
Developmental Toxicity and Apoptosis in Zebrafish: The Impact of Lithium Hexafluorophosphate (LiPF6) from Lithium-Ion Battery Electrolytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
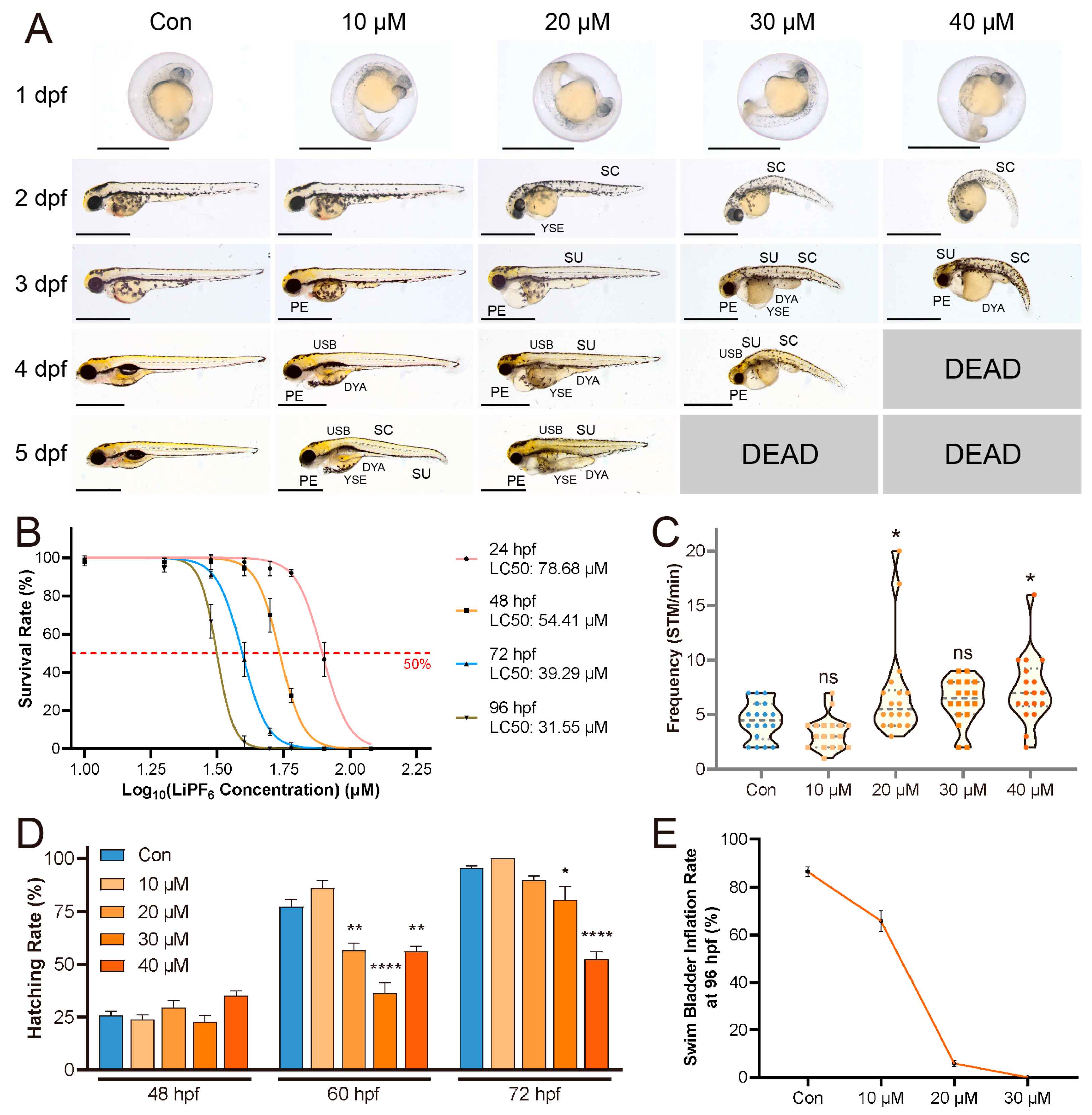
2.1. Developmental Abnormalities Induced by LiPF6 in Zebrafish
2.2. LiPF6-Induced Downregulation of anxa5b in Zebrafish Swim Bladder Development
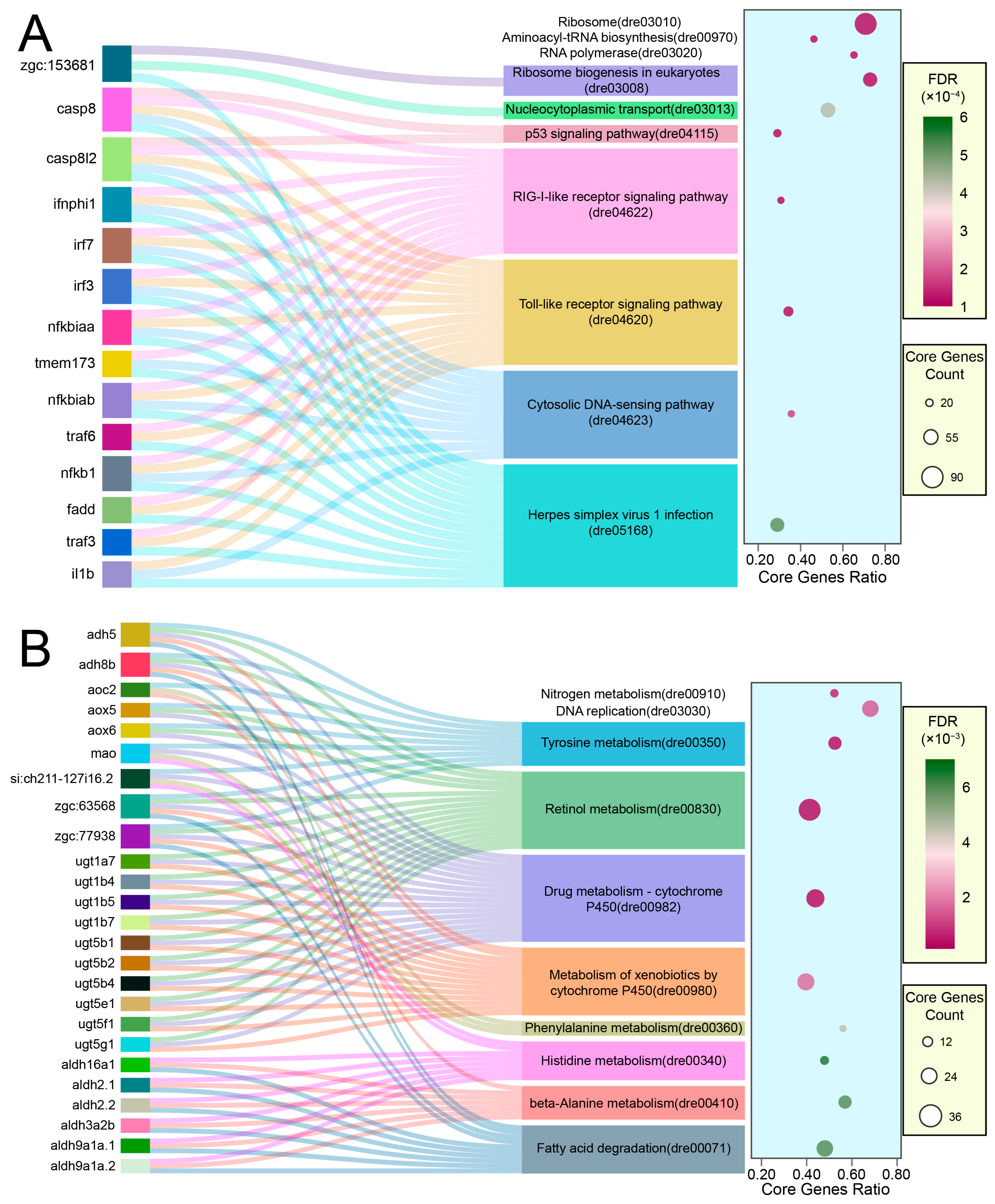
2.3. Molecular Signatures of LiPF6 Toxicity Unveiled through Transcriptomic Profiling
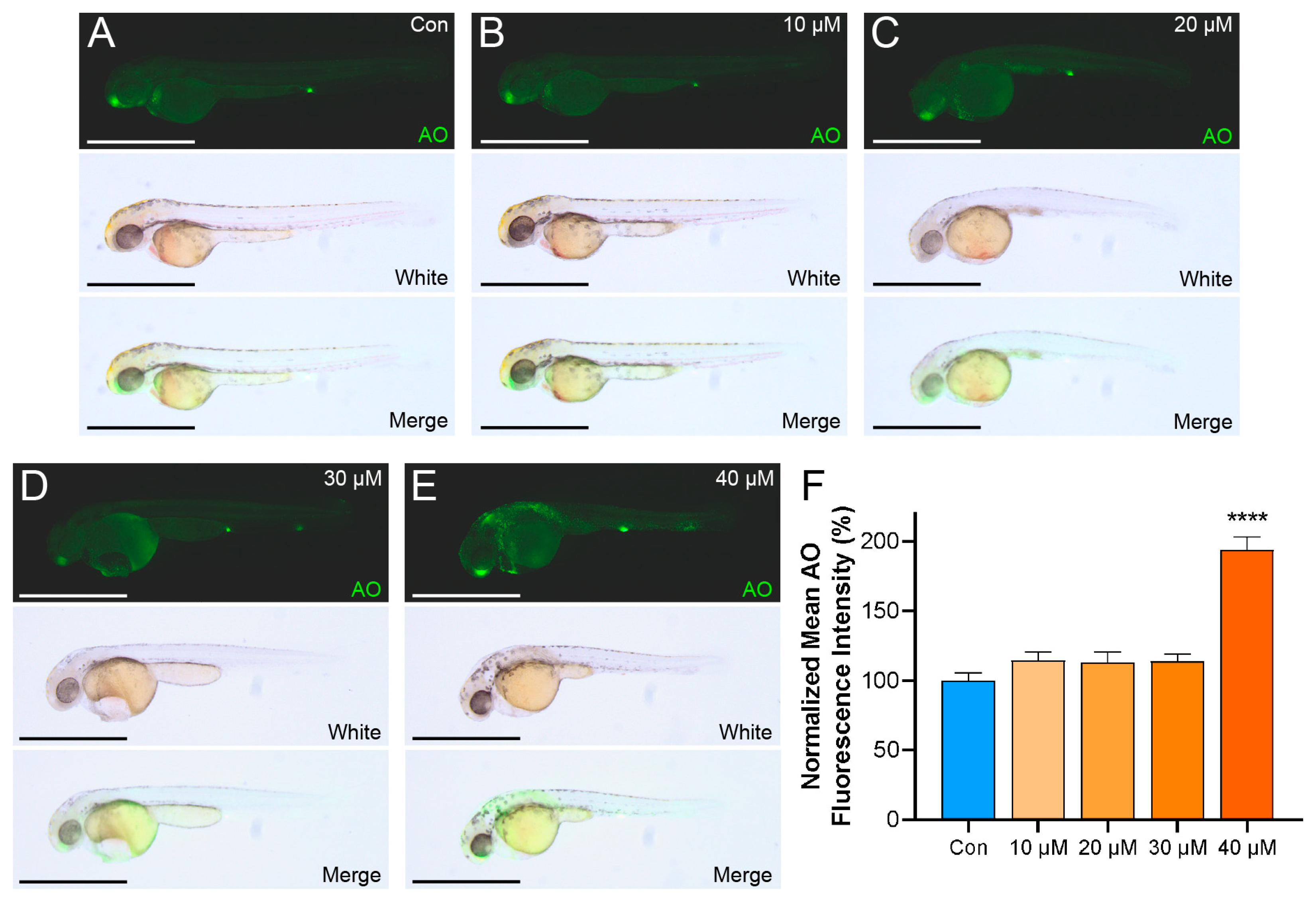
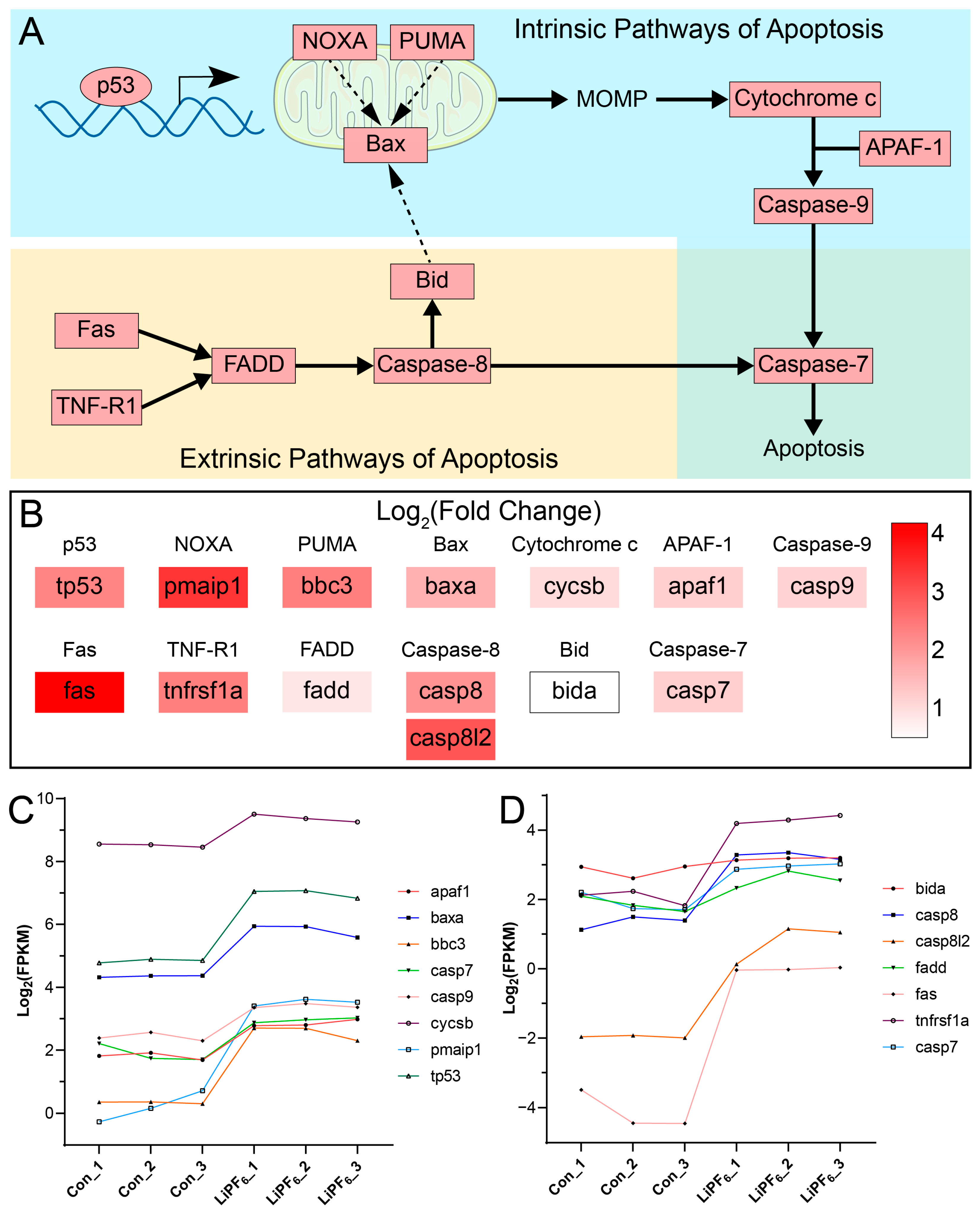
2.4. Zebrafish Exhibit a Pronounced Apoptotic Response upon LiPF6 Exposure
3. Discussion
4. Materials and Methods
4.1. Cultivation and Propagation of Zebrafish
4.2. Embryo Exposure Protocol
4.3. Assessment of LiPF6 Toxicity in Zebrafish Development
4.4. Whole-Mount In Situ Hybridization
4.5. RNA-Seq and Analysis
4.6. Assessment of Apoptosis in Zebrafish Embryos via AO Staining
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Oliveira Demarco, J.; Stefanello Cadore, J.; da Silveira de Oliveira, F.; Hiromitsu Tanabe, E.; Assumpção Bertuol, D. Recovery of metals from spent lithium-ion batteries using organic acids. Hydrometallurgy 2019, 190, 105169. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Y.; Zhou, T.; Liu, D.; Hu, H.; Fan, S. Hydrometallurgical recovery of metal values from sulfuric acid leaching liquor of spent lithium-ion batteries. Waste Manag. 2015, 38, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Voronina, N.; Sun, Y.-K.; Myung, S.-T. Co-Free Layered Cathode Materials for High Energy Density Lithium-Ion Batteries. ACS Energy Lett. 2020, 5, 1814–1824. [Google Scholar] [CrossRef]
- Ministry of Industry and Information Technology of the People’s Republic of China. The National Operation of Lithium-Ion Battery Industry in 2023. Available online: https://www.miit.gov.cn/gxsj/tjfx/dzxx/art/2024/art_7de3ac5f07514eec80039a81aa2e22d8.html (accessed on 12 July 2024).
- Liu, Y.-K.; Zhao, C.-Z.; Du, J.; Zhang, X.-Q.; Chen, A.-B.; Zhang, Q. Research Progresses of Liquid Electrolytes in Lithium-Ion Batteries. Small 2023, 19, 2205315. [Google Scholar] [CrossRef]
- Mao, Z.; Song, Y.; Zhen, A.G.; Sun, W. Recycling of electrolyte from spent lithium-ion batteries. Next Sustain. 2024, 3, 100015. [Google Scholar] [CrossRef]
- Nikiforidis, G.; Raghibi, M.; Sayegh, A.; Anouti, M. Low-Concentrated Lithium Hexafluorophosphate Ternary-based Electrolyte for a Reliable and Safe NMC/Graphite Lithium-Ion Battery. J. Phys. Chem. Lett. 2021, 12, 1911–1917. [Google Scholar] [CrossRef]
- Schmuch, R.; Wagner, R.; Hörpel, G.; Placke, T.; Winter, M. Performance and cost of materials for lithium-based rechargeable automotive batteries. Nat. Energy 2018, 3, 267–278. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; DeCarolis, J.F.; Barlaz, M.A. Evaluation of optimal model parameters for prediction of methane gener ation from selected U.S. landfills. Waste Manag. 2019, 91, 120–127. [Google Scholar] [CrossRef]
- Costa, A.M.; Alfaia, R.G.d.S.M.; Campos, J.C. Landfill leachate treatment in Brazil—An overview. J. Environ. Manag. 2019, 232, 110–116. [Google Scholar] [CrossRef]
- Havukainen, J.; Zhan, M.; Dong, J.; Liikanen, M.; Deviatkin, I.; Li, X.; Horttanainen, M. Environmental impact assessment of municipal solid waste management incorporating mechanical treatment of waste and incineration in Hangzhou, China. J. Clean. Prod. 2017, 141, 453–461. [Google Scholar] [CrossRef]
- Mrozik, W.; Rajaeifar, M.A.; Heidrich, O.; Christensen, P. Environmental impacts, pollution sources and pathways of spent lithium-ion batteries. Energy Environ. Sci. 2021, 14, 6099–6121. [Google Scholar] [CrossRef]
- Li, P.; Luo, S.; Zhang, L.; Liu, Q.; Wang, Y.; Lin, Y.; Xu, C.; Guo, J.; Cheali, P.; Xia, X. Progress, challenges, and prospects of spent lithium-ion batteries recycling: A review. J. Energy Chem. 2024, 89, 144–171. [Google Scholar] [CrossRef]
- Li, P.; Luo, S.-h.; Su, F.; Zhang, L.; Yan, S.; Lei, X.; Mu, W.; Wang, Q.; Zhang, Y.; Liu, X.; et al. Optimization of Synergistic Leaching of Valuable Metals from Spent Lithium-Ion Batteries by the Sulfuric Acid-Malonic Acid System Using Response Surface Methodology. ACS Appl. Mater. Interfaces 2022, 14, 11359–11374. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Stegmayr, B.; Salander Renberg, E.; Werneke, U. Lithium intoxication: Incidence, clinical course and renal function—A population-based retrospective cohort study. J. Psychopharmacol. 2016, 30, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Desmond, M.; Berk, M. Lithium and nephrotoxicity: A literature review of approaches to clinical management and risk stratification. BMC Nephrol. 2018, 19, 305. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, E.; Rajmohan, V. Lithium use in special populations. Indian J. Psychiatry 2007, 49, 211–218. [Google Scholar] [CrossRef]
- Canan, F.; Kaya, A.; Bulur, S.; Albayrak, E.S.; Ordu, S.; Ataoglu, A. Lithium intoxication related multiple temporary ecg changes: A case report. Cases J. 2008, 1, 156. [Google Scholar] [CrossRef]
- Kibirige, D.; Luzinda, K.; Ssekitoleko, R. Spectrum of lithium induced thyroid abnormalities: A current perspective. Thyroid. Res. 2013, 6, 3. [Google Scholar] [CrossRef]
- McKnight, R.F.; Adida, M.; Budge, K.; Stockton, S.; Goodwin, G.M.; Geddes, J.R. Lithium toxicity profile: A systematic review and meta-analysis. Lancet 2012, 379, 721–728. [Google Scholar] [CrossRef]
- Apostoli, P.; Catalani, S. Metal ions affecting reproduction and development. Met. Ions Life Sci. 2011, 8, 263–303. [Google Scholar]
- Jin, X.; Zhang, J.; An, T.; Zhao, H.; Fu, W.; Li, D.; Liu, S.; Cao, X.; Liu, B. A Genome-Wide Screen in Saccharomyces cerevisiae Reveals a Critical Role for Oxidative Phosphorylation in Cellular Tolerance to Lithium Hexafluorophosphate. Cells 2021, 10, 888. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Tal, T.; Yaghoobi, B.; Lein, P.J. Translational Toxicology in Zebrafish. Curr. Opin. Toxicol. 2020, 23–24, 56–66. [Google Scholar] [CrossRef]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An emerging model to study microplastic and nanoplastic toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef]
- Katoch, S.; Patial, V. Zebrafish: An emerging model system to study liver diseases and related drug discovery. J. Appl. Toxicol. 2021, 41, 33–51. [Google Scholar] [CrossRef]
- Winata, C.L.; Korzh, S.; Kondrychyn, I.; Zheng, W.; Korzh, V.; Gong, Z. Development of zebrafish swimbladder: The requirement of Hedgehog signaling in specification and organization of the three tissue layers. Dev. Biol. 2009, 331, 222–236. [Google Scholar] [CrossRef]
- Wu, Y.; Li, W.; Yuan, M.; Liu, X. The synthetic pyrethroid deltamethrin impairs zebrafish (Danio rerio) swim bladder development. Sci. Total Environ. 2020, 701, 134870. [Google Scholar] [CrossRef] [PubMed]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional control of human p53-regulated genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Deng, Y. Bax and BH3-domain-only proteins in p53-mediated apoptosis. FBL 2002, 7, 151–156. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef]
- Nakano, K.; Vousden, K.H. PUMA, a novel proapoptotic gene, is induced by p53. Mol. Cell 2001, 7, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C.; Borutaite, V. Regulation of apoptosis by the redox state of cytochrome c. Biochim. Biophys. Acta (BBA)-Bioenerg. 2008, 1777, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Kuida, K. Caspase-9. Int. J. Biochem. Cell Biol. 2000, 32, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, R.; Kheirollahi, A.; Davoodi, J. Apaf-1: Regulation and function in cell death. Biochimie 2017, 135, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D. Caspase-7: A protease involved in apoptosis and inflammation. Int. J. Biochem. Cell Biol. 2010, 42, 21–24. [Google Scholar] [CrossRef]
- Gaur, U.; Aggarwal, B.B. Regulation of proliferation, survival and apoptosis by members of the TNF superfamily. Biochem. Pharmacol. 2003, 66, 1403–1408. [Google Scholar] [CrossRef]
- Lee, E.W.; Seo, J.; Jeong, M.; Lee, S.; Song, J. The roles of FADD in extrinsic apoptosis and necroptosis. BMB Rep. 2012, 45, 496–508. [Google Scholar] [CrossRef]
- Mandal, R.; Barrón, J.C.; Kostova, I.; Becker, S.; Strebhardt, K. Caspase-8: The double-edged sword. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2020, 1873, 188357. [Google Scholar] [CrossRef]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef]
- Hung, C.-L.; Chang, H.-H.; Lee, S.W.; Chiang, Y.-W. Stepwise activation of the pro-apoptotic protein Bid at mitochondrial membranes. Cell Death Differ. 2021, 28, 1910–1925. [Google Scholar] [CrossRef]
- Schug, Z.T.; Gonzalvez, F.; Houtkooper, R.H.; Vaz, F.M.; Gottlieb, E. BID is cleaved by caspase-8 within a native complex on the mitochondrial membrane. Cell Death Differ. 2011, 18, 538–548. [Google Scholar] [CrossRef]
- Billen, L.P.; Shamas-Din, A.; Andrews, D.W. Bid: A Bax-like BH3 protein. Oncogene 2008, 27 (Suppl. S1), S93–S104. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Zheng, L.; Cheng, P.; Wang, X.; Wu, H.; Ma, Q.; Liu, J.; Feng, W.; Nie, J.; Yu, H.; et al. Taming the chemical instability of lithium hexafluorophosphate-based electrolyte with lithium fluorosulfonimide salts. J. Power Sources 2022, 526, 231105. [Google Scholar] [CrossRef]
- Spotte-Smith, E.W.C.; Petrocelli, T.B.; Patel, H.D.; Blau, S.M.; Persson, K.A. Elementary Decomposition Mechanisms of Lithium Hexafluorophosphate in Battery Electrolytes and Interphases. ACS Energy Lett. 2023, 8, 347–355. [Google Scholar] [CrossRef]
- Tasaki, K.; Kanda, K.; Nakamura, S.; Ue, M. Decomposition of LiPF6 and Stability of PF5 in Li-Ion Battery Electrolytes. Density Functional Theory and Molecular Dynamics Studies. ChemInform 2004, 35. [Google Scholar] [CrossRef]
- Sheng, L.; Zhu, D.; Yang, K.; Wu, Y.; Wang, L.; Wang, J.; Xu, H.; He, X. Unraveling the Hydrolysis Mechanism of LiPF6 in Electrolyte of Lithium Ion Batteries. Nano Lett. 2024, 24, 533–540. [Google Scholar] [CrossRef]
- Stich, M.; Göttlinger, M.; Kurniawan, M.; Schmidt, U.; Bund, A. Hydrolysis of LiPF6 in Carbonate-Based Electrolytes for Lithium-Ion Batteries and in Aqueous Media. J. Phys. Chem. C 2018, 122, 8836–8842. [Google Scholar] [CrossRef]
- Terborg, L.; Nowak, S.; Passerini, S.; Winter, M.; Karst, U.; Haddad, P.R.; Nesterenko, P.N. Ion chromatographic determination of hydrolysis products of hexafluorophosphate salts in aqueous solution. Anal. Chim. Acta 2012, 714, 121–126. [Google Scholar] [CrossRef]
- Valcarce, D.G.; Sellés-Egea, A.; Riesco, M.F.; De Garnica, M.-G.; Martínez-Fernández, B.; Herráez, M.P.; Robles, V. Early stress exposure on zebrafish development: Effects on survival, malformations and molecular alterations. Fish Physiol. Biochem. 2024, 50, 1545–1562. [Google Scholar] [CrossRef]
- Stinckens, E.; Vergauwen, L.; Schroeder, A.L.; Maho, W.; Blackwell, B.R.; Witters, H.; Blust, R.; Ankley, G.T.; Covaci, A.; Villeneuve, D.L.; et al. Impaired anterior swim bladder inflation following exposure to the thyroid peroxidase inhibitor 2-mercaptobenzothiazole part II: Zebrafish. Aquat. Toxicol. 2016, 173, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Van Dingenen, I.; Vergauwen, L.; Haigis, A.C.; Blackwell, B.R.; Stacy, E.; Villeneuve, D.L.; Knapen, D. Deiodinase inhibition impairs the formation of the three posterior swim bladder tissue layers during early embryonic development in zebrafish. Aquat. Toxicol. 2023, 261, 106632. [Google Scholar] [CrossRef]
- Sun, H.; Chen, M.; Wang, Z.; Zhao, G.; Liu, J.X. Transcriptional profiles and copper stress responses in zebrafish cox17 mutants. Environ. Pollut. 2020, 256, 113364. [Google Scholar] [CrossRef]
- Teoh, P.H.; Shu-Chien, A.C.; Chan, W.K. Pbx1 is essential for growth of zebrafish swim bladder. Dev. Dyn. 2010, 239, 865–874. [Google Scholar] [CrossRef]
- Abrams, J.M.; White, K.; Fessler, L.I.; Steller, H. Programmed cell death during Drosophila embryogenesis. Development 1993, 117, 29–43. [Google Scholar] [CrossRef]
- Arama, E.; Steller, H. Detection of apoptosis by terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling and acridine orange in Drosophila embryos and adult male gonads. Nat. Protoc. 2006, 1, 1725–1731. [Google Scholar] [CrossRef]
- Tucker, B.; Lardelli, M. A Rapid Apoptosis Assay Measuring Relative Acridine Orange Fluorescence in Zebrafish Embryos. Zebrafish 2007, 4, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Thisse, B.; Thisse, C. In Situ Hybridization on Whole-Mount Zebrafish Embryos and Young Larvae. In In Situ Hybridization Protocols; Nielsen, B.S., Ed.; Springer: New York, NY, USA, 2014; pp. 53–67. [Google Scholar]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef]
- Li, M.; Yu, T.; Lai, J.; Han, X.; Hu, J.; Deng, Z.; Li, D.; Ye, Z.; Wang, S.; Hu, C.; et al. Ethoprophos induces cardiac toxicity in zebrafish embryos. Ecotoxicol. Environ. Saf. 2021, 228, 113029. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Sun, L.; Peng, Z.; Zhang, Q.; Lin, M.; Peng, Z.; Zheng, L. Developmental Toxicity and Apoptosis in Zebrafish: The Impact of Lithium Hexafluorophosphate (LiPF6) from Lithium-Ion Battery Electrolytes. Int. J. Mol. Sci. 2024, 25, 9307. https://doi.org/10.3390/ijms25179307
Yang B, Sun L, Peng Z, Zhang Q, Lin M, Peng Z, Zheng L. Developmental Toxicity and Apoptosis in Zebrafish: The Impact of Lithium Hexafluorophosphate (LiPF6) from Lithium-Ion Battery Electrolytes. International Journal of Molecular Sciences. 2024; 25(17):9307. https://doi.org/10.3390/ijms25179307
Chicago/Turabian StyleYang, Boyu, Luning Sun, Zheng Peng, Qing Zhang, Mei Lin, Zhilin Peng, and Lan Zheng. 2024. "Developmental Toxicity and Apoptosis in Zebrafish: The Impact of Lithium Hexafluorophosphate (LiPF6) from Lithium-Ion Battery Electrolytes" International Journal of Molecular Sciences 25, no. 17: 9307. https://doi.org/10.3390/ijms25179307