjustment ability, and alleviates oxidative damages. | [34] |
| Hydrogen peroxide | 1.5 mM | Foliar spraying | Cucumbers (Cucumis sativus L.) | Promotes accumulating soluble sugar and proline, improves osmotic adjustments, and enhances antioxidant defenses and photosynthesis. | [35] |
| Potassium Kalium | 80 mgkg−1 | Fertilization | Rape (Brassica napus L.) | Increases the root length and root density, balances the root-to-shoot ratio to increase root water uptake potential, stimulates the root secretion of organic acids, and promotes nutrient acquisition and utilization. | [36] |
| Selenium | 40 mg L−1 | Foliar spraying | Wheat (Triticum aestivum L.) | It affects the accumulation of osmotic adjustment substances, reduces osmotic potential, and promotes the accumulation of soluble sugar and free amino acids. | [37] |
| Silicon | 2 mM | Fertilization | Wheat (Triticum aestivum L.) | Enhances osmotic adjustment ability; increases aquaporin activity, root water conductivity, and leaf water content; and promotes the nutrient renewal and photosynthetic rate. | [38] |
| Sodium chloride | 100 mM | Drench | Grass leaved orache (Atriplex patens) | Increases the content of glycine betaine and soluble sugar in leaves, accumulates Na+ to enhance water absorption, and reduces the retention of Na+ in photosynthetic organs, thereby protecting cell membrane and structure. | [21] |
| Exogenous Substance | Optimal Concentration | Application Method | Crop | Mechanism of Exogenous Substances | Reference |
|---|---|---|---|---|---|
| Spermidine | 1 mM | Drench | Wheat (Triticum aestivum L.) | Used to promote the activities of SOD, POD and CAT in plant grains and reduce the MDA content. | [55] |
| Putrescine | 100 μM | Hydroponics | Wheat (Triticum aestivum L.) | Used to alleviate oxidative damage, improve the antioxidant capacity, and enhance the GST and POD activity. | [26] |
| Chitosan | 100 mg L−1 | Foliar spraying | Potato (Solanum tuberosum) | Used to increase the activity of SOD and POD in leaves, remove the active oxygen, and stabilize cell membranes. | [22] |
| Trehalose | 30 mM | Foliar spraying | Maize (Zea mays L.) | Used to reduce malondialdehyde and SOD activity, and increase the POD and CAT activity. | [56] |
| Glutathione | 0.2 mM | Hydroponics | Cotton (Gossypium spp.) | Used to inhibit the accumulation of ROS in plant cells; upregulate genes coding for enzymatic and non-enzymatic antioxidants such as CAT, ascorbate peroxidase (APX), peroxidase (POX), reduce ascorbic acid (AsA), glutathione peroxidase (GSH), etc.; mitigate the severity of ROS-induced oxidative damage; and decrease H2O2 and malondialdehyde (MDA) accumulation. | [57] |
| Sorbitol | 10 mM | Foliar spraying | Maize (Zea mays L.) | Used to increase the activity of SOD, POD, CAT, and ASA, GSH and enhance the antioxidant capacity. | [58] |
| Inositol | 15 μM | Foliar spraying | Capsicum (Capsicum annuum L.) | Used to reduce the MDA content and enhance scavenging ROS by increasing POD and GR activities. | [59] |
| 5-Aminolevulinic acid | 75 mgL−1 | Foliar spraying | Sunflower (Helianthus annuus L.) | Used to increase chlorophylls and the activity of antioxidant enzymes (APX, SOD and CAT). | [45] |
| Arginine | 0.1 mM | Hydroponics | Wheat (Triticum aestivum L.) | Used to increase the activity of CAT, GPX, GST, and other antioxidant enzymes, and to increase the endogenous NO content for the purpose of regulating the antioxidant system and reducing ROS production. | [60] |
| γ-Aminobutyric acid | 2 mM | Foliar spraying | Bean (Phaseolus vulgaris L.) | Used to increase the activity of SOD, CAT, POX, and APX. | [61] |
| Proline | 30 mM | Foliar spraying | Rice (Oryza sativa L.) | Used to promote the activity of SOD, POD, and CAT and improve the antioxidant capacity. | [29] |
| Humic acid | 200 mg L−1 | Seed soaking | Millet (Setaria italica (L.) Beauv.) | Used to reduce H2O2 and increase the activity of SOD, POD, and CAT. | [30] |
| Fulvic acid | 1.5 mgL−1 | Foliar spraying | Maize (Zea mays L.) | Used to increase the SOD, POD, and CAT activity and proline level and to maintain the chlorophyll content and gas exchange rate. | [62] |
| α-Lipoic Acid | 0.02 mM | Seed soaking | Wheat (Triticum aestivum L.) | Used to enhance the activity of SOD, APX, CAT, and POX and remove ROS. | [63] |
| Cinnamic acid | 50 μM | Drench | Cucumbers (Cucumis sativus L.) | Used to enhance the activity of GPX, glutathione peroxidase (GSH-Px), DHAR, and GR and reduce the lipid peroxidation. | [31] |
| Citric acid | 50 mM | Foliar spraying | Tobacco (Nicotiana tabacum L.) | Used to enhance the activity of POD and CAT and reduce the ROS accumulation. | [64] |
| Ferulic acid | 0.5 mM | Drench | Cucumbers (Cucumis sativus L.) | Used to inhibit the ROS production; reduce the MDA content; induce the activity of SOD, CAT, GPX, GSH, and APX in leaves; and increase the proline and soluble sugar content. | [65] |
| S-excitation | 1000-fold | Seed soaking | Oat (Avena sativa L.) | Used to reduce the MDA content and increase the antioxidase activity. | [66] |
| Ethephon | 1.0 mM | Foliar spraying | Maize (Zea mays L.) | Used to reduce MDA and hydrogen peroxide and to increase proline and the activities of SOD, POD, and CAT, thereby reducing oxidative damage and maintaining membrane integrity and stability. | [67] |
| Abscisic acid | 60 μM | Foliar spraying | Kiwi fruit (Actinidia) | Used to keep cell membranes undamaged and promote the activity of the POD, CAT, SOD, APX, and GR antioxidant enzymes. | [68] |
| Melatonin | 0.1 mM | Foliar spraying | Tomato (Solanum lycopersicum) | Used to decrease the MDA content and increase the activity of POD, SOD, CAT, APX, and GR, as well as the content of ASA. | [69] |
| Brassinolide | 1 μM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to reduce the MDA content and alleviate oxidative stress. | [32] |
| Paclobutrazol | 2 mM | Hydroponics | Stevia (Stevia rebaudiana) | Used to reduce membrane damage, prevent the leakage of electrolytes, and decrease the MDA level. | [70] |
| 1-Methylcyclopropene | 2.4 g L−1 | Gas | Cotton (Gossypium spp.) | Used to maintain the integrity of cell membranes through increasing the activity of antioxidant enzymes such as POD. | [71] |
| 2-(3,4-Dichlorophenoxy) triethylamine | 15 mM | Hydroponics | Maize (Zea mays L.) | The activities of POD and CAT are enhanced, and the accumulation of reactive oxygen species is inhibited. | [72] |
| Nitric oxide | 100 μM | Foliar spraying | Soybean (Glycine max (Linn.) Merr) | Used to improve the activity of the SOD, CAT, APX, and POX antioxidant enzymes and alleviate oxidative damage. | [73] |
| Hydrogen sulfide | 0.3 mM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to reduce the content of H2O2 and MDA and improve the oxidative stress tolerance through increasing the activity of antioxidant enzymes. | [34] |
| Methylglyoxal | 15–25 mM | Foliar spraying | Maize (Zea mays L.) | Used to inhibit the accumulation of endogenous MG and activate the glyoxalase system in leaves. | [74] |
| Strigolactones | 20 μM | Foliar spraying | Maize (Zea mays L.) | Used to enhance the activity of antioxidant enzymes such as POD, SOD, CAT, and APX. | [75] |
| Hydrogen peroxide | 1.5 mM | Foliar spraying | Cucumbers (Cucumis sativus L.) | Used to increase the activity of SOD and POD and improve the ability of leaves to scavenge ROS. | [35] |
| Carbon monoxide | 0.1 μM | Hydroponics | Rice (Oryza sativa L.) | Used to promote the activity of SOD, CAT, and POD in leaves and enhance the antioxidant capacity. | [76] |
| Selenium | 40 mg L−1 | Foliar spraying | Wheat (Triticum aestivum L.) | Used to promote the activity of antioxidant enzymes such as CAT, POX, and APX. | [37] |
| Silicon | 2 mM | Fertilization | Wheat (Triticum aestivum L.) | Used to increase the activity of SOD, CAT, APX and POD and alleviate the damage of oxide film. | [38] |
| Boron | 50 μM | Hydroponics | Tomato (Solanum lycopersicum) | Used to reduce the MDA content and increase the APX and glutathione reductase (GR) activity. | [53] |
| Lanthanum nitrateon | 10 mM | Foliar spraying | Tomato (Solanum lycopersicum) | Used to increase the activity of all enzymes involved in the ASC-GSH cycle. | [77] |
| Lanthanum chloride | 400 mM | Seed soaking | Maize (Zea mays L.) | Used to enhance the activity of POD, CAT, and SOD. | [78] |
| AgNPs | 0.1 μM | Foliar spraying | Egg plant (Solanum melongena L.) | Used to increase the content of H2O2 and MDA and to promote the activity of SOD and CAT antioxidant enzymes. | [79] |
| Paraquat | 10 mM | Drench | Cucumbers (Cucumis sativus L.) | Used to increase the activity of SOD, CAT, GPX, APX, DHAR, MDHAR, GR, GSH, and AsA. | [80] |
| Seaweed extract | 0.5 L/ha | Fertilization | Cane (Saccharumofficinarum L.) | Increases the activity of SOD, POD, CAT, and PPO, and promotes the balance of ROS. | [81] |
| Exogenous Substance | Optimal Concentration | Application Method | Crop | Mechanism of Exogenous Substances | Reference |
|---|---|---|---|---|---|
| 5-Aminolevulinic acid | 10 mgL−1 | Seed soaking | Alfalfa (Medicago sativa L.) | Used to increase the chlorophyll content and photosynthetic rate of leaves, decrease the stomatal density of cotyledons, and increase the stomatal width. | [94] |
| γ-polyglutamic acid | 50 mgL−1 | Fertilization | Maize (Zea mays L.) | Used to promote the accumulation of chlorophyll and light and parameters and improve the net photosynthetic rate and stomatal conductance. | [86] |
| β-aminobutyric acid | 75 mM | Foliar spraying | Sunflower (Helianthus annuus L.) | Used to promote the green retention by increasing SPAD-chlorophyll content. | [95] |
| Humic acid | 200 mg L−1 | Seed soaking | Millet (Setaria italica (L.) Beauv.) | Used to repair damages to chlorophylls, prevent chlorophyll degradation, increase the stomatal conductance, and increase the photosynthetic rate. | [30] |
| Abscisic acid | 60 μM | Foliar spraying | Rice (Oryza sativa L.) | Used to improve photosynthesis through accumulating chlorophyll fluorescence and upregulating the expression of chloroplast genes. | [90] |
| Melatonin | 0.1 mM | Foliar spraying | Tomato (Solanum lycopersicum) | Used to enhance the chlorophyll metabolism of leaves, promote the accumulation of chlorophyll, delay the decomposition process, and maintain a high level of photosynthetic efficiency. | [69] |
| 6-Benzylaminoadenine | 60 mgL−1 | Foliar spraying | Sweetpotato (Dioscorea esculenta L.) | Used to increase the stomatal conductance, CO2 utilization, and net photosynthetic rate. | [87] |
| Choline chloride | 2.1 mM | Foliar spraying | Rehmannia glutinosa (Rehmannia) | Used to maintain a high level of leaf water content, delay leaf water loss, and promote the recovery of photosynthesis after rehydration. | [96] |
| Hydrogen peroxid | 1.5 mM | Foliar spraying | Cucumbers (Cucumis sativus L.) | Used to increase the chlorophyll content and leaf water content and improve the photosynthetic capacity. | [35] |
| Calcium | 10 mM | Foliar spraying | Tobacco (Nicotiana tabacum L.) | Used to stabilize the structure and function of chloroplast, mitochondria and inner membranes in mesophyll cells, maintain the normal net photosynthetic rate and gas exchange in leaves, and minimize the degradation of photosynthetic pigments. | [89] |
| Potassium Kalium | 2.5 mM | Fertilization | Hibiscus (Hibiscus syriacus L.) | Used to promote the increase in K+ concentration in chloroplasts to maintain the photosynthetic activity. | [97] |
| Silicon | 1.5 mM | Fertilization | Rice (Oryza sativa L.) | Used to improve the photosynthetic rate and mitigate impairments on photochemical reactions. | [88] |
| Exogenous Substance | Optimal Concentration | Application Method | Crop | Mechanism of Exogenous Substances | Reference |
|---|---|---|---|---|---|
| Spermidine | 1 mM | Pouring | Wheat (Triticum aestivum L.) | Used to regulate the endogenous hormone level, promote the synthesis of cytokinin (CTK) and starch in grains, and reduce the synthesis of ethylene. | [55] |
| Acetic acid | 8 mM | Pouring | Cotton (Gossypium spp.) | Used to activate more gene expression for higher levels of endogenous ABA and JA. | [117] |
| Gibberellins | 50 mgL−1 | Foliar spraying | Maize (Zea mays L.) | Used to increase the division of damaged cells, promote the enzyme activity, and balance the endogenous hormones through increasing the content of endogenous gibberellin. | [118] |
| Abscisic acid | 60 μM | Foliar spraying | Rice (Oryza sativa L.) | Used to regulate stomatal opening and closure, reduce transpiration, and inhibit the synthesis of endogenous ABA to balance the changes of endogenous hormone levels. | [90] |
| Brassinolide | 1 μM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to maintain normal photosynthetic activity, produce more antioxidants, generate osmotic protection, and induce other hormones involved in DS alleviation. | [32] |
| Methyl jasmonate | 0.5 mM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to affect the synthesis of endogenous hormones in plants, promote the accumulation of total soluble sugars, polysaccharides, carbohydrates, and osmotic protective agents; enhance the stomatal closure; improve the water use efficiency; and induce the transport of assimilates to increase yield. | [33] |
| Cytokinin | 100 μM | Foliar spraying | Creeping herbs (Agrostis stolonifera L.) | Used to repair damages due to membrane lipid peroxidation and promote the nitrogen metabolism. | [119] |
| Strigolactones | 20 μM | Foliar spraying | Maize (Zea mays L.) | Used to increase the leaf water content and chlorophyll content and improve the activity of antioxidant enzymes. | [75] |
| Coronatine | 10 μM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to boost the JA or jasmonate signaling pathway and ABA accumulation and to regulate the balance of endogenous hormones. | [120] |
| α-naphthaleneacetic acid | 40 mgL−1 | Foliar spraying | Soybean (Glycine max (Linn.) Merr) | Used to promote the movement of soluble sugar from leaves to roots, promote the distribution of photosynthetic assimilates to sucrose, inhibit the conversion of sucrose to starch and hexose, increase the photosynthetic rate, and reduce the inhibition of photosynthesis. | [115] |
| Hydrogen sulfide | 0.3 mM | Foliar spraying | Wheat (Triticum aestivum L.) | Used to increase the endogenous SA content, regulate the endogenous ABA content, and maintain the hormone balance. | [34] |
| lanthanum chloride | 400 mM | Seed soaking | Maize (Zea mays L.) | Used to improve the photosynthetic rate and antioxidant enzymes and to change the levels of endogenous hormones such as auxin and gibberellin during the reproductive period. | [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, D.; Liu, W.; Chen, K.; Ning, S.; Gao, Q.; Chen, J.; Liu, J.; Sun, X.; Xu, W. Exogenous Substances Used to Relieve Plants from Drought Stress and Their Associated Underlying Mechanisms. Int. J. Mol. Sci. 2024, 25, 9249. https://doi.org/10.3390/ijms25179249
Feng D, Liu W, Chen K, Ning S, Gao Q, Chen J, Liu J, Sun X, Xu W. Exogenous Substances Used to Relieve Plants from Drought Stress and Their Associated Underlying Mechanisms. International Journal of Molecular Sciences. 2024; 25(17):9249. https://doi.org/10.3390/ijms25179249
Chicago/Turabian StyleFeng, Di, Wenxin Liu, Ke Chen, Songrui Ning, Qian Gao, Jiao Chen, Jiao Liu, Xiaoan Sun, and Wanli Xu. 2024. "Exogenous Substances Used to Relieve Plants from Drought Stress and Their Associated Underlying Mechanisms" International Journal of Molecular Sciences 25, no. 17: 9249. https://doi.org/10.3390/ijms25179249
APA StyleFeng, D., Liu, W., Chen, K., Ning, S., Gao, Q., Chen, J., Liu, J., Sun, X., & Xu, W. (2024). Exogenous Substances Used to Relieve Plants from Drought Stress and Their Associated Underlying Mechanisms. International Journal of Molecular Sciences, 25(17), 9249. https://doi.org/10.3390/ijms25179249