
High-Throughput Transcriptomic Analysis of Circadian Rhythm of Chlorophyll Metabolism under Different Photoperiods in Tea Plants

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
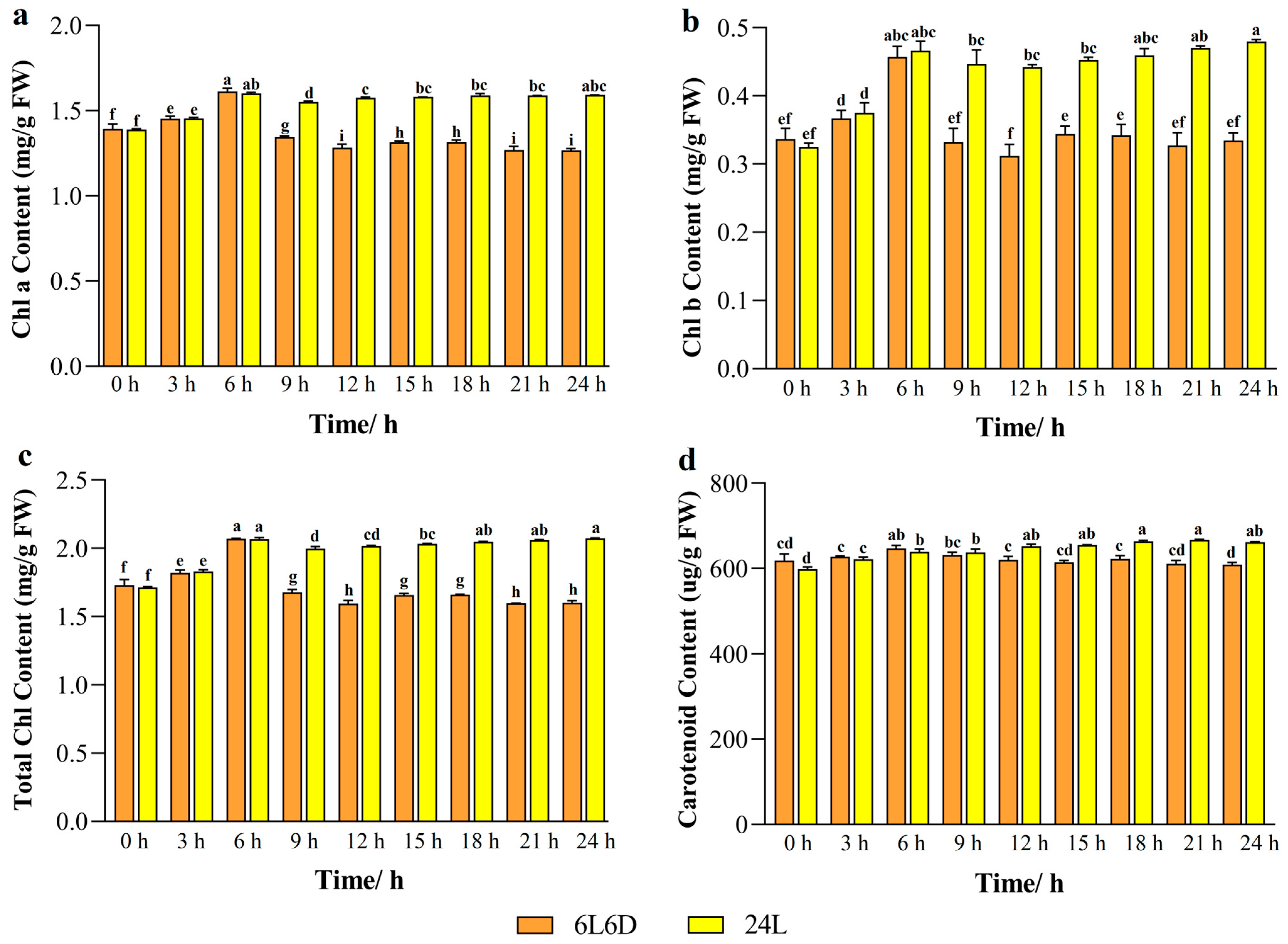
2.1. Chlorophyll and Carotenoid Content in Tea Plants Under Different Photoperiods
2.2. Quality Analysis of Tea Transcriptome Sequencing at Different Time Points
2.3. Functional Annotation and Classification
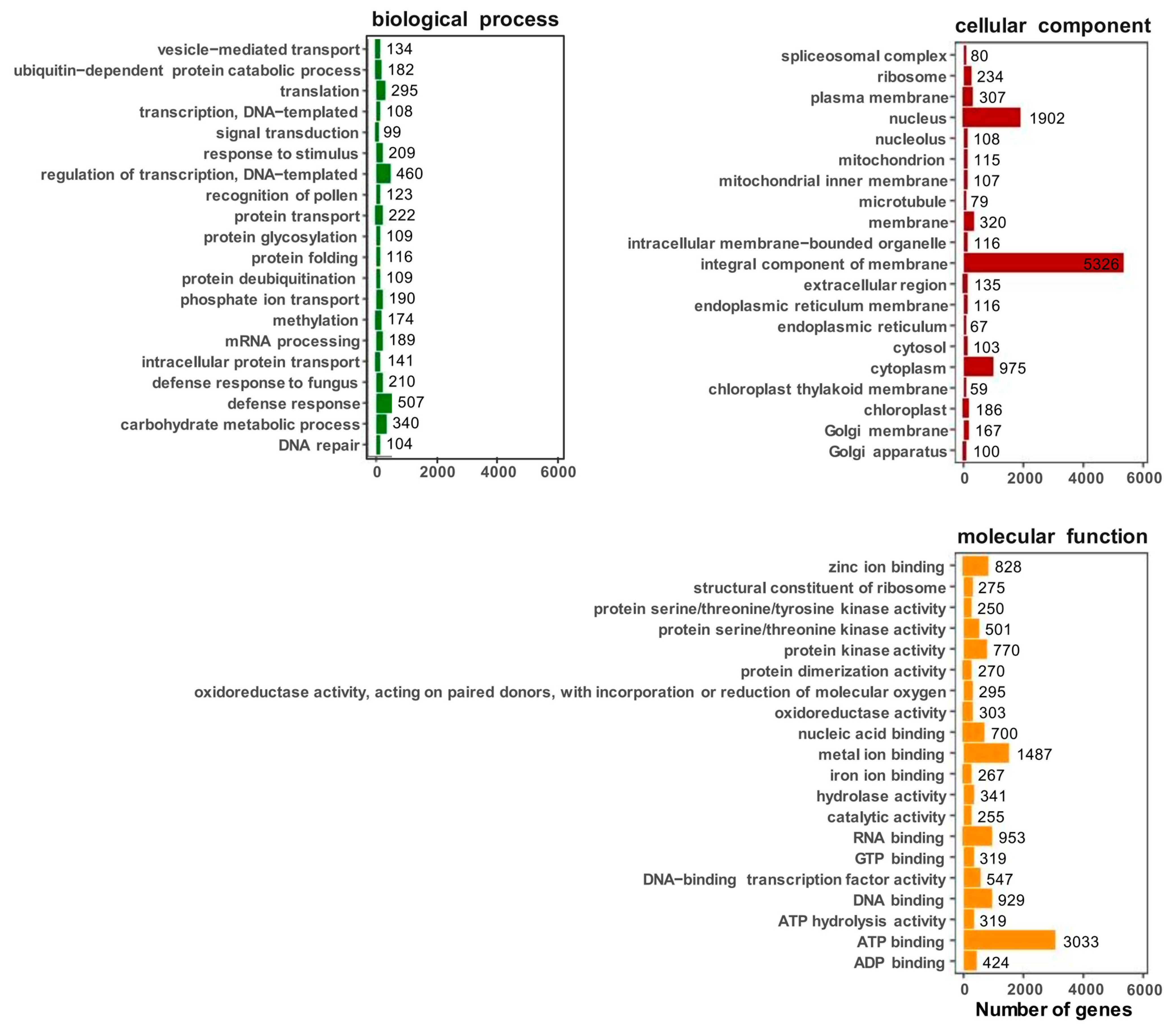
2.4. GO Analysis
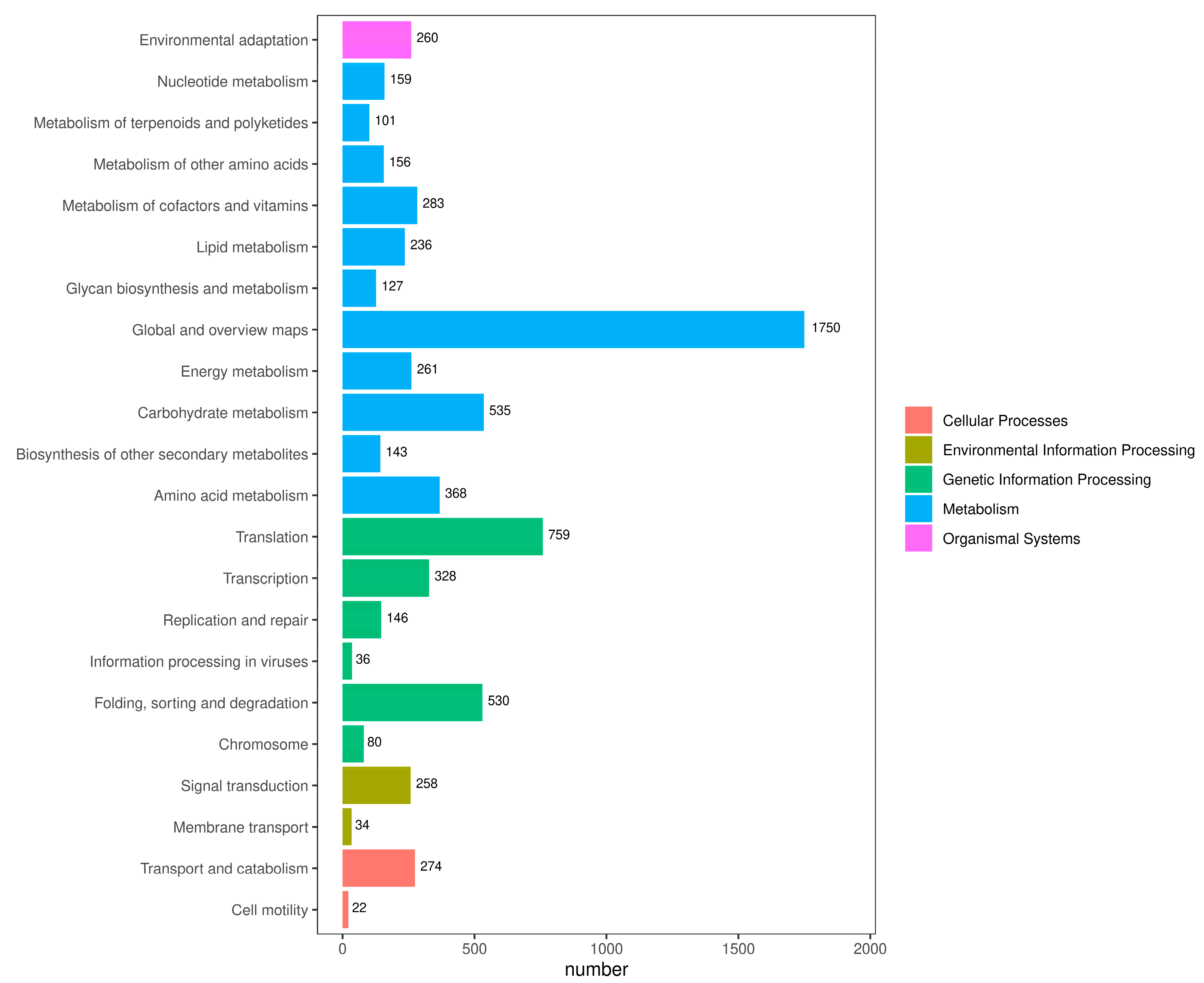
2.5. KEGG Classification
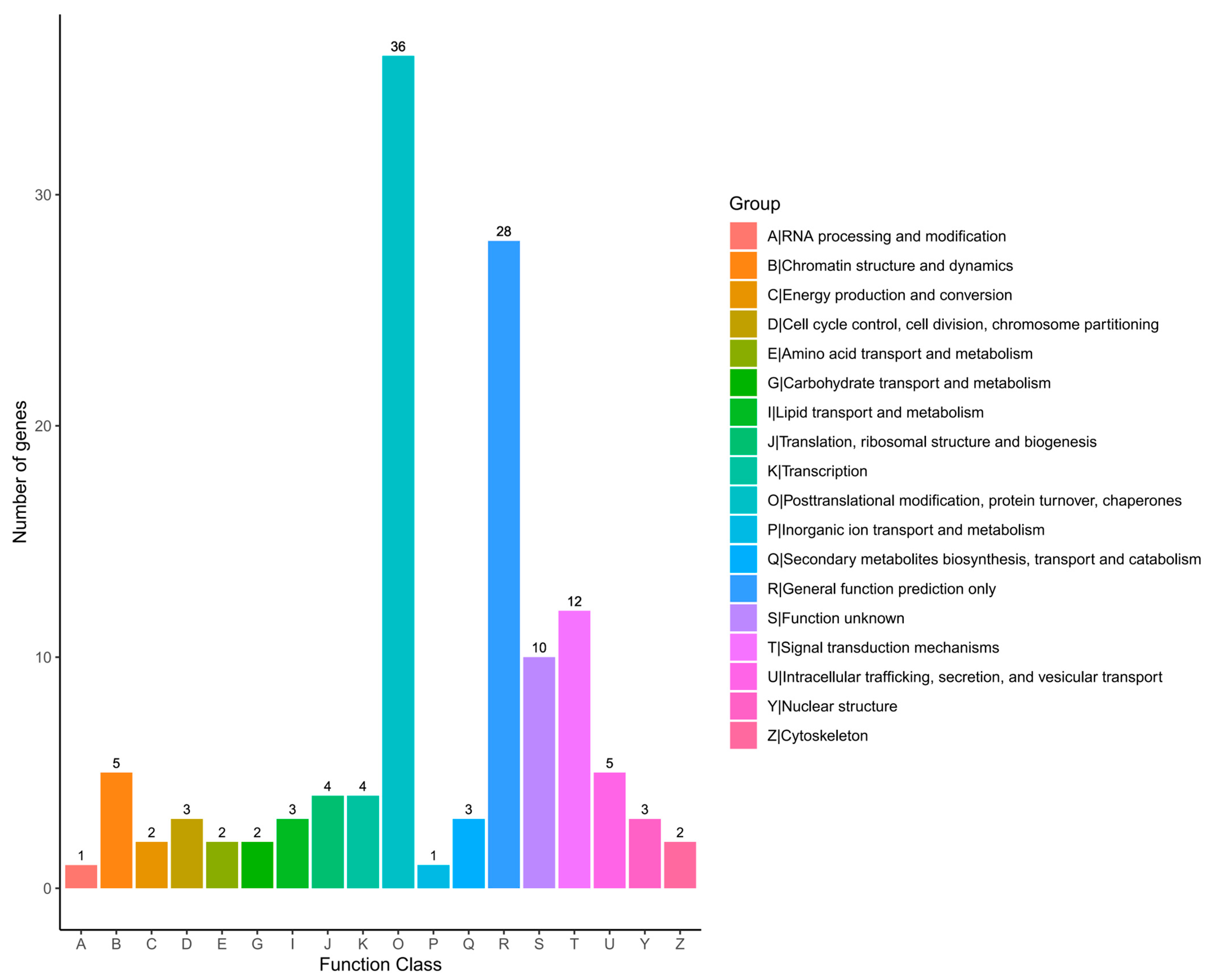
2.6. KOG Classification
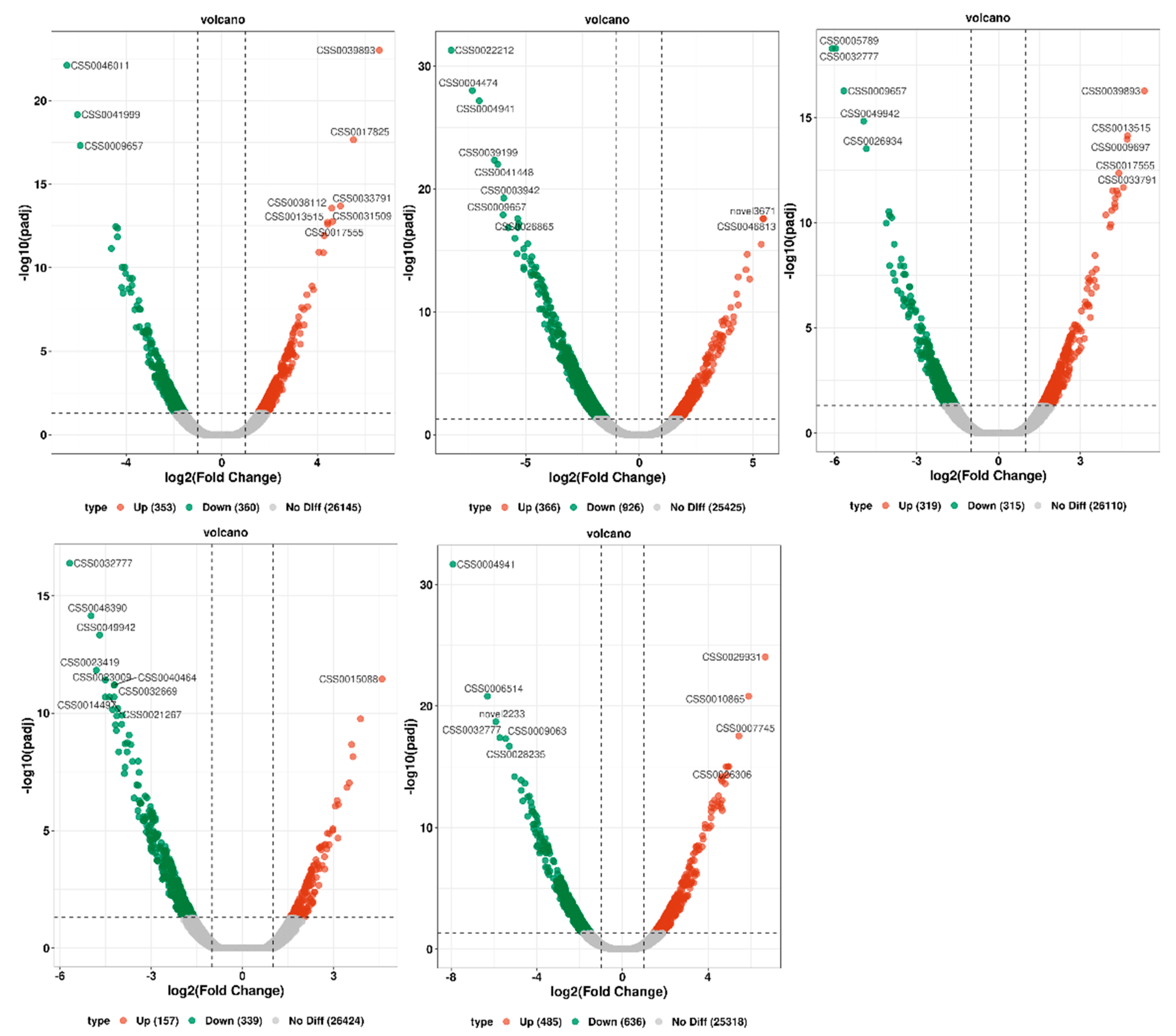
2.7. Screening of Differentially Expressed Genes in the Tea Transcriptome at Different Time Points
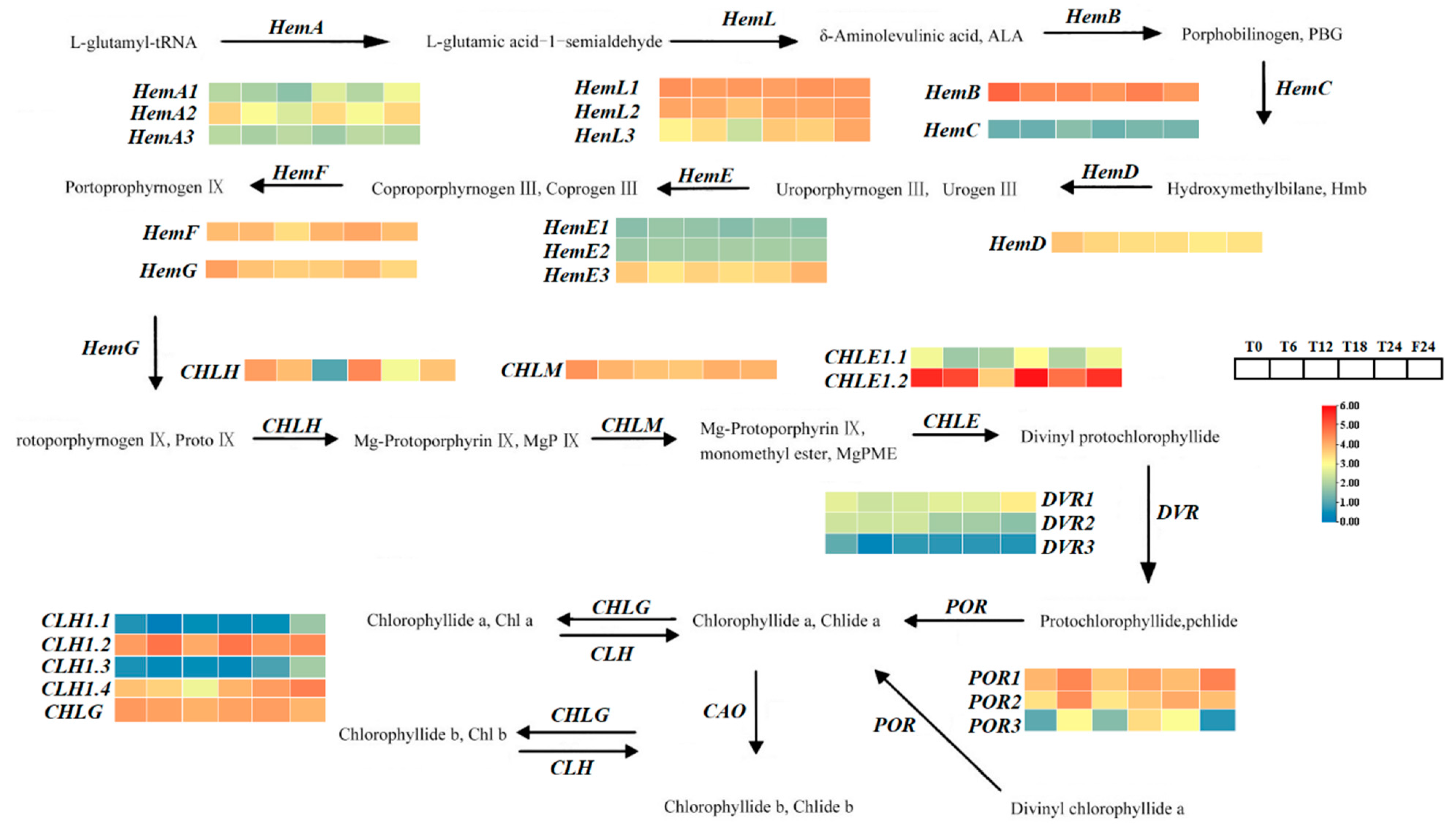
2.8. Screening of Different Genes in the Chlorophyll Metabolic Pathway in Tea Plants at Different Time Points
2.9. Fluorescence Quantitative PCR Verification
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Extraction and Measurement of Chlorophyll and Carotenoid Content
4.3. RNA Extraction, cDNA Library Construction and Sequencing
4.4. RNA-Seq Data and Enrichment Analysis of Differentially Expressed Genes
4.5. Functional Annotation Analysis
4.6. RT-qPCR Validation of Differentially Expressed Genes
4.7. Data Processing and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, X.M.; Liu, J.X.; Li, H.Y.; Wang, W.Z.; Liu, L.; Wang, P.Q.; Hu, J.H.; Zhang, X.F.; Qu, F.F. Greenhouse covering cultivation promotes chlorophyll accumulation of tea plant (Camellia sinensis) by activating relevant gene expression and enzyme activity. BMC Plant Biol. 2024, 24, 455. [Google Scholar] [CrossRef]
- Wu, J.Q.; Cheng, J.; Xu, C.M.; Qi, S.L.; Sun, W.R.; Wu, S. AUREA maintains the balance between chlorophyll synthesis and adventitious root formation in tomato. Hortic. Res. 2020, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S. Update on the biochemistry of chlorophyll breakdown. Plant Mol. Biol. 2013, 82, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Geng, R.D.; Pang, X.Q.; Li, X.; Shi, S.S.; Hedtke, B.; Grimm, B.; Bock, R.; Huang, J.R.; Zhou, W.B. PROGRAMMED CELL DEATH8 interacts with tetrapyrrole biosynthesis enzymes and ClpC1 to maintain homeostasis of tetrapyrrole metabolites in Arabidopsis. New Phytol. 2023, 238, 2545–2560. [Google Scholar] [CrossRef]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef]
- Suzuki, J.Y.; Bollivar, D.W.; Bauer, C.E. Genetic analysis of chlorophyll biosynthesis. Annu. Rev. Genet. 1997, 31, 61–89. [Google Scholar] [CrossRef] [PubMed]
- Cornah, J.E.; Terry, M.J.; Smith, A.G. Green or red: What stops the traffic in the tetrapyrrole pathway? Trends Plant Sci. 2003, 8, 224–230. [Google Scholar] [CrossRef]
- Li, G.Z.; Chen, X.; Zhao, Y.C.; Zhao, D.G. Gene expression regulation of the effect of shading on chlorophyll content in Fuding White Tea (Camellia sinensis L.). Tree Physiol. 2024, tpae049. [Google Scholar] [CrossRef]
- Li, Y.C.; Jeyaraj, A.; Yu, H.P.; Wang, Y.; Ma, Q.P.; Chen, X.; Sun, H.W.; Zhang, H.; Ding, Z.T.; Li, X.H. Metabolic Regulation Profiling of Carbon and Nitrogen in Tea Plants [Camellia sinensis (L.) O. Kuntze] in Response to Shading. J. Agric. Food. Chem. 2020, 68, 961–974. [Google Scholar] [CrossRef]
- Tamilselvi, E.; Anburaj, J.; Haripriya, D.; Santhosh, A.; Rajakumar, G.; Kavya, P.; Li, X.H. Influence of shading intensity on chlorophyll, carotenoid and metabolites biosynthesis to improve the quality of green tea: A review. Energy Nexus 2023, 12, 100241. [Google Scholar]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.X.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from chloroplasts converge to regulate nuclear gene expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- Sanchez, S.E.; Kay, S.A. The Plant Circadian Clock: From a Simple Timekeeper to a Complex Developmental Manager. Cold Spring Harb. Perspect. Biol. 2016, 8, a027748. [Google Scholar] [CrossRef] [PubMed]
- Duanmu, D.Q.; Casero, D.; Dent, R.M.; Gallaher, S.; Yang, W.Q.; Rockwell, N.C.; Martin, S.S.; Pellegrini, M.; Niyogi, K.K.; Merchant, S.S.; et al. Retrograde bilin signaling enables Chlamydomonas greening and phototrophic survival. Proc. Natl. Acad. Sci. USA 2013, 110, 3621–3626. [Google Scholar] [CrossRef]
- He, J.J.; Yang, J.P.; Yang, H.; Zhao, X.; Ye, X.Y. Effects of light intensity and nitrogen supply on the dynamic characteristics of leaf SPAD value of rice canopy. J. Zhejiang Univ. (Agric. Life Sci.) 2014, 40, 495–504. [Google Scholar]
- Ma, S.Y.; Li, S.; Niu, J.Y.; Zhang, Z.; Liu, Y.; Xue, C. Effects of different LED light on physiological and biochemical characters of grape rootstock plantlets. J. Gansu Agric. Univ. 2010, 45, 56–62. [Google Scholar]
- Wu, Z.J.; Li, X.H.; Liu, Z.W.; Xu, Z.S.; Zhuang, J. De novo assembly and transcriptome characterization: Novel insights into catechins biosynthesis in Camellia sinensis. BMC Plant Biol. 2014, 14, 277. [Google Scholar] [CrossRef]
- Liu, Z.W.; Li, H.; Liu, J.X.; Wang, Y.; Zhuang, J. Integrative transcriptome, proteome, and microRNA analysis reveals the effects of nitrogen sufficiency and deficiency conditions on theanine metabolism in the tea plant (Camellia sinensis). Hortic. Res. 2020, 7, 65. [Google Scholar] [CrossRef]
- Hu, Z.H.; Zhang, N.; Qin, Z.Y.; Li, J.W.; Yang, N.; Chen, Y.; Kong, J.Y.; Luo, W.; Xiong, A.S.; Zhuang, J. Differential Response of MYB Transcription Factor Gene Transcripts to Circadian Rhythm in Tea Plants (Camellia sinensis). Int. J. Mol. Sci. 2024, 25, 657. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.P. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–72. [Google Scholar]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The Universal Protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef]
- Sysoeva, M.I.; Markovskaya, E.F.; Shibaeva, T. Plants under continuous light: Review. Plant Stress 2010, 4, 5–17. [Google Scholar]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, K.; Omura, Y.; Kozai, T. Effects of air temperature regimes on physiological disorders and floral development of tomato seedlings grown under continuous light. HortScience 2005, 40, 1304–1306. [Google Scholar] [CrossRef]
- Demers, D.A.; Dorais, M.; Wien, C.H.; Gosselin, A. Effects of supplemental light duration on greenhouse tomato (Lycopersicon esculentum Mill.) plants and fruit yields. Sci. Hortic. 1998, 74, 295–306. [Google Scholar] [CrossRef]
- Li, H.Y.; Liu, H.H. Effects of Supplementary Illumination at Night on Hormones Content and Nutrient Absorption of Cucumber Seedlings. Chin. Agric. Sci. Bull. 2013, 29, 74–78. [Google Scholar]
- Ali, M.B.; Khandaker, L.; Oba, S. Comparative study on functional components, antioxidant activity and color parameters of selected colored leafy vegetables as affected by photoperiods. J. Food Agric. Environ. 2009, 7, 392–398. [Google Scholar]
- Park, J.E.; Park, Y.; Jeong, B.R.; Seungjae, H. Growth of lettuce in closed-type plant production system as affected by light intensity and photoperiod under influence of white LED light. Agric. Food Sci. 2013, 22, 228–233. [Google Scholar] [CrossRef]
- Gaudreau, L.; Charbonneau, J.; Vézina, L. Photoperiod and Photosynthetic Photon Flux Influence Growth and Quality of Greenhouse-grown Lettuce. HortScience 1994, 29, 1285–1289. [Google Scholar] [CrossRef]
- He, W.; Chen, D.Y.; Hu, X.T.; Wang, X.X.; Chen, L.H.; Zhang, H.C.; Yang, Z.C. Effects of different photoperiods and photonflux ratios of red and blur LEDs on growth and development of tomato plants. Acta Agric. Borealioccidentalis Sin. 2018, 27, 562–570. [Google Scholar]
- Kang, J.H.; KrishnaKuma, S.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hortic. Environ. Biotechnol. 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Stenbaek, A.; Jensen, P.E. Redox regulation of chlorophyll biosynthesis. Phytochemistry 2010, 71, 853–859. [Google Scholar] [CrossRef]
- Eckhardt, U.; Grimm, B.; Hörtensteiner, S. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol. Biol. 2004, 56, 1–14. [Google Scholar] [CrossRef]
- Bollivar, D.W. Recent advances in chlorophyll biosynthesis. Photosynth. Res. 2006, 90, 173–194. [Google Scholar] [CrossRef]
- Hu, X.Y.; Gu, T.Y.; Khan, I.; Zada, A.; Jia, T. Research Progress in the Interconversion, Turnover and Degradation of Chlorophyll. Cells 2021, 10, 3134. [Google Scholar] [CrossRef]
- Papenbrock, J.; Mock, H.P.; Kruse, E.; Grimm, B. Expression studies in tetrapyrrole biosynthesis: Inverse maxima of magnesium chelatase and ferro chelatase activity during cyclic photoperiods. Planta 1999, 208, 264–273. [Google Scholar] [CrossRef]
- Zhao, Y.Q.; Wang, W.J.; Zhan, X.H.; Zhang, M.Y.; Xiao, Y.; Hou, X.R.; Gao, M.; Xiao, B.; Gao, Y.F. CsCHLI plays an important role in chlorophyll biosynthesis of tea plant (Camellia sinensis). Beverage Plant Res. 2024, 4, e004. [Google Scholar] [CrossRef]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef]
- Ji, D.; Li, Q.; Guo, Y.; An, W.; Manavski, N.; Meurer, J.; Chi, W. NADP+ supply adjusts the synthesis of photosystem I in Arabidopsis chloroplasts. Plant Physiol. 2022, 189, 2128–2143. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Fusada, N.; Oosawa, N.; Takamatsu, K.; Yamamoto, Y.Y.; Ohto, M.; Nakamura, K.; Goto, K.; Shibata, D.; Shirano, Y. Functional analysis of isoforms of NADPH: Protochlorophyllide oxidoreductase (POR), PORB and PORC, in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 963–974. [Google Scholar] [CrossRef]
- Deng, Q.; Du, P.; Gangurde, S.S.; Hong, Y.; Xiao, Y.; Hu, D.; Li, H.; Lu, Q.; Li, S.; Liu, H. ScRNA-seq reveals dark- and light-induced differentially expressed gene atlases of seedling leaves in Arachis hypogaea L. Plant Biotechnol. J. 2024, 22, 1848–1866. [Google Scholar] [CrossRef]
- Chen, H.; Wu, W.Q.; Du, K.; Ling, A.; Kang, X.Y. The interplay of growth-regulating factor 5 and BZR1 in coregulating chlorophyll degradation in poplar. Plant Cell Environ. 2024. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Jia, T.; Hörtensteiner, S.; Tanaka, A.; Tanaka, R. Subcellular localization of chlorophyllase2 reveals it is not involved in chlorophyll degradation during senescence in Arabidopsis thaliana. Plant Sci. 2020, 290, 110314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Heyes, D.J.; Feng, L.; Sun, W.; Johannissen, L.O.; Liu, H.; Levy, C.W.; Li, X.; Yang, J.; Yu, X. Structural basis for enzymatic photocatalysis in chlorophyll biosynthesis. Nature 2019, 574, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yan, J.R.; Chen, X.Y.; Jiang, C.H.; Meng, S.D.; Liu, Y.F.; Xu, T.; Qi, M.F.; Li, T.L. Light Regulation of Chlorophyll Biosynthesis in Plants. Acta Hortic. Sin. 2019, 46, 975–994. [Google Scholar]
- Xu, X.; Chi, W.; Sun, X.; Feng, P.; Guo, H.; Li, J.; Lin, R.; Lu, C.; Wang, H.; Leister, D. Convergence of light and chloroplast signals for de-etiolation through ABI4-HY5 and COP1. Nat. Plants 2016, 2, 16066. [Google Scholar] [CrossRef]
- Huang, T.; Liu, H.; Tao, J.P.; Zhang, J.Q.; Zhao, T.M.; Hou, X.L.; Xiong, A.S.; You, X. Low light intensity elongates period and defers peak time of photosynthesis: A computational approach to circadian-clock-controlled photosynthesis in tomato. Hortic. Res. 2023, 10, uhad077. [Google Scholar] [CrossRef]
- Hu, Z.H.; Huang, T.; Zhang, N.; Chen, C.; Yang, K.X.; Sun, M.Z.; Yang, N.; Cheng, Y.; Tao, J.P.; Liu, H. Interference of skeleton photoperiod on circadian clock and photosynthetic efficiency of tea plant: In-depth analysis of mathematical model. Hortic. Res. 2024, uhae226. [Google Scholar] [CrossRef]
- Yu, X.L.; Hu, S.; He, C.; Zhou, J.T.; Qu, F.F.; Ai, Z.Y.; Chen, Y.Q.; Ni, D.J. Chlorophyll Metabolism in Postharvest Tea (Camellia sinensis L.) Leaves: Variations in Color Values, Chlorophyll Derivatives, and Gene Expression Levels under Different Withering Treatments. J. Agric. Food Chem. 2019, 67, 10624–10636. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, W.; Niu, D.; Liu, X.M. Effects of abiotic stress on chlorophyll metabolism. Plant Sci. 2024, 342, 112030. [Google Scholar] [CrossRef]
- Davis, E.M.; Sun, Y.; Liu, Y.; Kolekar, P.; Shao, Y.; Szlachta, K.; Mulder, H.L.; Ren, D.; Rice, S.V.; Wang, Z.; et al. SequencErr: Measuring and suppressing sequencer errors in next-generation sequencing data. Genome Biol. 2021, 22, 37. [Google Scholar] [CrossRef]
- Brown, J.; Pirrung, M.; McCue, L.A. FQC Dashboard: Integrates FastQC results into a web-based, interactive, and extensible FASTQ quality control tool. Bioinformatics 2017, 33, 3137–3139. [Google Scholar] [CrossRef]
- Si, Y.M.; Xing, Y.Q.; Cai, L. Differential splicing event analysis of liver tumor-educated blood platelets RNA-seq data with Hisat2 and MISO. J. Inn. Mong. Univ. Sci. Technol. 2016, 34, 274–279. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Li, H.; Liu, Z.W.; Wu, Z.J.; Wang, Y.X.; Teng, R.M.; Zhuang, J. Differentially expressed protein and gene analysis revealed the effects of temperature on changes in ascorbic acid metabolism in harvested tea leaves. Hortic. Res. 2018, 5, 65. [Google Scholar] [CrossRef]
- Dragan, A.I.; Pavlovic, R.; McGivney, J.B.; Casas-Finet, J.R.; Bishop, E.S.; Strouse, R.J.; Schenerman, M.A.; Geddes, C.D. SYBR Green I: Fluorescence properties and interaction with DNA. J. Fluoresc. 2012, 22, 1189–1199. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Reads | Clean Bases | Q20 Rate | Q30 Rate | GC Content |
|---|---|---|---|---|---|
| T0 h | 43,291,008 | 6,042,417,256 | 98.606% | 95.866% | 44.054% |
| T6 h | 43,294,090 | 6,071,741,360 | 98.397% | 96.011% | 43.882% |
| T12 h | 43,299,782 | 6,084,730,076 | 98.533% | 96.154% | 43.767% |
| T18 h | 43,297,578 | 6,037,877,452 | 97.840% | 94.419% | 44.213% |
| T24 h | 43,315,956 | 5,785,374,886 | 98.800% | 96.873% | 44.225% |
| F24 h | 43,325,480 | 6,027,509,621 | 98.460% | 96.353% | 44.188% |
| Item | Count | Percentage |
|---|---|---|
| All | 37,644 | 100.00% |
| KEGG | 8050 | 21.38% |
| Pathway | 4156 | 11.04% |
| Nr | 27,713 | 73.62% |
| Uniprot | 27,433 | 72.87% |
| GO | 20,207 | 53.68% |
| KOG (EuKaryotic Orthologous Groups) | 98 | 0.26% |
| Pfam | 15,849 | 42.10% |
| TF (Transcription factor) | 758 | 2.01% |
| Comparison | Significant Diff Number | Up | Down |
|---|---|---|---|
| T0 h vs. T6 h | 713 | 353 | 360 |
| T0 h vs. T12 h | 1292 | 366 | 926 |
| T0 h vs. T18 h | 634 | 319 | 315 |
| T0 h vs. T24 h | 496 | 157 | 339 |
| T0 h vs. F24 h | 1121 | 485 | 636 |
| Time | ZT 0-ZT 3 | ZT 3-ZT 6 | ZT 6-ZT 9 | ZT 9-ZT 12 | ZT 12-ZT 15 | ZT 15-ZT 18 | ZT 18-ZT 21 | ZT 21-ZT 24 | |
|---|---|---|---|---|---|---|---|---|---|
| Groups | |||||||||
| 6L6D | 6 h (09:00~15:00) | 6 h (15:00~21:00) | 6 h (21:00~ + 103:00) | 6 h (+ 103:00~ + 109:00) | |||||
| 24 L | 24 h (09:00~ + 109:00) | ||||||||
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| CsCHLE1.1 | TGAAGGCGAATCCAGAGT | ATACCGAGAGGCAGAAGAA |
| CsCHLE1.2 | TGCCTCTCGGTATATGTGA | TCTCTTGAACTCTGGATTCTC |
| CsCHLP1.1 | CCTTCATCATCTCCGCTAA | GCTCCAATCACAACATCAA |
| CsCHLP1.2 | CATAGACCGCCGAGTAAC | TGAGGAAGAGACCGTTGA |
| CsCLH1.1 | CGAAGCGTAGATGACCAA | GTGTTACAGGAGCAATAGTAG |
| CsCLH1.2 | TTCAATCCACACTGCCTAA | TCTTCATTCTCACCATCCAA |
| CsPOR1.1 | GTCGGCTCAATTACAGGTA | CCACATCACTCGCTTCTT |
| CsPOR1.2 | GTTGATGATGGAGGACTTGA | CGGATTCTTCTTCGGATACA |
| GAPDH | TTGGCATCGTTGAGGGTCT | CAGTGGGAACACGGAAAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.-H.; Sun, M.-Z.; Yang, K.-X.; Zhang, N.; Chen, C.; Xiong, J.-W.; Yang, N.; Chen, Y.; Liu, H.; Li, X.-H.; et al. High-Throughput Transcriptomic Analysis of Circadian Rhythm of Chlorophyll Metabolism under Different Photoperiods in Tea Plants. Int. J. Mol. Sci. 2024, 25, 9270. https://doi.org/10.3390/ijms25179270
Hu Z-H, Sun M-Z, Yang K-X, Zhang N, Chen C, Xiong J-W, Yang N, Chen Y, Liu H, Li X-H, et al. High-Throughput Transcriptomic Analysis of Circadian Rhythm of Chlorophyll Metabolism under Different Photoperiods in Tea Plants. International Journal of Molecular Sciences. 2024; 25(17):9270. https://doi.org/10.3390/ijms25179270
Chicago/Turabian StyleHu, Zhi-Hang, Meng-Zhen Sun, Kai-Xin Yang, Nan Zhang, Chen Chen, Jia-Wen Xiong, Ni Yang, Yi Chen, Hui Liu, Xing-Hui Li, and et al. 2024. "High-Throughput Transcriptomic Analysis of Circadian Rhythm of Chlorophyll Metabolism under Different Photoperiods in Tea Plants" International Journal of Molecular Sciences 25, no. 17: 9270. https://doi.org/10.3390/ijms25179270
APA StyleHu, Z.-H., Sun, M.-Z., Yang, K.-X., Zhang, N., Chen, C., Xiong, J.-W., Yang, N., Chen, Y., Liu, H., Li, X.-H., Chen, X., Xiong, A.-S., & Zhuang, J. (2024). High-Throughput Transcriptomic Analysis of Circadian Rhythm of Chlorophyll Metabolism under Different Photoperiods in Tea Plants. International Journal of Molecular Sciences, 25(17), 9270. https://doi.org/10.3390/ijms25179270